
Presenting Eco-Anatomical Data for Saponaria jagelii, a Species on the Edge of the Blade

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.1.1. Seed Germination
2.1.2. Microscopy
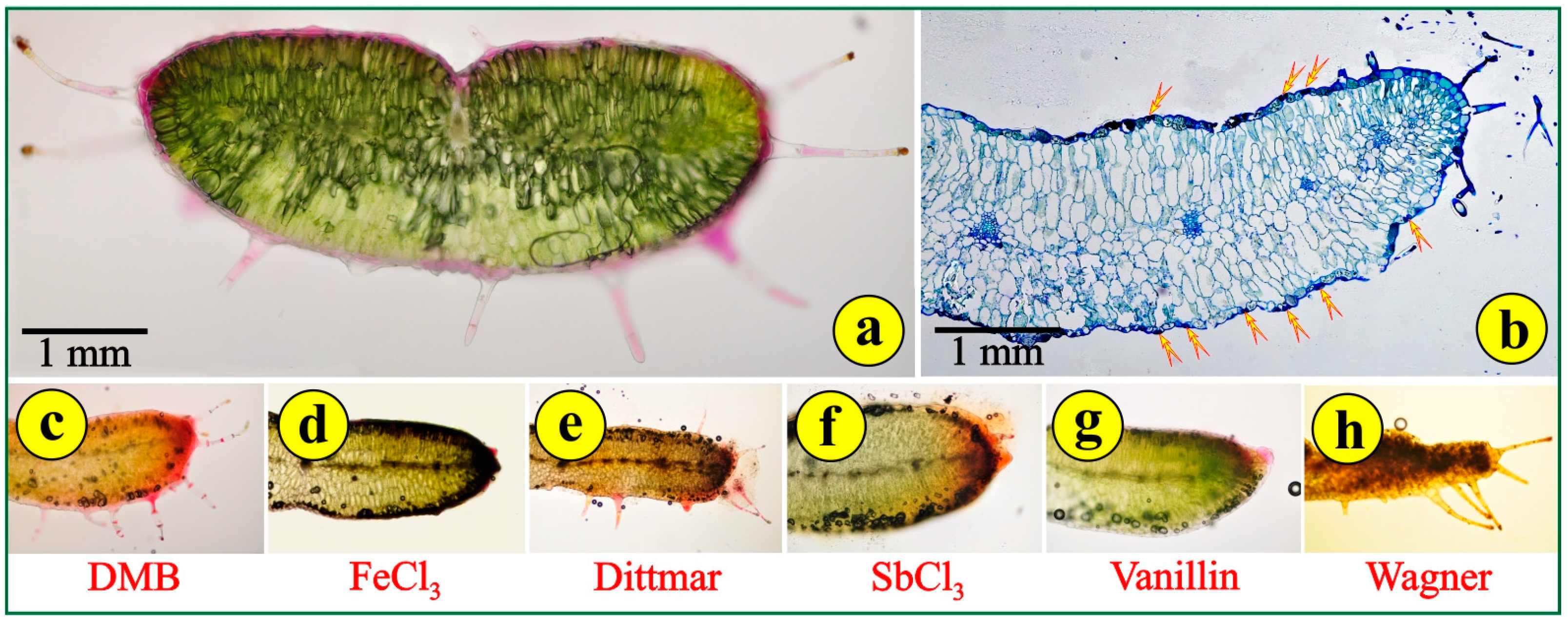
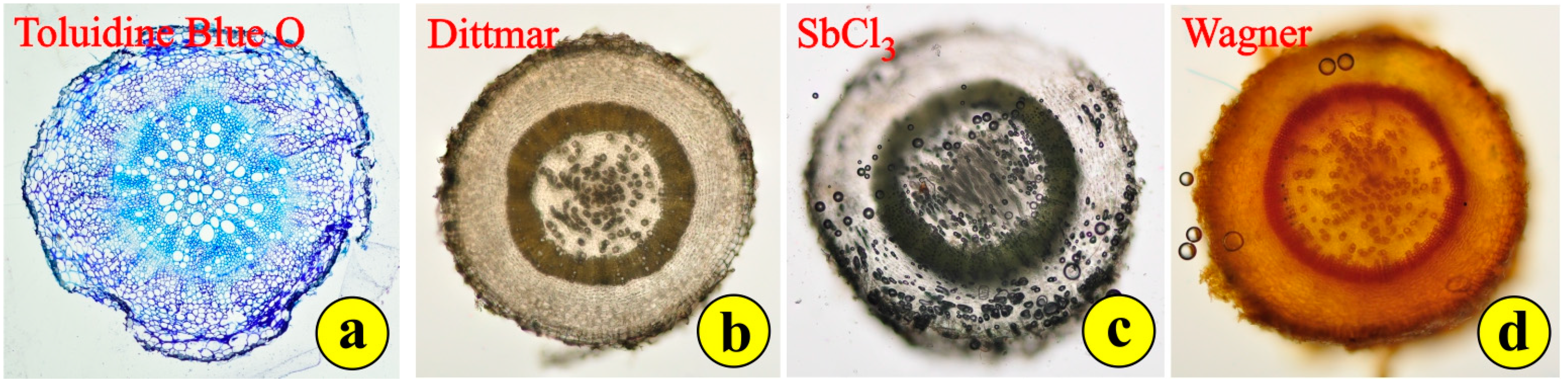
2.1.3. Histochemistry
- (a)
- Osmium tetroxide [13] for unsaturated lipids;
- (b)
- Concentrated H2SO4 [14] for sesquiterpenes;
- (c)
- Vanillin/HCl [15] for flavonoids;
- (d)
- Antimony trichloride (SbCl3) [16] for terpene-containing steroids;
- (e)
- Dittmar’s reagent [17] for alkaloids;
- (f)
- Potassium bichromate [18] for phenolics;
- (g)
- Alcoholic vanillin-HCl (vanillin test) [19] for phenolic compounds;
- (h)
- Ferric chloride [20] for polyphenols;
- (i)
- 4-Nitrosophenol in concentrated H2SO4 [21] for monoterpene phenols;
- (j)
- Wagner and Ellram reagents [17] for alkaloids;
- (k)
- DMB (3,4-dimethoxy-benzaldehyde or veratraldehyde) for phenolic tannin precursors [22].
3. Results and Discussion
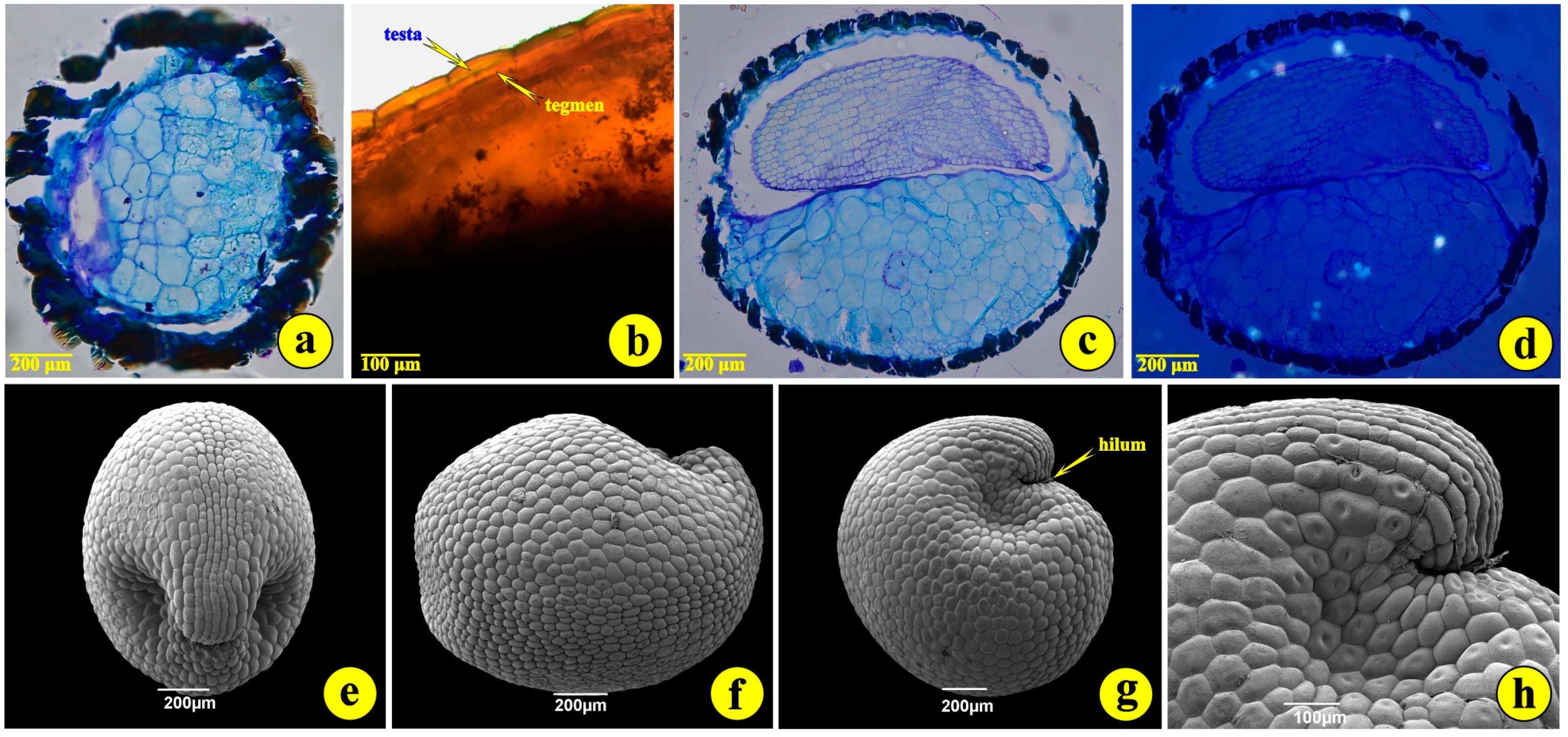
3.1. Seed Morphology and Anatomy—Germination
3.2. Leaf and Root Anatomy
3.3. Flower and Fruit Anatomy
3.4. Habitat, Threats and Protection
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phitos, D.; Greuter, W. Saponaria jagelii: A new species from the island of Elafonisos (Peloponnisos, Greece). Flora Mediterr. 1993, 3, 277–278. [Google Scholar]
- Bergmeier, E.; Ristow, M.; Krause, J.; Meyer, S.; Panitsa, M. Phytodiversity of Limnos (North Aegean, Greece)–an update and evaluation. Flora Mediterr. 2021, 31, 233–246. [Google Scholar] [CrossRef]
- Phitos, D.; Kamari, G. Saponaria jagelii. In The Red Data Book of Rare and Threatened Plants of Greece; Phitos, D., Strid, A., Snogerup, S., Greuter, W., Eds.; WWF-Greece: Athens, Greece, 1995. [Google Scholar]
- Iatrou, G. Saponaria jagelii. In The IUCN Red List of Threatened Species; Errata Version Published in 2016; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2013; p. E.T61639A50017799. Available online: https://www.iucnredlist.org/species/61639/103010620 (accessed on 1 February 2024).
- Pasta, S.; Perez-Graber, A.; Fazan, L.; de Montmollin, B. (Eds.) The Top 50 Mediterranean Island Plants UPDATE 2017; E-Book and on Line; IUCN/SSC/Mediterranean Plant Specialist Group: Neuchâtel, Switzerland, 2017; 141p, Available online: https://top50.iucn-mpsg.org/species/48 (accessed on 1 February 2024).
- Salafsky, N.; Salzer, D.; Stattersfield, A.J.; Hilton-Taylor, C.; Neugarten, R.; Butchart, S.H.; Collen, B.; Cox, N.; Master, L.L.; O’Connor, S.; et al. A standard lexicon for biodiversity conservation: Unified classifications of threats and actions. Conserv. Biol. 2008, 22, 897–911. [Google Scholar] [CrossRef] [PubMed]
- O’ Brien, T.P.; McCully, M.E. The Study of Plant Structure: Principles and Selected Methods; Termocarphy: Melbourne, Australia, 1981; 357p, ISBN 0-9594174-0-0. [Google Scholar]
- Christodoulakis, N.S.; Lampri, P.-N.; Fasseas, C. Structural and cytochemical investigation on the leaf of silverleaf nightshade (Solanum elαeagnifolium), a drought resistant alien weed of the Greek Flora. Aust. J. Bot. 2009, 57, 432–438. [Google Scholar] [CrossRef]
- Christodoulakis, N.S.; Kogia, D.; Mavroeidi, D.; Fasseas, C. Anatomical and cytochemical investigation on the leaf of Teucrium polium L., a pharmaceutical shrub of the Greek phryganic formations. J. Biol. Res. 2010, 14, 199–209. [Google Scholar]
- Bronner, R. Simultaneous demonstration of lipid and starch in plant tissues. Stain. Technol. 1975, 50, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Mowry, R.W. Alcian blue techniques for the histochemical study of acidic carbohydrates. J. Histochem. Cytochem. 1956, 4, 407–411. [Google Scholar]
- Fisher, D.B. Protein staining of ribboned epon sections for light microscopy. Histochemie 1968, 16, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Lison, L. Histochimie et Cytochimie Animales: Principes et Méthodes par L. Lison, 2nd ed.; Gauthier-Villars: Paris, France, 1960. [Google Scholar]
- Cappelletti, E.M.; Caniato, R.; Appendino, G. Localization of cytotoxic hydroperoxy-eudesmanolides in Artemisia umbelliformis. Biochem. Syst. Ecol. 1986, 14, 183–190. [Google Scholar] [CrossRef]
- Guerin, H.P.; Delaveau, P.G.; Paris, R.R. Localisations histochimiques. II: Procédes simples de localisation de pigments flavoniques. Application a quelques Phanerogrames. Bull. Soc. Bot. Fr. 1971, 118, 29–36. [Google Scholar] [CrossRef]
- Hardman, R.; Sofowora, E.A. Antimony trichloride as a test reagent for steroids, especially diosgenin and yamogenin, in plant tissues. Stain. Technol. 1972, 47, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Furr, M.; Mahlberg, P.G. Histochemical analyses of laticifers and glandular trichomes in Cannabis sativa. J. Nat. Prod. 1981, 44, 153–159. [Google Scholar] [CrossRef]
- Gabe, M. Techniques Histologiques; Masson & Cie: Paris, France, 1968; p. 1113. [Google Scholar]
- Gardner, R.O. Vanillin-hydrochloric acid as a histochemical test for tannin. Stain. Technol. 1975, 50, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill: New York, NY, USA; London, UK, 1940. [Google Scholar]
- Gersbach, P.V.; Wyllie, S.G.; Sarafis, V. A new histochemical method for localization of the site of monoterpene phenol accumulation in plant secretory structures. Ann. Bot. 2001, 88, 521–525. [Google Scholar] [CrossRef]
- Mace, M.E.; Howell, C.R. Histochemistry and identification of condensed tannin precursors in root of cotton seedlings. Can. J. Bot. 1972, 52, 2423–2426. [Google Scholar] [CrossRef]
- Toderich, K.; Shuyskaya, E.; Khujanazarov, T.; Ismail, S.; Kawabata, Y. The Structural and Functional Characteristics of Asiatic Desert Halophytes for Phytostabilization of Polluted Sites. In Plant Adaptation and Phytoremediation; Ashraf, M., Ozturk, M., Ahmad, M., Eds.; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar] [CrossRef]
- Mattana, E.; Carta, A.; Fernández Pascual, E.; Keeley, J.E.; Pritchard, H.W. Climate change and plant regeneration from seeds in Mediterranean regions of the northern hemisphere. In Plant Regeneration from Seeds: A Global Warming Perspective; Baskin, C.C., Baskin, J.M., Eds.; Academic Press: London, UK, 2022; pp. 101–114. [Google Scholar]
- Stefi, A.L.; Katsikis, N.; Thanos, C.A. An investigation on the population, ecophysiology and morphology of Saponaria jagelii, a critically endangered species. In Proceedings of the HELECOS-11 Conference, Patras, Greece, 4–7 October 2023. [Google Scholar]
- Stefi, A.L.; Thanos, C.A. There’s still hope for Saponaria jagelii. In Reserach Report for the Second Year of the Research Project “Conserving the Flora of the Balkans: Native Plants of Greece”; NKUA Seed Bank: Athens, Greece, 2023; Available online: http://users.uoa.gr/~cthanos/NKUASB_SeedConservationoftheFloraofGreece/pdfs/Saponaria_hope.pdf (accessed on 1 February 2024).
- Karabourniotis, G.; Fasseas, C. The dense indumentum with its polyphenol content may replace the protective role of the epidermis in some young xeromorphic leaves. Can. J. Bot. 1996, 74, 347–351. [Google Scholar] [CrossRef]
- Dimopoulos, P.; Raus, T.; Bergmeir, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular Plants of Greece: An Annotated Checklist. Supplement Willdenowia; Botanic Garden and Botanical Museum Berlin-Dahlem: Berlin, Germany; Hellenic Botanical Society: Athens, Greece, 2013; p. 81. [Google Scholar]
- Available online: https://weatherandclimate.com/greece/peloponnese/elafonissos (accessed on 30 January 2024).
- Zepner, L.; Karrasch, P.; Wiemann, F.; Bernard, L. ClimateCharts.net–an interactive climate analysis web platform. Int. J. Digit. Earth 2020, 14, 338–356. [Google Scholar] [CrossRef]
- Thanos, C.A.; Katsikis, N.; Oikonomidis, S.; Mouratidis, S.; Stefi, A.L.; Willey, I.; Faruk, A. The swan song for Greek endemic Saponaria jagelii? Samara Newsl. 2022, 38, 12. [Google Scholar]
- Kougioumoutzis, K. Species on the brink of extinction. In Research Report of the First Year of the Project SoBEx; Agricultural University of Athens: Athens, Greece, 2019. (In Greek) [Google Scholar]
- Hellenic Statistical Authority, ELSTAT. Available online: https://prnt.sc/57NfHHE6EOsI (accessed on 30 January 2024).
- Laguna, E. The Micro-Reserves as a Tool for Conservation of Threatened Plants in Europe; Nature and Environment 121; Council of Europe Publishing: Strasbourg, France, 2001. [Google Scholar]
- Laguna, E.; Fos, S.; Jiménez, J.; Volis, S. Role of Micro-Reserves in Conservation of Endemic, Rare and Endangered Plants of the Valencian Region (Eastern Spain). Isr. J. Plant Sci. 2016, 63, 320–332. [Google Scholar] [CrossRef]
- Fos, S.; Laguna, E.; Jiménez, J.; Gómez-Serrano, M.Á. Plant Micro-Reserves in Valencia (E. Spain): A Model to Preserve Threatened Flora in China? Plant Divers. 2017, 39, 383–389. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2018 | 2019 | 2020 * | 2021 * | 2022 | Months |
|---|---|---|---|---|---|
| 5.135 | 5.622 | 5.436 | 6.332 | 5.533 | J, F, M |
| 28.957 | 36.232 | 15.603 | 32.971 | 32.983 | A, M, J |
| 106.661 | 122.234 | 116.625 | 124.017 | 124.187 | J, A, S |
| 9.072 | 9.645 | 9.772 | 9.147 | 13.335 | O, N, D |
| 120.868 | 173.733 | 147.436 | 172.467 | 176.038 | total |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefi, A.L.; Mitsigiorgi, K.; Christodoulakis, N.S. Presenting Eco-Anatomical Data for Saponaria jagelii, a Species on the Edge of the Blade. Life 2024, 14, 398. https://doi.org/10.3390/life14030398
Stefi AL, Mitsigiorgi K, Christodoulakis NS. Presenting Eco-Anatomical Data for Saponaria jagelii, a Species on the Edge of the Blade. Life. 2024; 14(3):398. https://doi.org/10.3390/life14030398
Chicago/Turabian StyleStefi, Aikaterina L., Konstantina Mitsigiorgi, and Nikolaos S. Christodoulakis. 2024. "Presenting Eco-Anatomical Data for Saponaria jagelii, a Species on the Edge of the Blade" Life 14, no. 3: 398. https://doi.org/10.3390/life14030398
APA StyleStefi, A. L., Mitsigiorgi, K., & Christodoulakis, N. S. (2024). Presenting Eco-Anatomical Data for Saponaria jagelii, a Species on the Edge of the Blade. Life, 14(3), 398. https://doi.org/10.3390/life14030398