
Capillary-Electrophoresis-Based Species Barcoding of Big Cats: CR-mtDNA-Length Polymorphism

1
Institute for Environmental Sciences, Charles University, 128 00 Prague, Czech Republic
2
Forensic DNA Service, Budinova 2, 180 81 Prague, Czech Republic
3
Department of Forensic Medicine, Second Faculty of Medicine, Charles University, 128 00 Prague, Czech Republic
4
Bulovka University Hospital, 180 81 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
Life 2024, 14(4), 497; https://doi.org/10.3390/life14040497
Submission received: 8 January 2024
/
Revised: 6 April 2024
/
Accepted: 9 April 2024
/
Published: 11 April 2024
(This article belongs to the Section Animal Science)
Abstract
:This study aimed to provide an overview of the methodological approach used for the species determination of big cats. The molecular system described herein employs mitochondrial DNA control region (CR-mtDNA)-length polymorphism in combination with highly sensitive and precise capillary electrophoresis. We demonstrated that the described CR-mtDNA barcoding system can be utilized for species determination where the presence of biological material from big cats is expected or used as a confirmatory test alongside Sanger or massive parallel sequencing (MPS). We have also addressed the fact that species barcoding, when based on the analysis of mtDNA targets, can be biased by nuclear inserts of the mitochondrial genome (NUMTs). The CR-mtDNA barcoding system is suitable even for problematic and challenging samples, such as hair. CR-mtDNA-length polymorphisms can also distinguish hybrids from pure breeds.
1. Introduction
The illegal trade of endangered species of wild fauna and flora threatens many species. Numerous traditional Chinese medicine (TCM) treatments involve the use of wildlife products, including some that utilize ingredients derived from endangered flora and fauna [1], regardless of whether those organisms are protected by the Convention on the International Trade in Endangered Species (CITES) [2]. The investigation of wildlife crimes requires precise scientific species determination techniques. DNA barcoding [3] seems to be an optimal tool for the detection of source organisms in TCM products. DNA barcoding sensu stricto means that species identification is performed using one standardized DNA fragment. The definition of DNA barcoding sensu lato is not very restrictive and corresponds with any taxonomic-level identification using any DNA fragment [4]. DNA barcoding of the biological material used for the preparation of TCMs has been described for numerous animal [5,6,7,8] and plant [9,10,11,12] species. Non-human DNA typing for forensic purposes is also utilized for the identification of domesticated animals, such as cats [13] and dogs [14], or CITES organisms, such as elephants [15], pangolins [16], rhinos [17], and tigers [18].
Standard Sanger sequencing is not suitable for samples containing biological material from multiple species; thus, massively parallel sequencing must be used instead [19]. Mitochondrial DNA control region length polymorphisms are an alternative to sequencing and can serve as an additional or confirmatory test. PCR amplification using universal primers with subsequent restriction cleavage (PCR-RFLP) can be used to separate species using characteristic patterns [20], but the use of this method is limited by the existence of restriction sites and rather difficult interpretation. Species barcoding can also be achieved using the RAPD approach [21], AFLPs [22], species-specific PCR [23], the SNaPshot assay [24], MRMA analysis [25], or qPCR assays [26]. Another type of barcoding methodology utilizes interspecies insertions/deletions, where the target region is a hypervariable mtDNA D-loop [27], the highly variable regions 12S rRNA 16S rRNA [28], and the control region (CR-mtDNA) length polymorphisms [29].
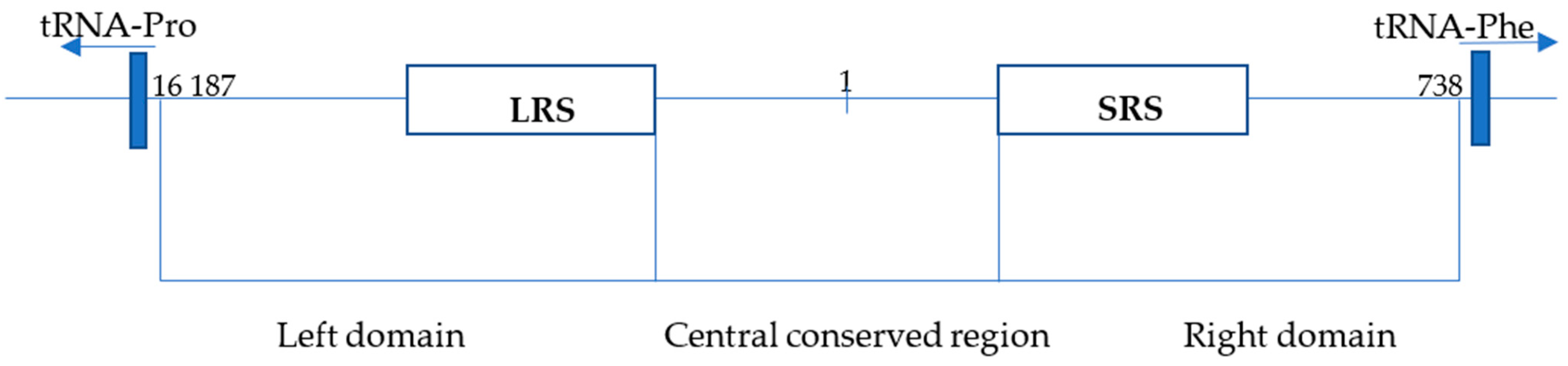
This study aims to provide an overview of the methodological approach used by our laboratory for species determination of big cats in our wildlife crime casework [30]. All species of big cats, including tigers, lions, leopards, cheetahs, and jaguars, are protected under the CITES convention but are poached in large numbers to serve as an article of illegal trade comprising bones, teeth, hide, and TCM products that are derived from these species [31,32]. The advantage of the CR-mtDNA barcoding system is its ease of use even in laboratories not performing sequencing techniques. The described assay can be utilized for species determination where the presence of biological material from big cats is expected or used as a confirmatory test alongside Sanger sequencing, MPS, or non-DNA barcoding methods. The molecular system described herein employs mitochondrial-DNA-control-region-length polymorphism (CR-mtDNA) [29] in combination with highly sensitive and precise capillary electrophoresis (see Figure 1).
2. Materials and Methods
The reference material used for this study was provided by the Czech Environmental Inspectorate and zoological gardens. The biological material came either from animals that died from natural causes in zoological gardens, or their excrements. The sampling did not involve the infliction of trauma to living animals. The research thus did not fall under the Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes.
Human DNA samples were sampled and used in accordance with the Regulation (EU) 2016/679 of the European Parliament and of the Council of 27 April 2016 on the protection of natural persons with regard to the processing of personal data and on the free movement of such data, following Directive 95/46/EC (General Data Protection Regulation).
DNA extraction from the reference material (see Table 1) was performed using a Quick-DNA Microprep Plus Kit (ZymoResearch, Irvine, CA, USA) with final elution to 20 µL H2O. The quantification of extracted DNA was achieved using UV–VIS or qPCR [18], where the nuclear DNA concentrations were measured by targeting the STR locus Pati01 [29].
2.1. PCR Composition and PCR Conditions for Fragment Analysis
The primers used for the amplification of mitochondrial-DNA-control-region-length polymorphisms have been described by Pun et al. [29]. Primer sequences: L15995 5′CTCCACTATCAGCACCCAAAG 3′; H16498 5′CCTGAAGTAAGAACCAGATG 3′.
The PCR mixture contained 1.25 μL of GoldStar 10× buffer (Promega, Madison, WI, USA), 10 μM L15995 + H16498 and 0.5 + 0.5 μL, and the primer L15995 was fluorescently labeled with 5-FAM, 0.25 μL (5 U/μL) of AmpliTag Gold DNA polymerase (Applied Biosystems, San Francisco, CA, USA), 10 pg of DNA, and H2O to a final volume of 12.5 μL. The PCR program was as follows: 95 °C for 10 min, 32× (95 °C for 15 s, 55 °C for 30 s, 72 °C for 1 min), and 72 °C for 30 min.
PCR was performed using a MasterCycler Nexus gradient thermocycler (Eppendorf, Germany). The resulting amplicons were visualized using capillary electrophoresis (SeqStudio 3200 Genetic Analyzer; Applied Biosystems, USA) under the following parameters: G5 matrix, 12 µL formamide, 0.4 µL LIZ 1200 (Applied Biosystems, USA), 1 µL of PCR product. Raw data processing was performed using GeneMapper5 (Applied Biosystems, USA).
2.2. PCR Composition and PCR Conditions for Agarose Gel Electrophoresis and Sanger Sequencing
The PCR mixture contained 10× Gold buffer (Applied Biosystems, USA), 2 μL of 25 mM MgCl2, 0.5 μL of 10 mM dNTPs, 0.5 + 0.5 μL of 10 μM L15995 + H16498, 0.2 μL of 5 U/μL AmpliTag Gold DNA polymerase (Applied Biosystems, USA), 0.1–1 ng of DNA, and H2O to a final volume of 25 μL. The PCR program was as follows: 95 °C for 10 min, 40× (95 °C for 15 s, 55 °C for 30 s, 72 °C for 1 min), and 72 °C for 30 min.
PCR was performed using a MasterCycler Nexus gradient thermocycler (Eppendorf, Germany). The resulting amplicons were visualized using agarose gel electrophoresis. DNA clean-up of the amplified fragments excised from the gel was performed using a Zymoclean Gel DNA recovery Kit (ZymoResearch, USA). Sanger sequencing was performed using a SeqStudio 3200 Genetic Analyzer (Applied Biosystems, USA). Raw data processing was performed using Sequencing Analysis Software v6.0 (Applied Biosystems, USA).
2.3. The Effect of Exonuclease V Treatment
Exonuclease V (RecBCD) (New England Biolabs, Ipswich, MA, USA) was used as suggested by the manufacturer.
To ensure nDNA removal, the molecular system Pleo Qplex was used. This system is based on quantitative RT-PCR with TaqMan probes and primers highly specific for Panthera leo (Forensic DNA Service, Prague, Czech Republic). The assay was performed in a single tube/well. Nuclear DNA concentration was measured using the STR locus Pati01 [29], and the concentration of mitochondrial DNA was measured using primers and probes targeted to the Cytochrome B gene of P. leo. Quantitative PCR was performed using QuantStudio 5 (Applied Biosystems, USA).
The assay validation followed the ANSI/ASB standard for the internal validation of forensic DNA analysis methods, as described by Webster et al. [37] (see Results section for validation).
3. Results
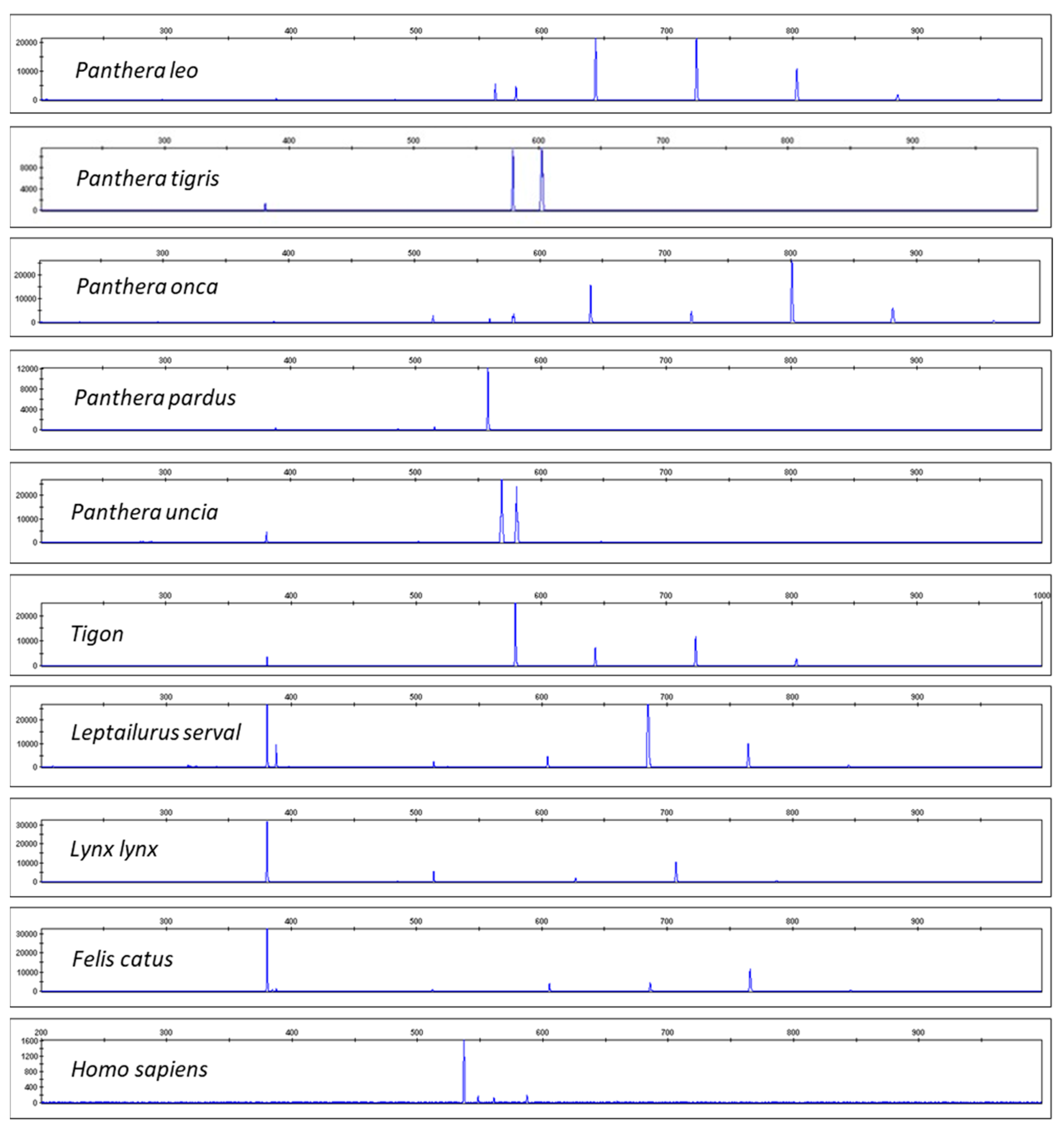
Figure 2 shows the resulting electropherograms for Panthera leo, Panthera tigris, Panthera onca, Panthera pardus, Panthera uncia, Tigon (a combination of P. tigris and P. leo), Leptailurus serval, Lynx lynx, Felis catus, and Homo sapiens. Differences in the resulting CR-mtDNA barcoding profiles should be noted. We tested multiple individuals per species, including the subspecies P. tigris sumatrae, P. tigris jacksoni, P. tigris altaica, and P. tigris ussuri. For details of the samples’ origin see Materials and Methods. The species assignment was also confirmed by Sanger sequencing of COI and cytB mtDNA genes with a subsequent comparison with databases BOLDsystems [38] or GenBank [39]. All resulting DNA CR-mtDNA-length profiles were specific for a given species (see the Supplementary Materials).
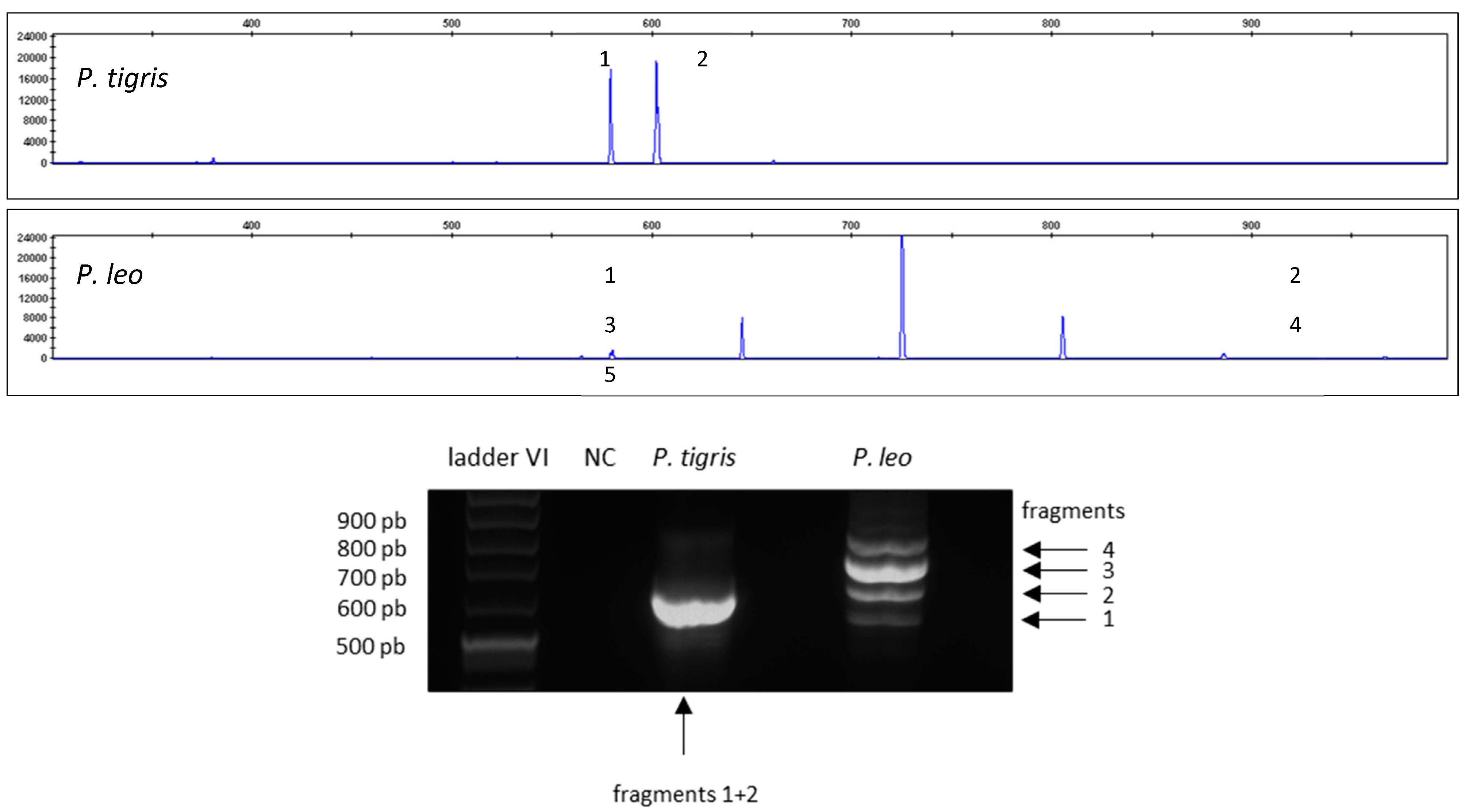
Figure 3 shows the different barcode patterns for P. tigris and P. leo analyzed via capillary electrophoresis (upper panel) and agarose gel electrophoresis (lower panel).
Figure 4 shows the sequence of P. leo CR-mtDNA amplicons 1–4 (see lower panel of Figure 3), which were purified from agarose gels. The start of the 80 bp repetitive sequence (LRS) is marked in yellow.
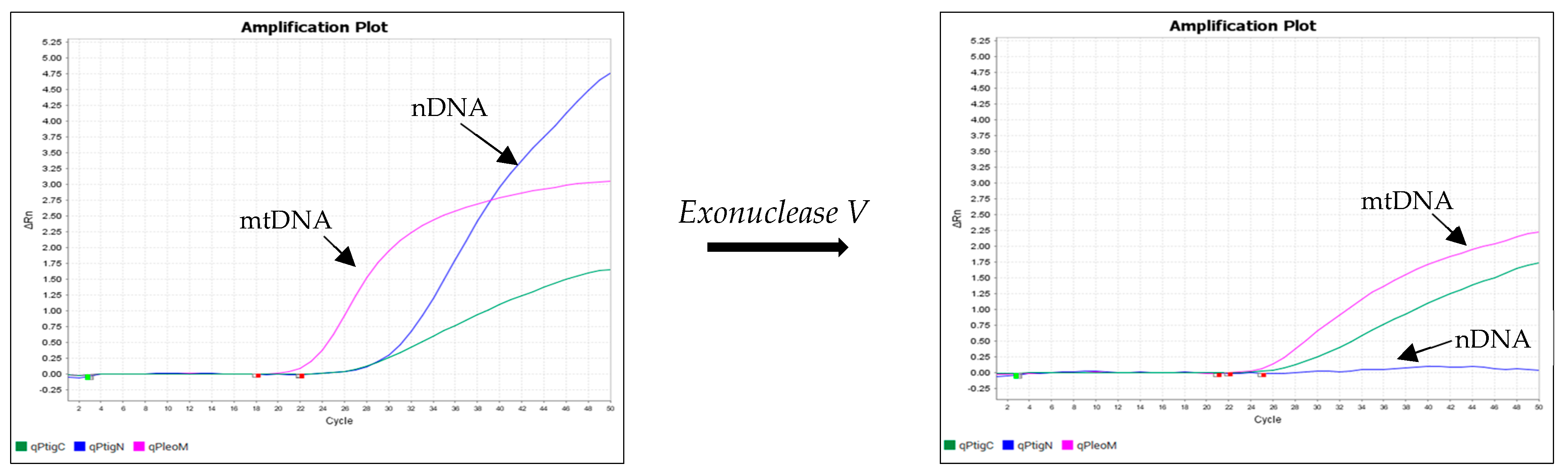
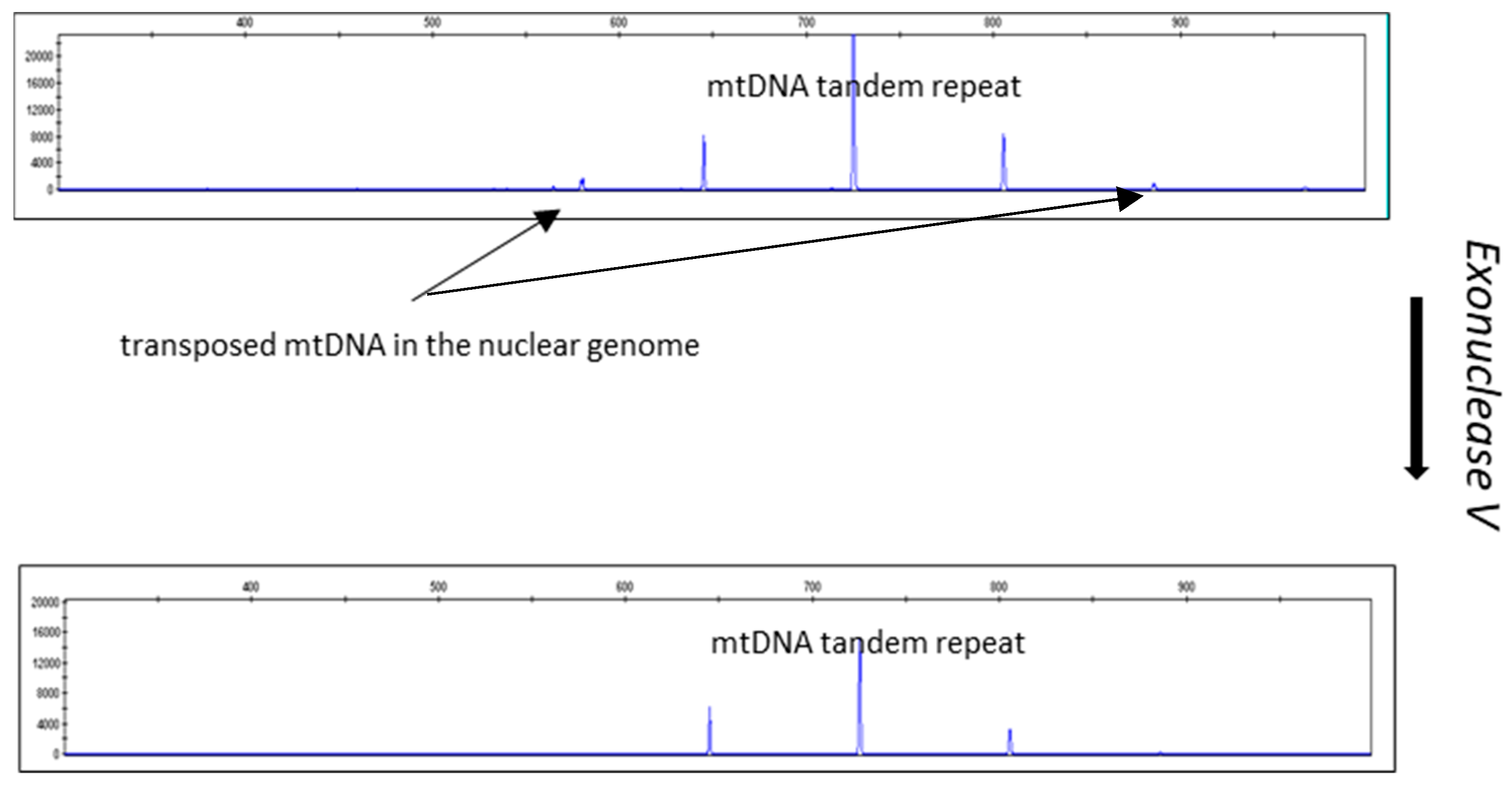
Figure 5 and Figure 6 show the results of an experiment in which the DNA isolates were treated with exonuclease V. The effect of exonuclease V was demonstrated by the multiplex qPCR assay Pleo Qplex (Figure 5), which targets nuclear DNA, mitochondrial DNA, and IPC (internal positive control), or by capillary electrophoresis (Figure 6).
Results of the Validation
Specificity: We applied the assay to DNA extracts of closely related species and species that may co-occur (species regularly processed in a lab) or be substituted for the target species (e.g., in TCMs) (Sus scrofa, Bos taurus, and C. lupus on top of the species listed in Table 1). The CR-mtDNA-length polymorphism assay was found to be species-specific with the following exceptions: testing of P. pardus subspecies produces slightly different patterns and P. onca can be misinterpreted as P. leo if only peak positions (without peak heights) are considered.
Sensitivity: Analytical sensitivity was tested on serial dilutions of DNA from seven target animals frequently occurring in our casework (P. tigris, P. leo, P. onca, P. pardus, L. lynx, H. sapiens, and F. catus), with the DNA input ranging from 2 ng to 5 pg. The validation showed that the method’s sensitivity is applicable to DNA inputs of more than 5 pg nDNA (qPCR quantitation results).
Robustness: We tested the robustness of the assay under temperatures shifted against the original protocol (annealing temperature: 55 °C). Changes in the annealing temperature greater than +2 °C will cause the assay to fail to generate the characteristic CE profile.
Repeatability: DNA extracts from different individuals from the target species (P. tigris (10), P. leo (10), P. pardus (5), P. uncia (3), L. serval (4), Tigon (2), and L. lynx (4)) were used to test the repeatability (see the Supplementary Material). The results were identical for all of the samples tested, except for the P. pardus subspecies which produced slightly different EPGs.
Reproducibility: We tested the within-lab reproducibility with two different analysts independently running replicates of the assay. The resulting EPGs were identical for all of the samples tested.
Negative and positive controls were run in all of the above experiments and produced results as expected.
4. Discussion
Species identification is a crucial aspect of solving criminal cases involving non-human biological materials. Identifying the species can help to narrow down the range of suspects and increase the chances of identifying the true culprit. Currently, there are many techniques based on mtDNA that allow for species identification. However, this relatively streamlined approach, relying on mtDNA typing, becomes challenging when a lab faces a mixture of DNA from different species that can be degraded [40]. When samples contain a mixture of DNA from multiple species, interpreting sequencing analysis results is difficult and requires either a special interpretation framework [41] or a special software deconvolution tool [42]. DNA metabarcoding using next-generation sequencing (NGS) technologies can overcome the DNA mixture problem [43,44,45,46], but it is not suitable for small-scale analyses routinely performed by local forensic laboratories, primarily because it is costly and time-consuming. DNA barcoding of mixed samples using NGS can overestimate the number of species when nuclear mitochondrial pseudogenes are coamplified and sequenced [35,47,48].
PCR analysis enables species barcoding from minute samples, but the number of identifiable taxa is limited when compared with sequencing techniques [40]. The scientific literature describes a relatively small number of species-specific nuclear markers. The banding pattern of the Alu I-digested prepro-gonadotropin-releasing hormone gene is species-specific in Atlantic salmon and brown trout and can be applied for the identification of their hybrids [49]. Trematomid fishes can also be identified using a species-specific nuclear marker [50]. Studies into transposable elements provide some leads for phylogenetic and population genetic investigations [51], but their use is limited. High-resolution melting analysis of the universal ITS2 region of genomic rDNA has been employed using the novel authentication approach for coffee beans and brewed beverages [52]. Unfortunately, none of these nuclear markers would mitigate the limits of the current barcoding system based on mtDNA barcoding. Nuclear markers should be universally applicable across the majority of species and carry sufficient information to discriminate between closely related species [53]. An analysis of metazoan-level universal single-copy orthologs (metazoan USCOs) shows that this approach might also be applicable outside the metazoans [54].
Nuclear inserts of mitochondrial DNA (NUMTs) have been found in major clades of eukaryotic organisms, but the number of NUMTs can significantly vary between different species [55,56,57,58,59], the distribution of NUMTs is lineage-specific, and the presence of NUMTs cannot be known a priori.
Barcoding analysis of mtDNA targets can be biased by NUMTs [35,36]; however, NUMTs can also be used as population genetic markers, e.g., in gorillas [60]. We have demonstrated that exonuclease V treatment can be used to remove nuclear DNA prior to PCR. Other possibilities are to use selective enrichment [61], design the mtDNA barcoding primers in a way that the amplification of NUMTs is greatly reduced [62], use long-range amplification, or use pre-PCR dilution. NUMT sequences are also available in public databases and are wrongly identified as authentic mtDNA [63]. This cannot only cause problems in phylogenetic studies [64] but also in forensic applications [59,65].
5. Conclusions
When a confiscated TCM artifact indicates the presence of a CITES organism, this is considered legal proof of its presence. Unlabeled and suspicious TCM artifacts must be sent to the forensic laboratory for species determination. The described CR-mtDNA polymorphic region can be utilized for species determination when the presence of biological material from big cats is expected or used as a confirmatory test alongside Sanger sequencing, MPS, or non-DNA barcoding methods [66,67,68,69]. The advantage of the CR-mtDNA barcoding system is its ease of use even in laboratories not performing sequencing techniques. The polymorphic CR-mtDNA system has relatively short amplicons, and its main advantage is its high sensitivity and thus suitability even for problematic samples, such as hair or tanned hide. The above-described assay has been successfully applied as a confirmatory test for a numerous number of casework samples. The system is sufficiently informative for big cats and does not show intraspecies variability. However, it is necessary to point out that the testing of P. pardus subspecies produces slightly different patterns, and P. onca can be misinterpreted as P. leo if only peak positions (without peak heights) are considered. The greatest disadvantage of mtDNA species barcoding assays is that they do not enable the differentiation of hybrids (for instance, tigon = male tiger and female lion) but rather the female line only [70]. CR-mtDNA-length polymorphism typing, as described herein, can differentiate not only different species of big cats but also hybrids (see Figure 2). Species identification is an important component of forensic science, and the development of new techniques or the upgrading of existing ones will continue to improve the accuracy and efficiency of species identification.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/life14040497/s1, Panthera leo–samples 1–12; Panthera tigris—samples 1–12; Panthera pardus—samples 1–4; Panthera onca—samples 1–2; Panthera uncia—samples 1–2; Tigon—samples 1–2; Lyns lynx—samples 1–4; Leptailurus serval—samples 1–4; Homo sapiens—samples 1–2.
Author Contributions
L.V.: conceptualization; methodology; data curation; formal analysis; investigation; writing—original draft. D.V.: conceptualization; funding acquisition; methodology; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
The research was covered by the project “An effective use of forensic methods in wildlife crime investigation” (Ministry of Interior, Czech Republic, VJ01010026).
Institutional Review Board Statement
The reference material used for this study was provided by the Czech Environmental Inspectorate and zoological gardens. The biological material came either from animals that died from natural causes in zoological gardens, or their excrements. The sampling did not involve the infliction of trauma to living animals. The research thus did not fall under the Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes.
Informed Consent Statement
Human DNA samples (laboratory staff) were sampled and used in accordance with the Regulation (EU) 2016/679 of the European Parliament and of the Council of 27 April 2016 on the protection of natural persons with regard to the processing of personal data and on the free movement of such data, following Directive 95/46/EC (General Data Protection Regulation).
Data Availability Statement
Data are contained within the Supplementary Materials.
Acknowledgments
The reference samples were kindly provided by the Czech Environmental Inspectorate and various zoos.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cheung, H.; Doughty, H.; Hinsley, A.; Hsu, E.; Lee, T.M.; Milner-Gulland, E.; Possingham, H.P.; Biggs, D. Understanding Traditional Chinese Medicine to strengthen conservation outcomes. People Nat. 2021, 3, 115–128. [Google Scholar] [CrossRef]
- Secretariat, C.; de l’Environnement, M.I. Convention on International Trade in Endangered Species of Wild Fauna and Flora; Citeseer: Princeton, NJ, USA, 2011. [Google Scholar]
- Pečnikar, Ž.F.; Buzan, E.V. 20 years since the introduction of DNA barcoding: From theory to application. J. Appl. Genet. 2014, 55, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Valentini, A.; Pompanon, F.; Taberlet, P. DNA barcoding for ecologists. Trends Ecol. Evol. 2009, 24, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, Z.; Li, C.; Ping, X.; Cui, S.; Tang, S.; Chu, H.; Liu, B. Identification of ungulates used in a traditional Chinese medicine with DNA barcoding technology. Ecol. Evol. 2015, 5, 1818–1825. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Gan, S.; Zhang, J.; Fan, Y.; Li, B.; Wan, L.; Nie, J.; Wang, X.; Chen, J. Application of DNA Barcoding for the Identification of Snake Gallbladders as a Traditional Chinese Medicine. Rev. Bras. Farmacogn. 2022, 32, 663–668. [Google Scholar] [CrossRef]
- Zhang, H.; Yao, H.; Cui, L.; Du, H.; Lin, Z.; Gao, X.; Lang, X.; Song, J.; Luo, K.; Shi, L.; et al. Application of COI-based DNA barcoding for identifying animal medical materials in the Chinese pharmacopoeia. World Sci. Technol. Mod. Tradit. Chin. Med. 2013, 12, 371–380. [Google Scholar]
- Yang, F.; Ding, F.; Chen, H.; He, M.; Zhu, S.; Ma, X.; Jiang, L.; Li, H. DNA barcoding for the identification and authentication of animal species in traditional medicine. Evid. Based Complement. Altern. Med. 2018, 2018, 5160254. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Sun, W.; Li, J.; Yao, H.; Shi, Y.; Wang, P.; Huang, B.; Shi, L.; Liu, D.; Hu, Z. Identifying the species of seeds in traditional Chinese medicine using DNA barcoding. Front. Pharmacol. 2018, 9, 701. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Liu, Q.; Qiu, S.; Dai, J.; Gao, X. DNA barcoding: An efficient technology to authenticate plant species of traditional Chinese medicine and recent advances. Chin. Med. 2022, 17, 112. [Google Scholar] [CrossRef]
- Gong, L.; Qiu, X.H.; Huang, J.; Xu, W.; Bai, J.Q.; Zhang, J.; Su, H.; Xu, C.M.; Huang, Z.H. Constructing a DNA barcode reference library for southern herbs in China: A resource for authentication of southern Chinese medicine. PLoS ONE 2018, 13, e0201240. [Google Scholar] [CrossRef]
- Miao, L.; Xi-Wen, L.; Bao-Sheng, L.; Lu, L.; Yue-Ying, R. Species identification of poisonous medicinal plant using DNA barcoding. Chin. J. Nat. Med. 2019, 17, 585–590. [Google Scholar]
- Menotti-Raymond, M.; David, V.A.; Stephens, J.C.; Lyons, L.A.; O’Brien, S.J. Genetic individualization of domestic cats using feline STR loci for forensic applications. J. Forensic Sci. 1997, 42, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, A.P.; Rohleder, U.; Eichmann, C.; Pfeiffer, I.; Parson, W.; Schleenbecker, U. A proposal for standardization in forensic canine DNA typing: Allele nomenclature of six canine-specific STR loci. J. Forensic Sci. 2006, 51, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Potoczniak, M.J.; Chermak, M.; Quarino, L.; Tobe, S.S.; Conte, J. Development of a multiplex, PCR-based genotyping assay for African and Asian elephants for forensic purposes. Int. J. Leg. Med. 2020, 134, 55–62. [Google Scholar] [CrossRef]
- Singh, A.; Priyambada, P.; Jabin, G.; Singh, S.K.; Joshi, B.D.; Venkatraman, C.; Chandra, K.; Sharma, L.K.; Thakur, M. Pangolin Indexing System: Implications in forensic surveillance of large seizures. Int. J. Leg. Med. 2020, 134, 1613–1618. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.K. RhODIS®(The Rhinoceros DNA Index System): The Application of Simple Forensic and Genetic Tools Help Conserve African Rhinoceros. In Wildlife Biodiversity Conservation; Springer: Berlin/Heidelberg, Germany, 2021; pp. 463–485. [Google Scholar]
- Vaněk, D.; Ehler, E.; Vaňková, L. Development of DNA quantitation and STR typing systems for Panthera tigris species determination and individual identification in forensic casework. Eur. J. Environ. Sci. 2021, 11, 113–118. [Google Scholar] [CrossRef]
- Wilkinson, M.J.; Szabo, C.; Ford, C.S.; Yarom, Y.; Croxford, A.E.; Camp, A.; Gooding, P. Replacing Sanger with Next Generation Sequencing to improve coverage and quality of reference DNA barcodes for plants. Sci. Rep. 2017, 7, 46040. [Google Scholar] [CrossRef]
- Haider, N.; Nabulsi, I.; Al-Safadi, B. Identification of meat species by PCR-RFLP of the mitochondrial COI gene. Meat Sci. 2012, 90, 490–493. [Google Scholar] [CrossRef]
- Noikotr, K.; Chaveerach, A.; Pinthong, K.; Tanomtong, A.; Sudmoon, R.; Tanee, T. RAPD and barcode analyses of groupers of the genus Epinephelus. Genet. Mol. Res 2013, 12, 5721–5732. [Google Scholar] [CrossRef]
- Hoffman, J.; Clark, M.; Amos, W.; Peck, L. Widespread amplification of amplified fragment length polymorphisms (AFLPs) in marine Antarctic animals. Polar Biol. 2012, 35, 919–929. [Google Scholar] [CrossRef]
- Lahiff, S.; Glennon, M.; Lyng, J.; Smith, T.; Maher, M.; Shilton, N. Species-specific PCR for the identification of ovine, porcine and chicken species in meat and bone meal (MBM). Mol. Cell. Probes 2001, 15, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Shin, S.E.; Ko, K.S.; Park, S.H. Identification of forensically important Calliphoridae and Sarcophagidae species collected in Korea using SNaPshot multiplex system targeting the cytochrome c oxidase subunit i gene. BioMed Res. Int. 2018, 2018, 2953892. [Google Scholar] [CrossRef] [PubMed]
- Denyingyhot, A.; Phraephaisarn, C.; Vesaratchavest, M.; Dahlan, W.; Keeratipibul, S. A new tool for quality control to monitor contamination of six non-halal meats in food industry by multiplex high-resolution melting analysis (HRMA). NFS J. 2021, 25, 31–40. [Google Scholar] [CrossRef]
- Friedenberger, A.; Doyle, C.; Couillard, L.; Kyle, C.J. The bear necessities: A sensitive qPCR assay for bear DNA detection from bile and derived products to complement wildlife forensic enforcement. Forensic Sci. Int. Genet. 2023, 67, 102935. [Google Scholar] [CrossRef] [PubMed]
- Mori, C.; Matsumura, S. Development and validation of simultaneous identification of 26 mammalian and poultry species by a multiplex assay. Int. J. Leg. Med. 2021, 136, 1–12. [Google Scholar] [CrossRef]
- Pereira, F.; Carneiro, J.; Matthiesen, R.; van Asch, B.; Pinto, N.; Gusmao, L.; Amorim, A. Identification of species by multiplex analysis of variable-length sequences. Nucleic Acids Res. 2010, 38, e203. [Google Scholar] [CrossRef] [PubMed]
- Pun, K.M.; Albrecht, C.; Castella, V.; Fumagalli, L. Species identification in mammals from mixed biological samples based on mitochondrial DNA control region length polymorphism. Electrophoresis 2009, 30, 1008–1014. [Google Scholar] [CrossRef]
- Vankova, L.; Vanek, D. DNA-based identification of big cats and traditional Chinese medicine artifacts in the Czech Republic. Forensic Sci. Int. Genet. Suppl. Ser. 2022, 8, 122–124. [Google Scholar] [CrossRef]
- Warchol, G.L. The transnational illegal wildlife trade. In Transnational Environmental Crime; Routledge: London, UK, 2017; pp. 379–396. [Google Scholar]
- Petrossian, G.A.; Pires, S.F.; van Uhm, D.P. An overview of seized illegal wildlife entering the United States. Glob. Crime 2016, 17, 181–201. [Google Scholar] [CrossRef]
- Bagatharia, S.B.; Joshi, M.N.; Pandya, R.V.; Pandit, A.S.; Patel, R.P.; Desai, S.M.; Sharma, A.; Panchal, O.; Jasmani, F.P.; Saxena, A.K. Complete mitogenome of asiatic lion resolves phylogenetic status within Panthera. BMC Genom. 2013, 14, 572. [Google Scholar] [CrossRef]
- Lopez, J.V.; Cevario, S.; O’Brien, S.J. Complete nucleotide sequences of the domestic cat (Felis catus) mitochondrial genome and a transposed mtDNA tandem repeat (Numt) in the nuclear genome. Genomics 1996, 33, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Natl. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef]
- Zhang, D.-X.; Hewitt, G.M. Nuclear integrations: Challenges for mitochondrial DNA markers. Trends Ecol. Evol. 1996, 11, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Shankaranarayanan, P.; Banerjee, M.; Kacker, R.K.; Aggarwal, R.K.; Singh, L. Genetic variation in Asiatic lions and Indian tigers. Electrophoresis 1997, 18, 1693–1700. [Google Scholar] [CrossRef]
- Webster, L.M.; Prigge, T.-L.; Frankham, G.J. A guide for the validation of DNA based species identification in forensic casework. Forensic Sci. Int. Anim. Environ. 2024, 5, 100080. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2012, 41, D36–D42. [Google Scholar] [CrossRef]
- Mori, C.; Matsumura, S. Current issues for mammalian species identification in forensic science: A review. Int. J. Leg. Med. 2021, 135, 3–12. [Google Scholar] [CrossRef]
- Egeland, T.; Salas, A. A statistical framework for the interpretation of mtDNA mixtures: Forensic and medical applications. PLoS ONE 2011, 6, e26723. [Google Scholar] [CrossRef]
- Mandape, S.N.; Smart, U.; King, J.L.; Muenzler, M.; Kapema, K.B.; Budowle, B.; Woerner, A.E. MMDIT: A tool for the deconvolution and interpretation of mitochondrial DNA mixtures. Forensic Sci. Int. Genet. 2021, 55, 102568. [Google Scholar] [CrossRef]
- Holland, M.M.; McQuillan, M.R.; O’Hanlon, K.A. Second generation sequencing allows for mtDNA mixture deconvolution and high resolution detection of heteroplasmy. Croat. Med. J. 2011, 52, 299–313. [Google Scholar] [CrossRef]
- Kim, H.; Erlich, H.A.; Calloway, C.D. Analysis of mixtures using next generation sequencing of mitochondrial DNA hypervariable regions. Croat. Med. J. 2015, 56, 208–217. [Google Scholar] [CrossRef]
- Churchill, J.D.; Stoljarova, M.; King, J.L.; Budowle, B. Massively parallel sequencing-enabled mixture analysis of mitochondrial DNA samples. Int. J. Leg. Med. 2018, 132, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Wisner, M.; Erlich, H.; Shih, S.; Calloway, C. Resolution of mitochondrial DNA mixtures using a probe capture next generation sequencing system and phylogenetic-based software. Forensic Sci. Int. Genet. 2021, 53, 102531. [Google Scholar] [CrossRef]
- Schultz, J.A.; Hebert, P.D. Do pseudogenes pose a problem for metabarcoding marine animal communities? Mol. Ecol. Resour. 2022, 22, 2897–2914. [Google Scholar] [CrossRef]
- Cruaud, P.; Rasplus, J.-Y.; Rodriguez, L.J.; Cruaud, A. High-throughput sequencing of multiple amplicons for barcoding and integrative taxonomy. Sci. Rep. 2017, 7, 41948. [Google Scholar] [CrossRef]
- Gross, R.; Nilsson, J.; Schmitz, M. A new species-specific nuclear DNA marker for identification of hybrids between Atlantic salmon and brown trout. J. Fish Biol. 1996, 49, 537–540. [Google Scholar]
- Van de Putte, A.P.; Van Houdt, J.; Maes, G.; Janko, K.; Koubbi, P.; Rock, J.; Volckaert, F. Species identification in the trematomid family using nuclear genetic markers. Polar Biol. 2009, 32, 1731–1741. [Google Scholar] [CrossRef]
- Purugganan, M.; Wessler, S. Transposon signatures: Species-specific molecular markers that utilize a class of multiple-copy nuclear DNA. Mol. Ecol. 1995, 4, 265–270. [Google Scholar] [CrossRef]
- Bosmali, I.; Lagiotis, G.; Stavridou, E.; Haider, N.; Osathanunkul, M.; Pasentsis, K.; Madesis, P. Novel authentication approach for coffee beans and the brewed beverage using a nuclear-based species-specific marker coupled with high resolution melting analysis. LWT 2021, 137, 110336. [Google Scholar] [CrossRef]
- Eberle, J.; Ahrens, D.; Mayer, C.; Niehuis, O.; Misof, B. A plea for standardized nuclear markers in metazoan DNA taxonomy. Trends Ecol. Evol. 2020, 35, 336–345. [Google Scholar] [CrossRef]
- Dietz, L.; Eberle, J.; Mayer, C.; Kukowka, S.; Bohacz, C.; Baur, H.; Espeland, M.; Huber, B.A.; Hutter, C.; Mengual, X. Standardized nuclear markers improve and homogenize species delimitation in Metazoa. Methods Ecol. Evol. 2023, 14, 543–555. [Google Scholar] [CrossRef]
- Pereira, S.L.; Baker, A.J. Low number of mitochondrial pseudogenes in the chicken (Gallus gallus) nuclear genome: Implications for molecular inference of population history and phylogenetics. BMC Evol. Biol. 2004, 4, 17. [Google Scholar] [CrossRef]
- Nacer, D.F.; do Amaral, F.R. Striking pseudogenization in avian phylogenetics: Numts are large and common in falcons. Mol. Phylogenetics Evol. 2017, 115, 1–6. [Google Scholar] [CrossRef]
- Richly, E.; Leister, D. NUMTs in sequenced eukaryotic genomes. Mol. Biol. Evol. 2004, 21, 1081–1084. [Google Scholar] [CrossRef]
- Hazkani-Covo, E.; Zeller, R.M.; Martin, W. Molecular poltergeists: Mitochondrial DNA copies (numts) in sequenced nuclear genomes. PLoS Genet. 2010, 6, e1000834. [Google Scholar] [CrossRef]
- Cihlar, J.C.; Strobl, C.; Lagacé, R.; Muenzler, M.; Parson, W.; Budowle, B. Distinguishing mitochondrial DNA and NUMT sequences amplified with the precision ID mtDNA whole genome panel. Mitochondrion 2020, 55, 122–133. [Google Scholar] [CrossRef]
- Soto-Calderón, I.D.; Clark, N.J.; Wildschutte, J.V.H.; DiMattio, K.; Jensen-Seaman, M.I.; Anthony, N.M. Identification of species-specific nuclear insertions of mitochondrial DNA (numts) in gorillas and their potential as population genetic markers. Mol. Phylogenetics Evol. 2014, 81, 61–70. [Google Scholar] [CrossRef]
- Wolff, J.N.; Shearman, D.C.; Brooks, R.C.; Ballard, J.W. Selective enrichment and sequencing of whole mitochondrial genomes in the presence of nuclear encoded mitochondrial pseudogenes (numts). PLoS ONE 2012, 7, e37142. [Google Scholar] [CrossRef]
- Morgan, K.I.; Ewart, K.M.; Nguyen, T.Q.; Sitam, F.T.; Ouitavon, K.; Lightson, A.L.; Kotze, A.; McEwing, R. Avoiding common numts to provide reliable species identification for tiger parts. Forensic Sci. Int. Rep. 2021, 3, 100166. [Google Scholar] [CrossRef]
- Kunz, D.; Tay, W.T.; Elfekih, S.; Gordon, K.H.J.; De Barro, P.J. Take out the rubbish–Removing NUMTs and pseudogenes from the Bemisia tabaci cryptic species mtCOI database. bioRxiv 2019. [Google Scholar] [CrossRef]
- Ožana, S.; Dolný, A.; Pánek, T. Nuclear copies of mitochondrial DNA as a potential problem for phylogenetic and population genetic studies of Odonata. Syst. Entomol. 2022, 47, 591–602. [Google Scholar] [CrossRef]
- Marshall, C.; Parson, W. Interpreting NUMTs in forensic genetics: Seeing the forest for the trees. Forensic Sci. Int. Genet. 2021, 53, 102497. [Google Scholar] [CrossRef] [PubMed]
- Rossel, S.; Uhlenkott, K.; Peters, J.; Vink, A.; Arbizu, P.M. Evaluating species richness using proteomic fingerprinting and DNA barcoding—A case study on meiobenthic copepods from the Clarion Clipperton Fracture Zone. Mar. Biodivers. 2022, 52, 67. [Google Scholar] [CrossRef]
- Raupach, M.J.; Amann, R.; Wheeler, Q.D.; Roos, C. The application of “-omics” technologies for the classification and identification of animals. Org. Divers. Evol. 2016, 16, 1–12. [Google Scholar] [CrossRef]
- Davidson, N.B.; Koch, N.I.; Sarsby, J.; Jones, E.; Hurst, J.L.; Beynon, R.J. Rapid identification of species, sex and maturity by mass spectrometric analysis of animal faeces. BMC Biol. 2019, 17, 66. [Google Scholar] [CrossRef]
- Zhong, Y.; Wang, H.; Wei, Q.; Cao, R.; Zhang, H.; He, Y.; Wang, L. Combining DNA barcoding and HPLC fingerprints to trace species of an important traditional Chinese medicine fritillariae bulbus. Molecules 2019, 24, 3269. [Google Scholar] [CrossRef]
Figure 1.
Structure of the control region (CR) of the Panthera tigris isolate PTI. The CR of P. tigris isolate PTI is located between tRNA-Pro and tRNA-Phe and is 1345 bp long (position 16 187-738). CR is divided into 3 parts: the left domain, with a long repetitive sequence of 80 bp (LRS; position 16 385-16 545); the central conserved domain (position 16 546-16 793, 1-273); and the right domain, with a short repeat sequence of 8 bp (SRS; position 273-377) (based on Bagatharia [33] and Lopez [34]).
Figure 1.
Structure of the control region (CR) of the Panthera tigris isolate PTI. The CR of P. tigris isolate PTI is located between tRNA-Pro and tRNA-Phe and is 1345 bp long (position 16 187-738). CR is divided into 3 parts: the left domain, with a long repetitive sequence of 80 bp (LRS; position 16 385-16 545); the central conserved domain (position 16 546-16 793, 1-273); and the right domain, with a short repeat sequence of 8 bp (SRS; position 273-377) (based on Bagatharia [33] and Lopez [34]).

Figure 2.
CR-mtDNA electropherograms for P. leo, P. tigris, Panthera onca, Panthera pardus, Panthera uncia, Tigon (hybrid of P. tigris and P. leo), Leptailurus serval, Lynx lynx, Felis catus, and Homo sapiens. Note that all of the species (including the hybrid tiger–lion species) have distinct barcodes.
Figure 2.
CR-mtDNA electropherograms for P. leo, P. tigris, Panthera onca, Panthera pardus, Panthera uncia, Tigon (hybrid of P. tigris and P. leo), Leptailurus serval, Lynx lynx, Felis catus, and Homo sapiens. Note that all of the species (including the hybrid tiger–lion species) have distinct barcodes.

Figure 3.
Resulting electropherograms for CR-mtDNA-length polymorphism amplifications of P. tigris and P. leo analyzed via capillary electrophoresis (upper panel) and agarose gel electrophoresis (lower panel). Peak 5 is not visible on the agarose gel due to lower sensitivity.
Figure 3.
Resulting electropherograms for CR-mtDNA-length polymorphism amplifications of P. tigris and P. leo analyzed via capillary electrophoresis (upper panel) and agarose gel electrophoresis (lower panel). Peak 5 is not visible on the agarose gel due to lower sensitivity.
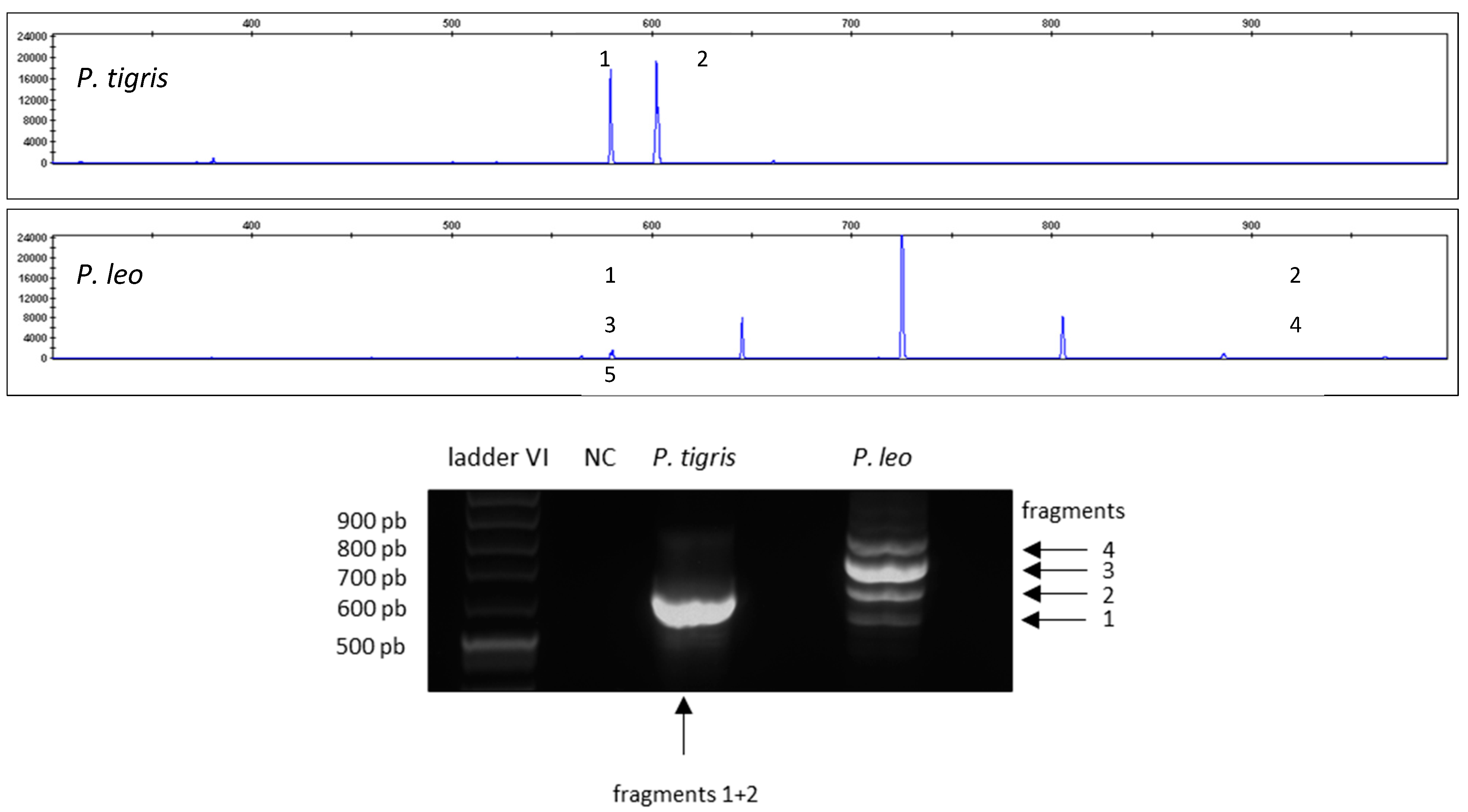
Figure 4.
The sequence of P. leo CR-mtDNA-length polymorphism amplicons 1–4 (see lower panel of Figure 3), purified from agarose gel. The start of the 80 bp repetitive sequence is marked in yellow.
Figure 4.
The sequence of P. leo CR-mtDNA-length polymorphism amplicons 1–4 (see lower panel of Figure 3), purified from agarose gel. The start of the 80 bp repetitive sequence is marked in yellow.

Figure 5.
Effect of exonuclease V treatment on DNA isolates, as demonstrated by the multiplex qPCR assay Pleo Qplex, which targets nuclear DNA (blue), mitochondrial DNA (pink), and internal positive control DNA (green).
Figure 5.
Effect of exonuclease V treatment on DNA isolates, as demonstrated by the multiplex qPCR assay Pleo Qplex, which targets nuclear DNA (blue), mitochondrial DNA (pink), and internal positive control DNA (green).
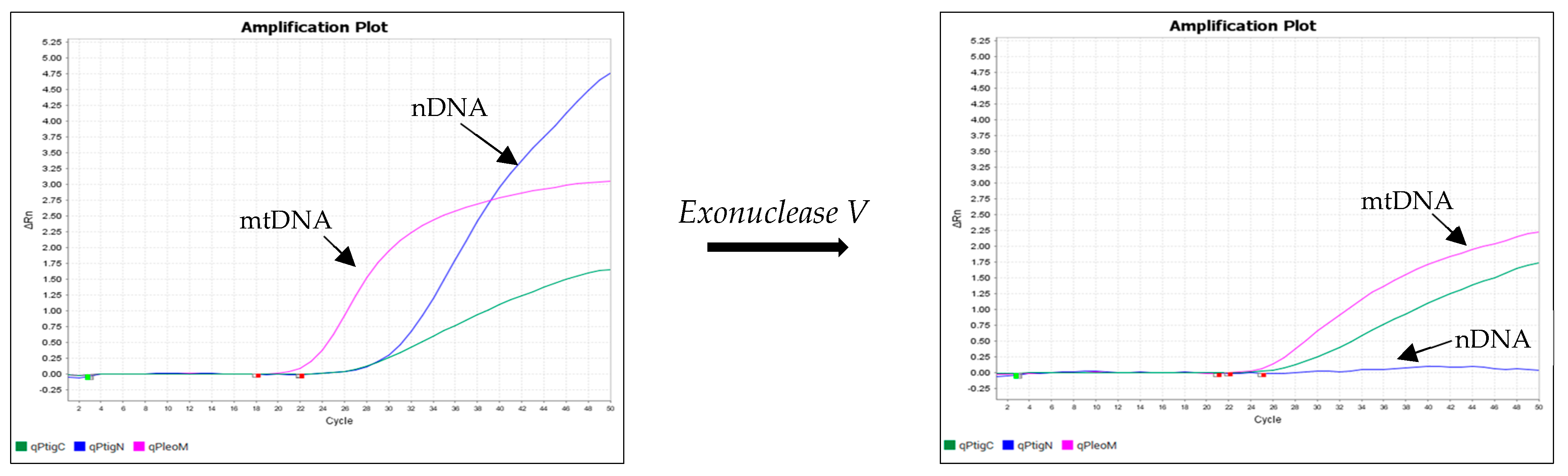
Figure 6.
Effect of exonuclease V treatment on DNA isolates visualized by capillary electrophoresis of CR-mtDNA-length polymorphism P. leo amplicons. Please note the disappearance of the NUMTs’ peaks (1 and 5). The sample used for the exonuclease V treatment differs from the sample used for the CR-mtDNA electropherograms shown in Figure 2. The mtDNA peaks remaining after ExoV treatment are species-specific.
Figure 6.
Effect of exonuclease V treatment on DNA isolates visualized by capillary electrophoresis of CR-mtDNA-length polymorphism P. leo amplicons. Please note the disappearance of the NUMTs’ peaks (1 and 5). The sample used for the exonuclease V treatment differs from the sample used for the CR-mtDNA electropherograms shown in Figure 2. The mtDNA peaks remaining after ExoV treatment are species-specific.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Species analyzed in this study.
| Species | No. of Samples Tested | Sample Type (Number) |
|---|---|---|
| Panthera leo | 30 | hair (15), blood (15) |
| Panthera tigris | 30 | hair (15), tissue (5), blood (5), excrement (5) |
| Panthera onca | 2 | hair (2) |
| Panthera pardus | 10 | hair (7), blood (3) |
| Panthera uncia | 3 | hair (3) |
| Tigon (P. tigris x P. leo) | 2 | hair (2) |
| Leptailurus serval | 4 | hair (4) |
| Lynx lynx | 4 | hair (2), excrement (2) |
| Felis catus | 2 | buccal swab (2) |
| Homo sapiens | 5 | buccal swab (5) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vankova, L.; Vanek, D. Capillary-Electrophoresis-Based Species Barcoding of Big Cats: CR-mtDNA-Length Polymorphism. Life 2024, 14, 497. https://doi.org/10.3390/life14040497
AMA Style
Vankova L, Vanek D. Capillary-Electrophoresis-Based Species Barcoding of Big Cats: CR-mtDNA-Length Polymorphism. Life. 2024; 14(4):497. https://doi.org/10.3390/life14040497
Chicago/Turabian StyleVankova, Lenka, and Daniel Vanek. 2024. "Capillary-Electrophoresis-Based Species Barcoding of Big Cats: CR-mtDNA-Length Polymorphism" Life 14, no. 4: 497. https://doi.org/10.3390/life14040497
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.