
Genome-Wide Identification, Evolution, and Female-Biased Expression Analysis of Odorant Receptors in Tuta absoluta (Lepidoptera: Gelechiidae)

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of OR Genes in T. absoluta
2.2. Gene Structure Analysis and Chromosomal Location of OR Genes
2.3. Phylogenetic Analysis
2.4. Expression Profiles of 58 TabsORs in Female and Male Antennae
2.5. RT-qPCR Verification of the Candidate Female-Antennae-Biased Expression ORs
2.6. Statistical Analysis
3. Results
3.1. Identification and Gene Structure Analysis of OR Genes in T. absoluta
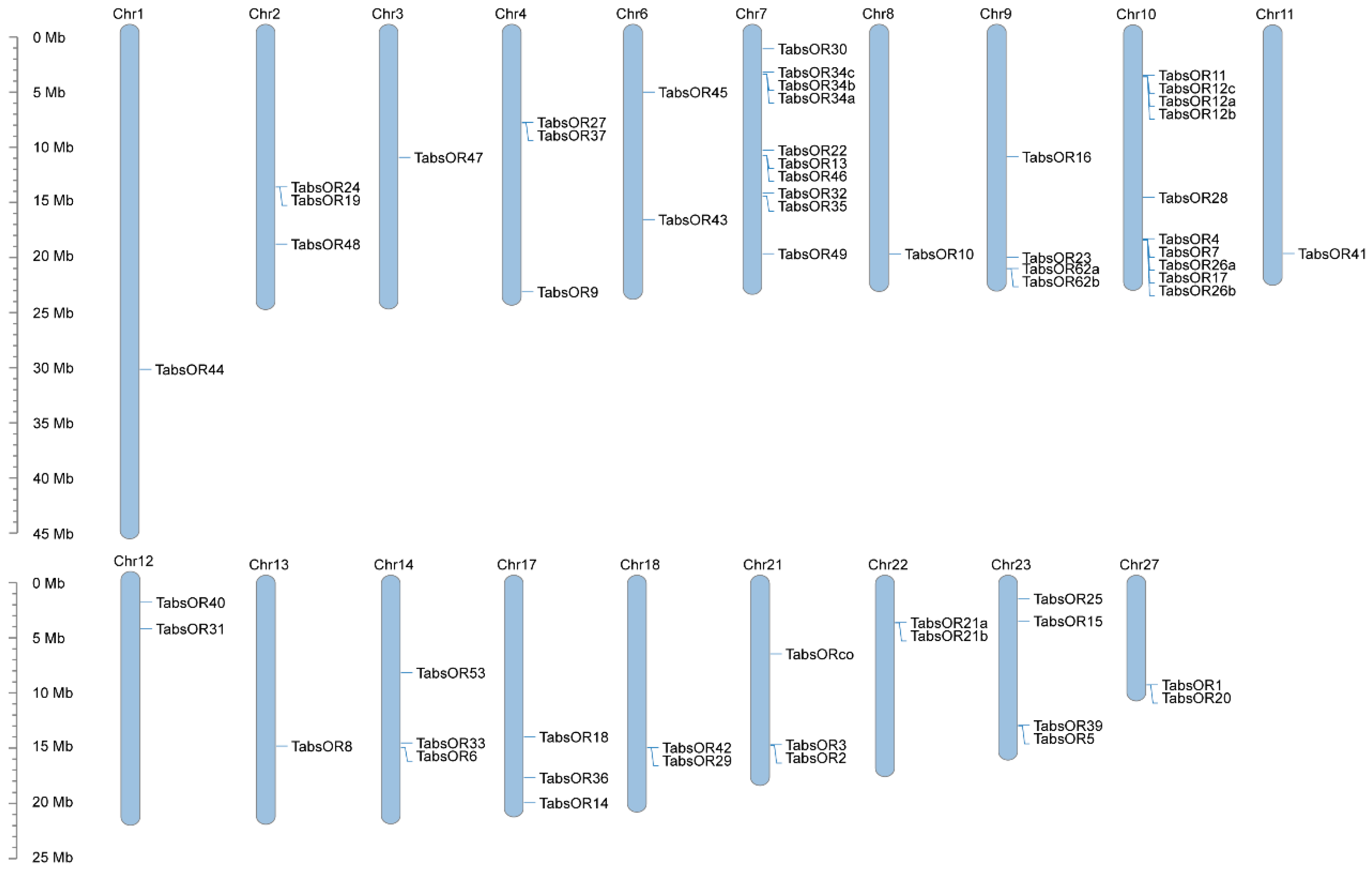
3.2. Chromosome Location of TabsOR Genes
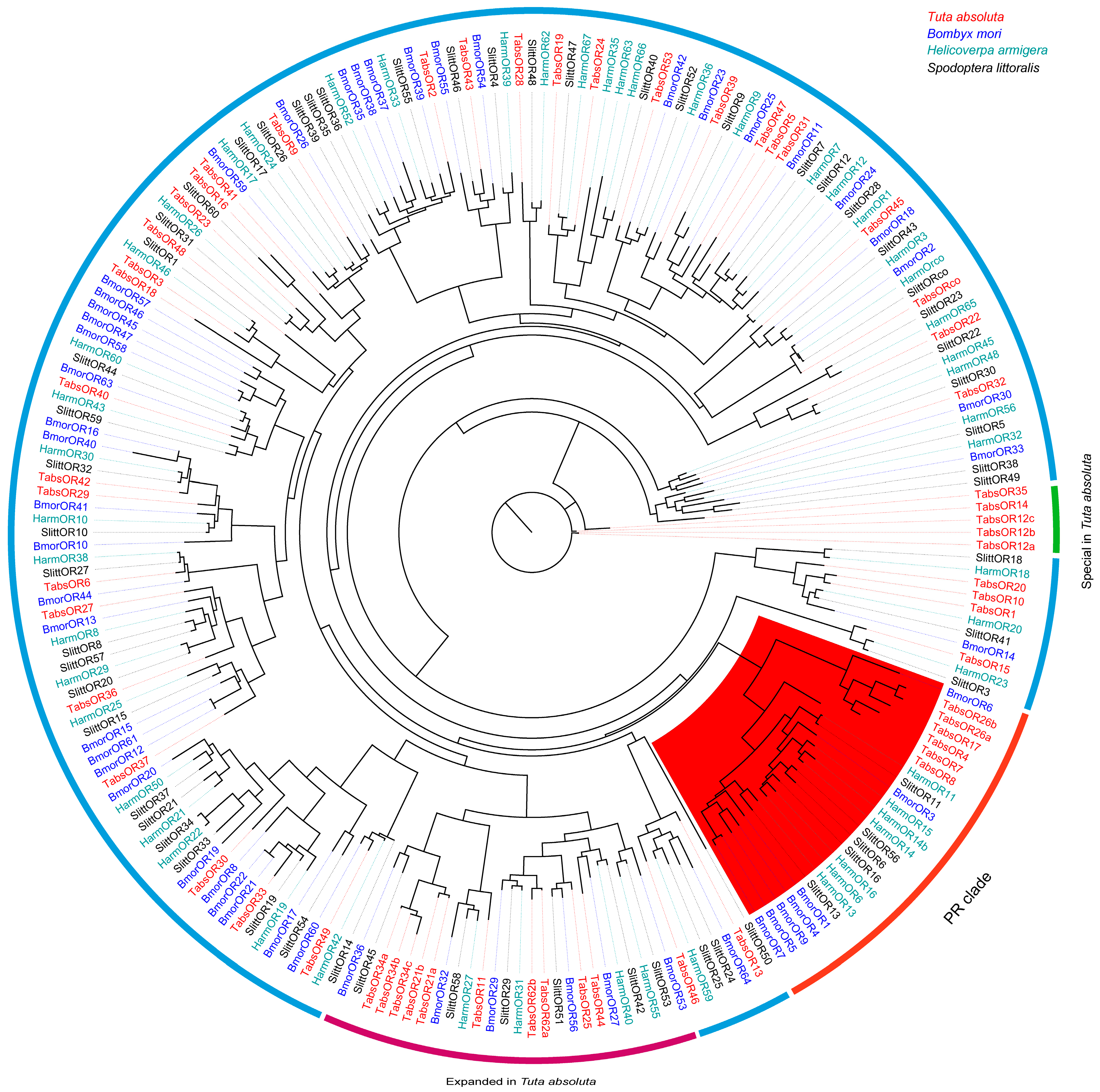
3.3. Phylogenetic Analysis of ORs
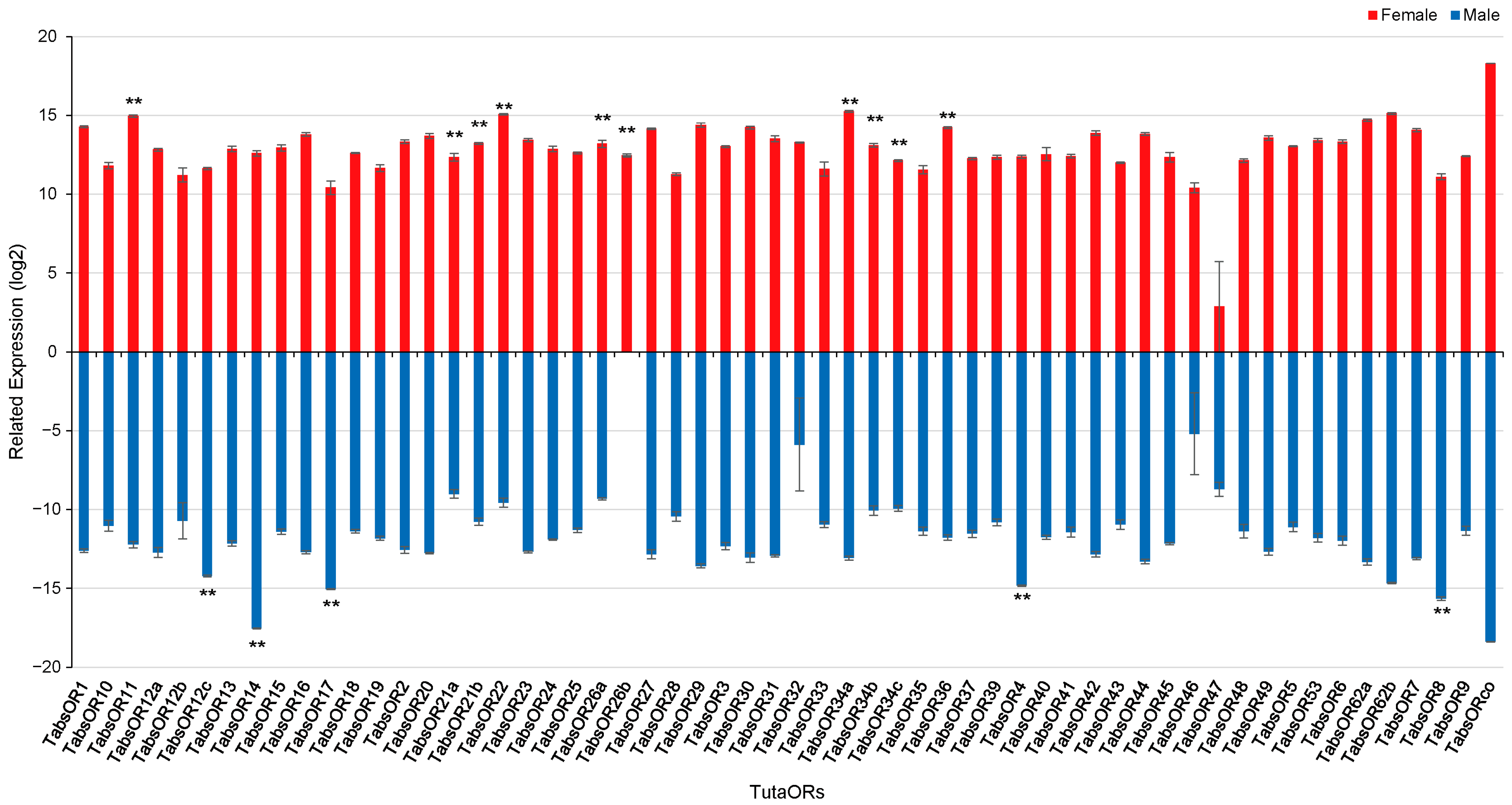
3.4. Expression of TabsOR Genes in Antennae of Female and Male Adults
3.5. RT-qPCR Analysis of Candidate Female-Antennae-Biased Expression ORs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carrasco, D.; Larsson, M.C.; Anderson, P. Insect host plant selection in complex environments. Curr. Opin. Insect Sci. 2015, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Knolhoff, L.M.; Heckel, D.G. Behavioral assays for studies of host plant choice and adaptation in Herbivorous insects. Annu. Rev. Entomol. 2014, 59, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Shi, C.; Wang, B.; Xiu, P.; Wang, Y.; Liu, Y.; Wang, G.R. Evolutionary shifts in pheromone receptors contribute to speciation in four species. Cell. Mol. Life Sci. 2023, 80, 199. [Google Scholar] [CrossRef] [PubMed]
- Tom, M.T.; Llorca, L.C.; Bucks, S.; Bisch-Knaden, S.; Hansson, B.S. Sex- and tissue-specific expression of chemosensory receptor genes in a hawkmoth. Front. Ecol. Evol. 2022, 10, 976521. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Guo, J.M.; Wei, Z.Q.; Zhang, X.T.; Liu, S.R.; Guo, H.F.; Dong, S.L. Identification and sex expression profiles of olfactory-related genes in Mythimna loreyi based on antennal transcriptome analysis. J. Asia-Pac. Entomol. 2022, 25, 101934. [Google Scholar] [CrossRef]
- An, X.; Khashaveh, A.; Liu, D.; Xiao, Y.; Wang, Q.; Wang, S.; Geng, T.; Gu, S.; Zhang, Y. Functional characterization of one sex pheromone receptor (AlucOR4) in Apolygus lucorum (Meyer-Dur). J. Insect Physiol. 2020, 120, 103986. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.M.; Liu, X.L.; Liu, S.R.; Wei, Z.Q.; Han, W.K.; Guo, Y.; Dong, S.L. Functional characterization of sex pheromone receptors in the fall armyworm (Spodoptera frugiperda). Insects 2020, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Mo, B.T.; Li, G.C.; Li, Z.L.; Huang, L.Q.; Sun, Y.L.; Dong, J.F.; Smith, D.P.; Wang, C.Z. Sex pheromone communication in an insect parasitoid, Campoletis chlorideae Uchida. Proc. Natl. Acad. Sci. USA 2022, 119, e2215442119. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dong, H.; Qu, Y.; Zhou, Y.; Qin, J.; Li, K.; Luo, C.; Ren, B.; Cao, Y.; Zhang, S.; et al. Circabidian rhythm of sex pheromone reception in a scarab beetle. Curr. Biol. 2024, 34, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.N.; Corcoran, J.A.; Zhang, D.D.; Hillbur, Y.; Newcomb, R.D.; Lofstedt, C. A sex pheromone receptor in the Hessian fly Mayetiola destructor (Diptera, Cecidomyiidae). Front. Cell. Neurosci. 2016, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Zhao, T.; Lv, H.X.; Li, M.J.; Xing, D.; Zhao, T.Y.; Li, C.X. Screening for odorant receptor genes expressed in Aedes aegypti involved in host-seeking, blood-feeding and oviposition behaviors. Parasit Vectors 2022, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Beaurepaire, A.; Rogers, C.W.; Lopez, D.; Evans, J.D.; Straub, L.; Neumann, P.; Cook, S.C.; Huang, Q. Gene expression and functional analyses of odorant receptors in small hive beetles (Aethina tumida). Int. J. Mol. Sci. 2020, 21, 4582. [Google Scholar] [CrossRef]
- Xu, L.; Tang, K.Y.; Chen, X.F.; Tao, Y.; Jiang, H.B.; Wang, J.J. Comparative transcriptomic analysis reveals female-biased olfactory genes potentially involved in plant volatile-mediated oviposition behavior of Bactrocera dorsalis. BMC Genom. 2021, 22, 25. [Google Scholar] [CrossRef]
- Sun, Y.L.; Dong, J.F.; Ning, C.; Ding, P.P.; Huang, L.Q.; Sun, J.G.; Wang, C.Z. An odorant receptor mediates the attractiveness of cis-jasmone to Campoletis chlorideae, the endoparasitoid of Helicoverpa armigera. Insect Mol. Biol. 2019, 28, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Q.; Guo, J.; Li, J.; Wang, J.; Wen, M.; Zhao, H.; Ren, B. Molecular basis of peripheral olfactory sensing during oviposition in the behavior of the parasitic wasp Anastatus japonicus. Insect Biochem. Mol. Biol. 2017, 89, 58–70. [Google Scholar] [CrossRef]
- Wang, C.; Li, G.; Miao, C.; Zhao, M.; Wang, B.; Guo, X. Nonanal modulates oviposition preference in female Helicoverpa assulta (Lepidoptera: Noctuidae) via the activation of peripheral neurons. Pest Manag. Sci. 2020, 76, 3159–3167. [Google Scholar] [CrossRef]
- Wu, H.; Li, R.T.; Dong, J.F.; Jiang, N.J.; Huang, L.Q.; Wang, C.Z. An odorant receptor and glomerulus responding to farnesene in Helicoverpa assulta (Lepidoptera: Noctuidae). Insect Biochem. Mol. Biol. 2019, 115, 103106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, Y.; Guo, M.; Sun, D.; Zhang, M.; Chu, X.; Berg, B.G.; Wang, G. A female-specific odorant receptor mediates oviposition deterrence in the moth Helicoverpa armigera. Curr. Biol. 2024, 34, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Zhang, J.; Yan, Q.; Miao, C.L.; Han, W.K.; Hou, W.; Yang, K.; Hansson, B.S.; Peng, Y.C.; Guo, J.M.; et al. The molecular basis of host selection in a Crucifer-specialized moth. Curr. Biol. 2020, 30, 4476–4482. [Google Scholar] [CrossRef]
- Zhang, J.; Komail Raza, S.A.; Wei, Z.; Keesey, I.W.; Parker, A.L.; Feistel, F.; Chen, J.; Cassau, S.; Fandino, R.A.; Grosse-Wilde, E.; et al. Competing beetles attract egg laying in a hawkmoth. Curr. Biol. 2022, 32, 861–869. [Google Scholar] [CrossRef]
- Zhang, G.F.; Xian, X.Q.; Zhang, Y.B.; Liu, W.X.; Liu, H.; Feng, X.D.; Ma, D.Y.; Wang, Y.S.; Gao, Y.H.; Zhang, R.; et al. Outbreak of the South American tomato leafminer, Tuta absoluta, in the Chinese mainland: Geographic and potential host range expansion. Pest Manag. Sci. 2021, 77, 5475–5488. [Google Scholar] [CrossRef] [PubMed]
- Tarusikirwa, V.L.; Machekano, H.; Mutamiswa, R.; Chidawanyika, F.; Nyamukondiwa, C. Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) on the “offensive” in Africa: Prospects for integrated management initiatives. Insects 2020, 11, 764. [Google Scholar] [CrossRef] [PubMed]
- Biondi, A.; Guedes RN, C.; Wan, F.H.; Desneux, N. Ecology, worldwide spread, and management of the invasive South American tomato pinworm, Tuta absoluta: Past, present, and future. Annu. Rev. Entomol. 2018, 63, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Guedes RN, C.; Roditakis, E.; Campos, M.R.; Haddi, K.; Bielza, P.; Siqueira, H.A.; Tsagkarakou, A.; Vontas, J.; Nauen, R. Insecticide resistance in the tomato pinworm Tuta absoluta: Patterns, spread, mechanisms, management and outlook. J. Pest Sci. 2019, 92, 1329–1342. [Google Scholar] [CrossRef]
- Gregg, P.C.; Del Socorro, A.P.; Landolt, P.J. Advances in attract-and-kill for agricultural pests: Beyond pheromones. Annu. Rev. Entomol. 2018, 63, 453–470. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, X.; Zhang, J.; He, T.; Huang, J.; Zhang, Z.; Wang, Y.; Hafeez, M.; Zhou, S.; Ren, X.; et al. Comprehensive metabolome and volatilome analyses in eggplant and tomato reveal their differential responses to Tuta absoluta infestation. Front. Plant Sci. 2021, 12, 757230. [Google Scholar] [CrossRef] [PubMed]
- Miano, R.N.; Ayelo, P.M.; Musau, R.; Hassanali, A.; Mohamed, S.A. Electroantennogram and machine learning reveal a volatile blend mediating avoidance behavior by Tuta absoluta females to a wild tomato plant. Sci. Rep. 2022, 12, 8965. [Google Scholar] [CrossRef] [PubMed]
- Anastasaki, E.; Drizou, F.; Milonas, P.G. Electrophysiological and oviposition responses of Tuta absoluta females to herbivore-induced volatiles in tomato plants. J. Chem. Ecol. 2018, 44, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Choo, Y.M.; Xu, P.; Hwang, J.K.; Zeng, F.; Tan, K.; Bhagavathy, G.; Chauhan, K.R.; Leal, W.S. Reverse chemical ecology approach for the identification of an oviposition attractant for Culex quinquefasciatus. Proc. Natl. Acad. Sci. USA 2018, 115, 714–719. [Google Scholar] [CrossRef]
- Xu, C.; Yang, F.; Duan, S.; Li, D.; Li, L.; Wang, M.; Zhou, A. Discovery of behaviorally active semiochemicals in Aenasius bambawalei using a reverse chemical ecology approach. Pest Manag. Sci. 2021, 77, 2843–2853. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhou, T.; Li, C.; Li, R.; Ye, X.; Tian, Z. Reverse chemical ecology guides the screening for Grapholita molesta pheromone synergists. Pest Manag. Sci. 2022, 78, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.-J.; Shi, J.-T.; Wang, C.-Z.; Guo, H.; Jiang, X.-J. Identification of an adult attractant for Anomala corpulenta by the reverse chemical ecology approach. J. Pest Sci. 2024, 1–11. [Google Scholar] [CrossRef]
- Franco, T.A.; Xu, P.; Brito, N.F.; Oliveira, D.S.; Wen, X.; Moreira, M.F.; Unelius, C.R.; Leal, W.S.; Melo AC, A. Reverse chemical ecology-based approach leading to the accidental discovery of repellents for Rhodnius prolixus, a vector of Chagas diseases refractory to DEET. Insect Biochem. Mol. Biol. 2018, 103, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Uda, Y.; Ono, Y.; Nakagawa, T.; Suwa, M.; Yamaoka, R.; Touhara, K. Highly selective tuning of a silkworm olfactory receptor to a key mulberry leaf volatile. Curr. Biol. 2009, 19, 881–890. [Google Scholar] [CrossRef] [PubMed]
- De Fouchier, A.; Walker, W.B., 3rd; Montagne, N.; Steiner, C.; Binyameen, M.; Schlyter, F.; Chertemps, T.; Maria, A.; Francois, M.C.; Monsempes, C.; et al. Functional evolution of Lepidoptera olfactory receptors revealed by deorphanization of a moth repertoire. Nat. Commun. 2017, 8, 15709. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Du, L.; Chen, Q.; Feng, Y.; Zhang, J.; Zhang, X.; Tian, K.; Cao, S.; Huang, T.; Jacquin-Joly, E.; et al. Odorant receptors for detecting flowering plant cues are functionally conserved across moths and butterflies. Mol. Biol. Evol. 2021, 38, 1413–1427. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, X.; Yin, Y.; Li, X.; He, K.; Zhao, X.; Li, X.; Luo, X.; Mei, Y.; Wang, Z.; et al. A chromosome-level genome assembly of tomato pinworm, Tuta absoluta. Sci. Data 2023, 10, 390. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Legan, A.W.; Jernigan, C.M.; Miller, S.E.; Fuchs, M.F.; Sheehan, M.J. Expansion and accelerated evolution of 9-exon odorant receptors in Polistes paper wasps. Mol. Biol. Evol. 2021, 38, 3832–3846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yang, R.R.; Jiang, X.C.; Xu, X.X.; Wang, B.; Wang, G.R. Genome-wide analysis of the odorant receptor gene family in Solenopsis invicta, Ooceraea biroi, and Monomorium pharaonis (Hymenoptera: Formicidae). Int. J. Mol. Sci. 2023, 24, 6624. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.D.; Chuang, C.H.; Kao, C.H.; Chen, S.H.; Wang, S.C.; Hsieh, P.H.; Chen, G.Y.; Mao, C.C.; Li, J.Y.; Jade Lu, M.Y.; et al. Decoding the genome of bloodsucking midge Forcipomyia taiwana (Diptera: Ceratopogonidae): Insights into odorant receptor expansion. Insect Biochem. Mol. Biol. 2024, 168, 104115. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Yu, Z.; He, X.; Hao, Y.; Qiao, L.; Luo, S.; Zhang, J.; Chen, B. Genome-wide identification and expression profiling of odorant receptor genes in the malaria vector Anopheles sinensis. Parasit Vectors 2022, 15, 143. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Crabbe MJ, C.; Ren, Z. Genome-wide identification and characterization of the chemosensory relative protein genes in Rhus gall aphid Schlechtendalia chinensis. BMC Genom. 2023, 24, 222. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Yin, C.; Tang, R.; Chen, M.; Wu, Q.; Huang, C.; Qian, W.; Rota-Stabelli, O.; Yang, N.; Wang, S.; et al. A chromosome-level genome assembly of Cydia pomonella provides insights into chemical ecology and insecticide resistance. Nat. Commun. 2019, 10, 4237. [Google Scholar] [CrossRef] [PubMed]
- Li, R.T.; Huang, L.Q.; Dong, J.F.; Wang, C.Z. A moth odorant receptor highly expressed in the ovipositor is involved in detecting host-plant volatiles. Elife 2020, 9, e53706. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.; Ou, X.; Wang, Y.; Zhou, Y.; Zhang, G.; Liu, W.; Wan, F.; Jiang, H.; Zhang, Y. Genome-Wide Identification, Evolution, and Female-Biased Expression Analysis of Odorant Receptors in Tuta absoluta (Lepidoptera: Gelechiidae). Life 2024, 14, 872. https://doi.org/10.3390/life14070872
Huang C, Ou X, Wang Y, Zhou Y, Zhang G, Liu W, Wan F, Jiang H, Zhang Y. Genome-Wide Identification, Evolution, and Female-Biased Expression Analysis of Odorant Receptors in Tuta absoluta (Lepidoptera: Gelechiidae). Life. 2024; 14(7):872. https://doi.org/10.3390/life14070872
Chicago/Turabian StyleHuang, Cong, Xiaolan Ou, Yusheng Wang, Yanan Zhou, Guifen Zhang, Wanxue Liu, Fanghao Wan, Hongbo Jiang, and Yibo Zhang. 2024. "Genome-Wide Identification, Evolution, and Female-Biased Expression Analysis of Odorant Receptors in Tuta absoluta (Lepidoptera: Gelechiidae)" Life 14, no. 7: 872. https://doi.org/10.3390/life14070872