

Xenobiology for the Biocontainment of Synthetic Organisms: Opportunities and Challenges

Abstract

{kind=link}
{kind=link}
{kind=link}
1. Background
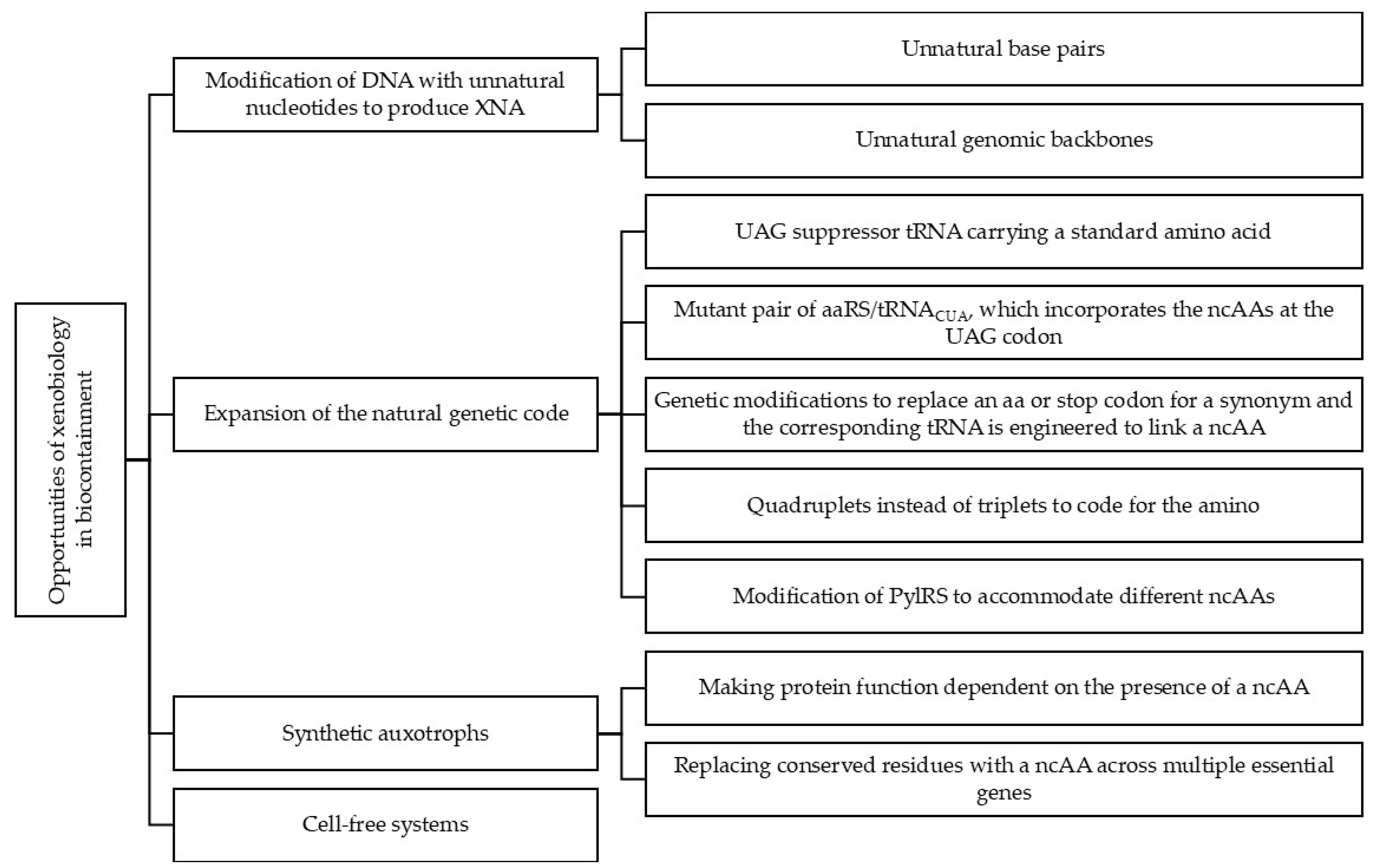
2. Opportunities of Xenobiology in Biocontainment
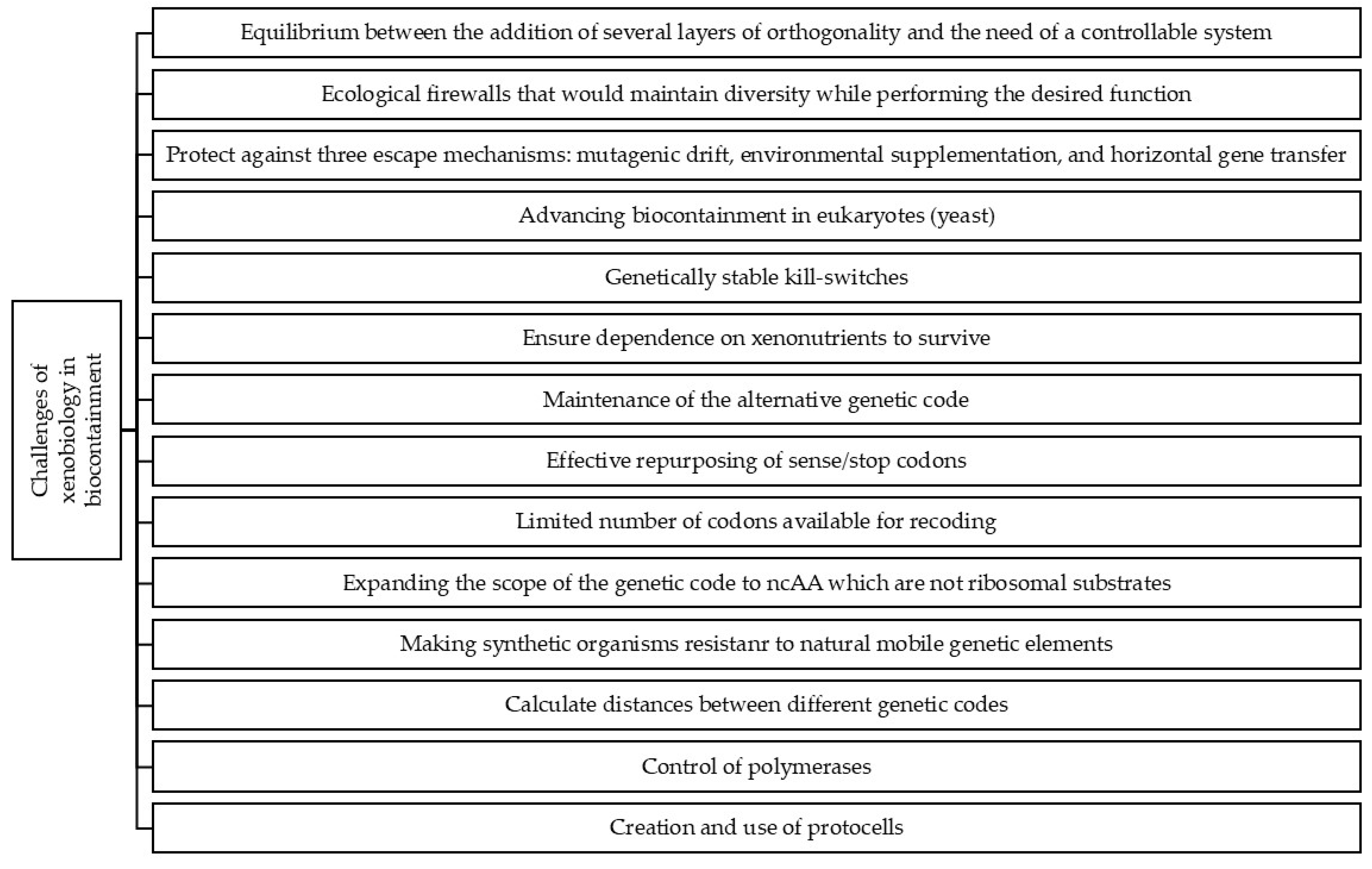
3. Challenges of Xenobiology in Biocontainment
4. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wooster, H. Xenobiology. Science 1961, 134, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Pei, L. Synthetic Toxicology: Where Engineering Meets Biology and Toxicology. Toxicol. Sci. 2011, 120 (Suppl. S1), S204–S224. [Google Scholar] [CrossRef] [PubMed]
- Deplazes, A. Piecing together a puzzle. An exposition of synthetic biology. EMBO Rep. 2009, 10, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Synthetic Biology II—Risk Assessment Methodologies and Safety Aspects. Available online: https://health.ec.europa.eu/publications/synthetic-biology-ii-risk-assessment-methodologies-and-safety-aspects_en (accessed on 23 February 2024).
- Schmidt, M.; Pei, L.; Budisa, N. Xenobiology: State-of-the-art, ethics, and philosophy of new-to-nature organisms. Adv. Biochem. Eng. Biotechnol. 2018, 162, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Kubyshkin, V.; Budisa, N. The Alanine World Model for the Development of the Amino Acid Repertoire in Protein Biosynthesis. Int. J. Mol. Sci. 2019, 20, 5507. [Google Scholar] [CrossRef] [PubMed]
- Kubyshkin, V.; Budisa, N. Anticipating alien cells with alternative genetic codes: Away from the alanine world! Curr. Opin. Biotechnol. 2019, 60, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Berg, P.; Baltimore, D.; Brenner, S.; Roblin, R.O.; Singer, M.F. Summary Statement of the Asilomar Conference on Recombinant DNA Molecules. Proc. Natl. Acad. Sci. USA 1975, 72, 1981–1984. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Rocha, C.G. The Synthetic Nature of Biology. In Ambivalences of Creating Life: Societal and Philosophical Dimensions of Synthetic Biology; Springer: Berlin/Heidelberg, Germany, 2015; Volume 45, pp. 9–53. [Google Scholar] [CrossRef]
- Rovner, A.J.; Haimovich, A.D.; Katz, S.R.; Li, Z.; Grome, M.W.; Gassaway, B.M.; Amiram, M.; Patel, J.R.; Gallagher, R.R.; Rinehart, J.; et al. Recoded Organisms Engineered to Depend on Synthetic Amino Acids. Nature 2015, 518, 89–93. [Google Scholar] [CrossRef]
- Chaput, J.C.; Herdewijn, P.; Hollenstein, M. Orthogonal Genetic Systems. Chembiochem 2020, 21, 1408–1411. [Google Scholar] [CrossRef]
- Kubyshkin, V.; Budisa, N. Synthetic Alienation of Microbial Organisms by Using Genetic Code Engineering: Why and How? Biotechnol. J. 2017, 12, 1600097. [Google Scholar] [CrossRef]
- Schmidt, M. A Metric Space for Semantic Containment: Towards the Implementation of Genetic Firewalls. Biosystems 2019, 185, 104015. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M. Xenobiology: A New Form of Life as the Ultimate Biosafety Tool. Bioessays 2010, 32, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Torres, L.; Krüger, A.; Csibra, E.; Gianni, E.; Pinheiro, V.B. Synthetic biology approaches to biological containment: Pre-emptively tackling potential risks. Essays. Biochem. 2016, 60, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.J. NIH Guidelines for Research Involving Recombinant DNA Molecules. Account. Res. 1993, 3, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Diwo, C.; Budisa, N. Alternative Biochemistries for Alien Life: Basic Concepts and Requirements for the Design of a Robust Biocontainment System in Genetic Isolation. Genes 2018, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, S.A.; Diggans, J.; Densmore, D.; Dai, J.; Knight, T.; Leproust, E.; Boeke, J.D.; Wheeler, N.; Cai, Y. Safety by design: Biosafety and biosecurity in the age of synthetic genomics. iScience 2023, 26, 106165. [Google Scholar] [CrossRef] [PubMed]
- Vladilo, G.; Hassanali, A. Hydrogen Bonds and Life in the Universe. Life 2018, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Beiranvand, N.; Freindorf, M.; Kraka, E. Hydrogen Bonding in Natural and Unnatural Base Pairs-A Local Vibrational Mode Study. Molecules 2021, 26, 2268. [Google Scholar] [CrossRef] [PubMed]
- Herdewijn, P.; Marlière, P. Toward safe genetically modified organisms through the chemical diversification of nucleic acids. Chem. Biodivers. 2009, 6, 791–808. [Google Scholar] [CrossRef]
- Benner, S.A.; Karalkar, N.B.; Hoshika, S.; Laos, R.; Shaw, R.W.; Matsuura, M.; Fajardo, D.; Moussatche, P. Alternative Watson-Crick Synthetic Genetic Systems. Cold Spring Harb. Perspect. Biol. 2016, 8, a023770. [Google Scholar] [CrossRef]
- Kim, H.J.; Leal, N.A.; Hoshika, S.; Benner, S.A. Ribonucleosides for an artificially expanded genetic information system. J. Org. Chem. 2014, 79, 3194–3199. [Google Scholar] [CrossRef] [PubMed]
- Sefah, K.; Yang, Z.; Bradley, K.M.; Hoshika, S.; Jiménez, E.; Zhang, L.; Zhu, G.; Shanker, S.; Yu, F.; Turek, D.; et al. In Vitro selection with artificial expanded genetic information systems. Proc. Natl. Acad. Sci. USA 2014, 111, 1449–1454. [Google Scholar] [CrossRef]
- Zhang, Y.; Ptacin, J.L.; Fischer, E.C.; Aerni, H.R.; Caffaro, C.E.; San Jose, K.; Feldman, A.W.; Turner, C.R.; Romesberg, F.E. A semi-synthetic organism that stores and retrieves increased genetic information. Nature 2017, 551, 644–647. [Google Scholar] [CrossRef]
- Hoshika, S.; Leal, N.A.; Kim, M.J.; Kim, M.S.; Karalkar, N.B.; Kim, H.J.; Bates, A.M.; Watkins, N.E., Jr.; SantaLucia, H.A.; Meyer, A.J.; et al. Hachimoji DNA and RNA: A genetic system with eight building blocks. Science 2019, 363, 884–887. [Google Scholar] [CrossRef]
- Futami, K.; Kimoto, M.; Lim, Y.W.S.; Hirao, I. Genetic Alphabet Expansion Provides Versatile Specificities and Activities of Unnatural-Base DNA Aptamers Targeting Cancer Cells. Mol. Ther. Nucleic Acids 2019, 14, 158–170. [Google Scholar] [CrossRef]
- Malyshev, D.A.; Dhami, K.; Lavergne, T.; Chen, T.; Dai, N.; Foster, J.M.; Corrêa, I.R., Jr.; Romesberg, F.E. A semi-synthetic organism with an expanded genetic alphabet. Nature 2014, 509, 385–388. [Google Scholar] [CrossRef]
- Dien, V.T.; Morris, S.E.; Karadeema, R.J.; Romesberg, F.E. Expansion of the genetic code via expansion of the genetic alphabet. Curr. Opin. Chem. Biol. 2018, 46, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Dien, V.T.; Holcomb, M.; Feldman, A.W.; Fischer, E.C.; Dwyer, T.J.; Romesberg, F.E. Progress Toward a Semi-Synthetic Organism with an Unrestricted Expanded Genetic Alphabet. J. Am. Chem. Soc. 2018, 140, 16115–16123. [Google Scholar] [CrossRef]
- Budisa, N. Expanded genetic code for the engineering of ribosomally synthetized and post-translationally modified peptide natural products (RiPPs). Curr. Opin. Biotechnol. 2013, 24, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y. Translational Control Using an Expanded Genetic Code. Int. J. Mol. Sci. 2019, 20, 887. [Google Scholar] [CrossRef]
- Fredens, J.; Wang, K.; de la Torre, D.; Funke, L.F.H.; Robertson, W.E.; Christova, Y.; Chia, T.; Schmied, W.H.; Dunkelmann, D.L.; Beránek, V.; et al. Total Synthesis of Escherichia Coli with a Recoded Genome. Nature 2019, 569, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Lajoie, M.J.; Xiao, H.; Church, G.M.; Schultz, P.G. A bacterial strain with a unique quadruplet codon specifying non-native amino acids. ChemBioChem 2014, 15, 1782–1786. [Google Scholar] [CrossRef]
- Fan, C.; Xiong, H.; Reynolds, N.M.; Söll, D. Rationally evolving tRNAPyl for efficient incorporation of noncanonical amino acids. Nucleic. Acids. Res. 2015, 43, e156. [Google Scholar] [CrossRef] [PubMed]
- Crnković, A.; Suzuki, T.; Söll, D.; Reynolds, N.M. Pyrrolysyl-tRNA Synthetase, an Aminoacyl-tRNA Synthetase for Genetic Code Expansion. Croat. Chem. Acta 2016, 89, 163–174. [Google Scholar] [CrossRef]
- Koch, N.G.; Baumann, T.; Budisa, N. Efficient Unnatural Protein Production by Pyrrolysyl-tRNA Synthetase With Genetically Fused Solubility Tags. Front. Bioeng. Biotechnol. 2021, 9, 807438. [Google Scholar] [CrossRef]
- Mayer, C. Selection, Addiction and Catalysis: Emerging Trends for the Incorporation of Noncanonical Amino Acids into Peptides and Proteins In Vivo. ChemBioChem 2019, 20, 1357–1364. [Google Scholar] [CrossRef]
- Lee, J.W.; Chan, C.T.Y.; Slomovic, S.; Collins, J.J. Next-Generation Biocontainment Systems for Engineered Organisms. Nat. Chem. Biol. 2018, 14, 530–537. [Google Scholar] [CrossRef]
- Chang, T.; Ding, W.; Yan, S.; Wang, Y.; Zhang, H.; Zhang, Y.; Ping, Z.; Zhang, H.; Huang, Y.; Zhang, J.; et al. A Robust Yeast Biocontainment System with Two-Layered Regulation Switch Dependent on Unnatural Amino Acid. Nat. Commun. 2023, 14, 6487. [Google Scholar] [CrossRef] [PubMed]
- Orelle, C.; Carlson, E.D.; Szal, T.; Florin, T.; Jewett, M.C.; Mankin, A.S. Protein Synthesis by Ribosomes with Tethered Subunits. Nature 2015, 524, 119–124. [Google Scholar] [CrossRef]
- Jia, B.; Qi, H.; Li, B.-Z.; Pan, S.; Liu, D.; Liu, H.; Cai, Y.; Yuan, Y.-J. Orthogonal Ribosome Biofirewall. ACS Synth. Biol. 2017, 6, 2108–2117. [Google Scholar] [CrossRef]
- Kato, Y. An Engineered Bacterium Auxotrophic for an Unnatural Amino Acid: A Novel Biological Containment System. PeerJ 2015, 3, e1247. [Google Scholar] [CrossRef] [PubMed]
- Mandell, D.J.; Lajoie, M.J.; Mee, M.T.; Takeuchi, R.; Kuznetsov, G.; Norville, J.E.; Gregg, C.J.; Stoddard, B.L.; Church, G.M. Biocontainment of Genetically Modified Organisms by Synthetic Protein Design. Nature 2015, 518, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Tack, D.S.; Ellefson, J.W.; Thyer, R.; Wang, B.; Gollihar, J.; Forster, M.T.; Ellington, A.D. Addicting Diverse Bacteria to a Noncanonical Amino Acid. Nat. Chem. Biol. 2016, 12, 138–140. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Schultz, P.G. A Strategy for Creating Organisms Dependent on Noncanonical Amino Acids. Angew. Chem. Weinh. Bergstr. Ger. Weinh. 2017, 129, 9298–9301. [Google Scholar] [CrossRef]
- Koh, M.; Yao, A.; Gleason, P.R.; Mills, J.H.; Schultz, P.G. A General Strategy for Engineering Noncanonical Amino Acid Dependent Bacterial Growth. J. Am. Chem. Soc. 2019, 141, 16213–16216. [Google Scholar] [CrossRef] [PubMed]
- Saccharomyces Cerevisiae 2.0. Available online: https://syntheticyeast.github.io/ (accessed on 23 February 2024).
- Kato, Y. Tight Translational Control Using Site-Specific Unnatural Amino Acid Incorporation with Positive Feedback Gene Circuits. ACS Synth. Biol. 2018, 7, 1956–1963. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yuan, Z.; Niu, W.; Li, Q.; Guo, J. Synthetic Biology Approach for the Development of Conditionally Replicating HIV-1 Vaccine. J. Chem. Technol. Biotechnol. 2017, 92, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N.; Lai, Y.; Fuerte-Stone, J.; Mimee, M.; Lu, T.K. Cas9-assisted biological containment of a genetically engineered human commensal bacterium and genetic elements. Nat. Commun. 2024, 15, 2096. [Google Scholar] [CrossRef] [PubMed]
- Fujino, T.; Tozaki, M.; Murakami, H. An Amino Acid-Swapped Genetic Code. ACS Synth. Biol. 2020, 9, 2703–2713. [Google Scholar] [CrossRef]
- Whitford, C.M.; Dymek, S.; Kerkhoff, D.; März, C.; Schmidt, O.; Edich, M.; Droste, J.; Pucker, B.; Rückert, C.; Kalinowski, J. Auxotrophy to Xeno-DNA: An Exploration of Combinatorial Mechanisms for a High-Fidelity Biosafety System for Synthetic Biology Applications. J. Biol. Eng. 2018, 12, 13. [Google Scholar] [CrossRef]
- Chemla, Y.; Ozer, E.; Schlesinger, O.; Noireaux, V.; Alfonta, L. Genetically Expanded Cell-Free Protein Synthesis Using Endogenous Pyrrolysyl Orthogonal Translation System: Genetically Expanded Cell-Free Protein Synthesis. Biotechnol. Bioeng. 2015, 112, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Des Soye, B.J.; Patel, J.R.; Isaacs, F.J.; Jewett, M.C. Repurposing the Translation Apparatus for Synthetic Biology. Curr. Opin. Chem. Biol. 2015, 28, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Ntai, I.; Haimovich, A.D.; Kelleher, N.L.; Isaacs, F.J.; Jewett, M.C. Cell-Free Protein Synthesis from a Release Factor 1 Deficient Escherichia Coli Activates Efficient and Multiple Site-Specific Nonstandard Amino Acid Incorporation. ACS Synth. Biol. 2014, 3, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Khambhati, K.; Bhattacharjee, G.; Gohil, N.; Braddick, D.; Kulkarni, V.; Singh, V. Exploring the Potential of Cell-Free Protein Synthesis for Extending the Abilities of Biological Systems. Front. Bioeng. Biotechnol. 2019, 7, 248. [Google Scholar] [CrossRef] [PubMed]
- Copeland, C.E.; Langlois, A.; Kim, J.; Kwon, Y.-C. The Cell-Free System: A New Apparatus for Affordable, Sensitive, and Portable Healthcare. Biochem. Eng. J. 2021, 175, 108124. [Google Scholar] [CrossRef]
- Karig, D.K. Cell-free synthetic biology for environmental sensing and remediation. Curr. Opin. Biotechnol. 2017, 45, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Lee, J.A.; Biondo, J.R.; Lux, J.E.; Raig, R.M.; Berger, P.N.; Bernhards, C.B.; Kuhn, D.L.; Gupta, M.K.; Lux, M.W. Cell-Free Protein Expression in Polymer Materials. ACS Synth. Biol. 2024, 13, 1152–1164. [Google Scholar] [CrossRef]
- Lee, M.S.; Raig, R.M.; Gupta, M.K.; Lux, M.W. Lyophilized Cell-Free Systems Display Tolerance to Organic Solvent Exposure. ACS Synth. Biol. 2020, 9, 1951–1957. [Google Scholar] [CrossRef]
- Wright, O.; Stan, G.-B.; Ellis, T. Building-in Biosafety for Synthetic Biology. Microbiology 2013, 159, 1221–1235. [Google Scholar] [CrossRef]
- Vidiella, B.; Solé, R. Ecological firewalls for synthetic biology. iScience 2022, 25, 104658. [Google Scholar] [CrossRef]
- Pavão, G.; Sfalcin, I.; Bonatto, D. Biocontainment Techniques and Applications for Yeast Biotechnology. Fermentation 2023, 9, 341. [Google Scholar] [CrossRef]
- Richardson, S.M.; Mitchell, L.A.; Stracquadanio, G.; Yang, K.; Dymond, J.S.; DiCarlo, J.E.; Lee, D.; Huang, C.L.; Chandrasegaran, S.; Cai, Y.; et al. Design of a synthetic yeast genome. Science 2017, 355, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Chari, R.; Church, G.M. Beyond editing to writing large genomes. Nat. Rev. Genet. 2017, 18, 749–760. [Google Scholar] [CrossRef]
- Kato, Y.; Mori, H. Genetically stable kill-switch using “demon and angel” expression construct of essential genes. Front. Bioeng. Biotechnol. 2024, 12, 1365870. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Kubyshkin, V. How to Quantify a Genetic Firewall? A Polarity-Based Metric for Genetic Code Engineering. ChemBioChem 2021, 22, 1268–1284. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.M.; Nakano, Y. Breaking the deadlock in genetic code expansion. Nat. Chem. Biol. 2024, 20, 406–407. [Google Scholar] [CrossRef] [PubMed]
- Hartman, H.; Smith, T.F. The Evolution of the Ribosome and the Genetic Code. Life 2014, 4, 227–249. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.F.; Hartman, H. The Evolution of Class II Aminoacyl-tRNA Synthetases and the First Code. FEBS Lett. 2015, 589, 3499–3507. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.M.; Reynolds, N.M.; Rivera, K.; Connolly, M.; Guo, L.-T.; Ling, J.; Pappin, D.J.; Church, G.M.; Söll, D. Efficient Reassignment of a Frequent Serine Codon in Wild-Type Escherichia Coli. ACS Synth. Biol. 2016, 5, 163–171. [Google Scholar] [CrossRef]
- Dunkelmann, D.L.; Piedrafita, C.; Dickson, A.; Liu, K.C.; Elliott, T.S.; Fiedler, M.; Bellini, D.; Zhou, A.; Cervettini, D.; Chin, J.W. Adding α, α-disubstituted and β-linked monomers to the genetic code of an organism. Nature 2024, 625, 603–610. [Google Scholar] [CrossRef]
- Zürcher, J.F.; Robertson, W.E.; Kappes, T.; Petris, G.; Elliott, T.S.; Salmond, G.P.C.; Chin, J.W. Refactored Genetic Codes Enable Bidirectional Genetic Isolation. Science 2022, 378, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Nyerges, A.; Vinke, S.; Flynn, R.; Owen, S.V.; Rand, E.A.; Budnik, B.; Keen, E.; Narasimhan, K.; Marchand, J.A.; Baas-Thomas, M.; et al. A Swapped Genetic Code Prevents Viral Infections and Gene Transfer. Nature 2023, 615, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Ghadessy, F.J.; Holliger, P. Compartmentalized Self-Replication: A Novel Method for the Directed Evolution of Polymerases and Other Enzymes. Methods Mol. Biol. 2007, 352, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Sismour, A.M.; Sheng, P.; Puskar, N.L.; Benner, S.A. Enzymatic Incorporation of a Third Nucleobase Pair. Nucleic Acids Res. 2007, 35, 4238–4249. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Chen, L.; Nilsson, M.; Abe, S. Bridging Nonliving and Living Matter. Artif. Life 2003, 9, 269–316. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, W.; Wang, Z.; Zhao, H.; Shi, S. Development of Host-Orthogonal Genetic Systems for Synthetic Biology. Adv. Biol. 2021, 5, e2000252. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Tatay, L.; Hernández-Andreu, J.M. Xenobiology for the Biocontainment of Synthetic Organisms: Opportunities and Challenges. Life 2024, 14, 996. https://doi.org/10.3390/life14080996
Gómez-Tatay L, Hernández-Andreu JM. Xenobiology for the Biocontainment of Synthetic Organisms: Opportunities and Challenges. Life. 2024; 14(8):996. https://doi.org/10.3390/life14080996
Chicago/Turabian StyleGómez-Tatay, Lucía, and José Miguel Hernández-Andreu. 2024. "Xenobiology for the Biocontainment of Synthetic Organisms: Opportunities and Challenges" Life 14, no. 8: 996. https://doi.org/10.3390/life14080996
APA StyleGómez-Tatay, L., & Hernández-Andreu, J. M. (2024). Xenobiology for the Biocontainment of Synthetic Organisms: Opportunities and Challenges. Life, 14(8), 996. https://doi.org/10.3390/life14080996