
Influence of Radical Generation and Elimination on Sebum Production of Hamster Sebaceous Gland Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Matrerials
Samples and Reagents
2.2. Cell and Cell Culture
2.3. Sample Addition
2.4. Viability of Cells
2.5. Extraction of Lipids from Cells
2.6. Quantitative Analysis of Lipids
2.7. Measurement of Differentiated Cell Number
2.8. Determination of Scavenging Efficiency
2.8.1. Superoxide Anion Radical
2.8.2. Nitric Oxide
2.8.3. Hydrogen Peroxide (H2O2)
2.9. Statistical Analysis
3. Results
3.1. Effect of Antioxidants on Viability of Hamster Sebaceous Gland Cells
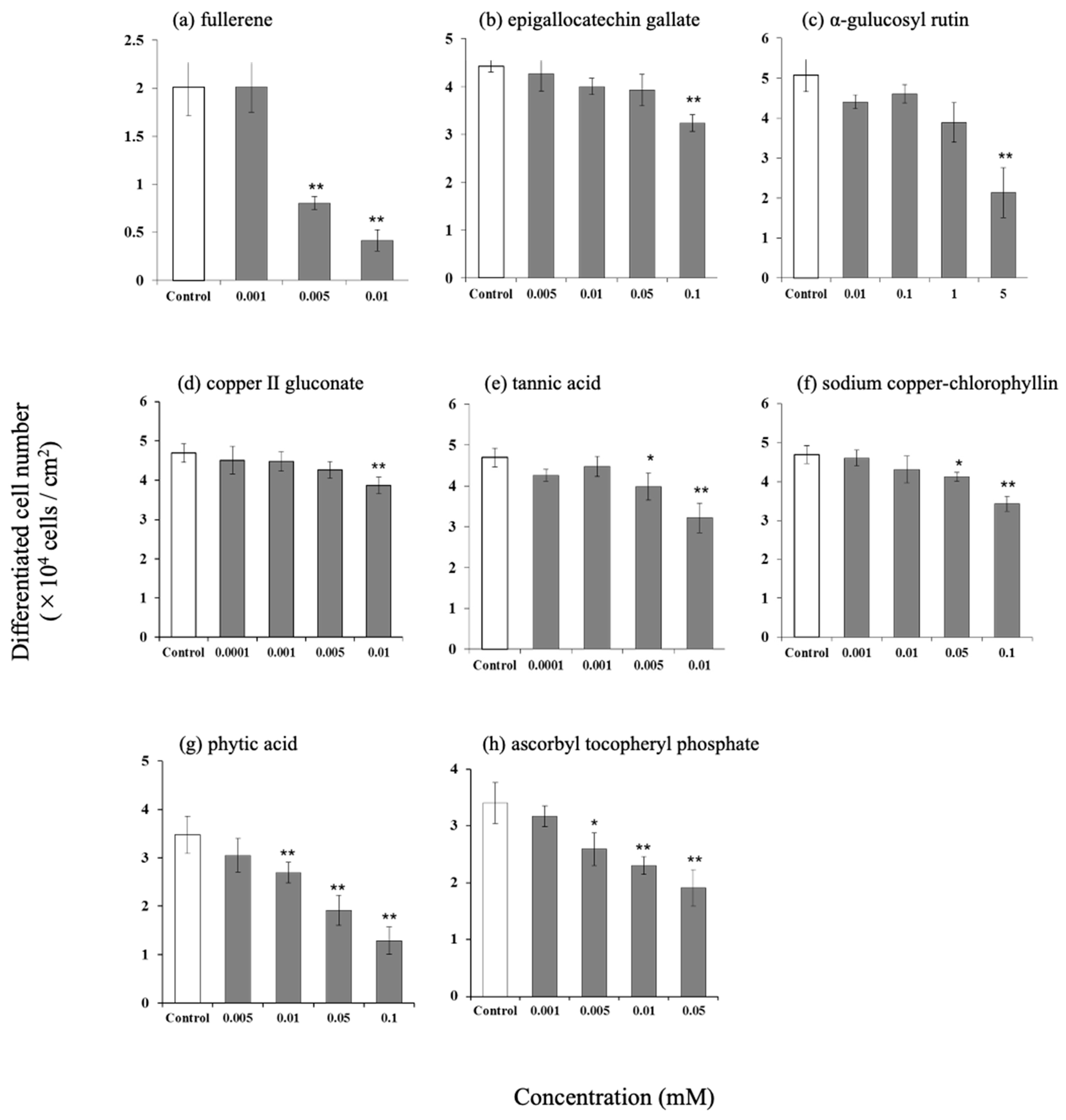
3.2. Effect of Antioxidants on the Number of Differentiated Cells in Hamster Sebaceous Gland Cells
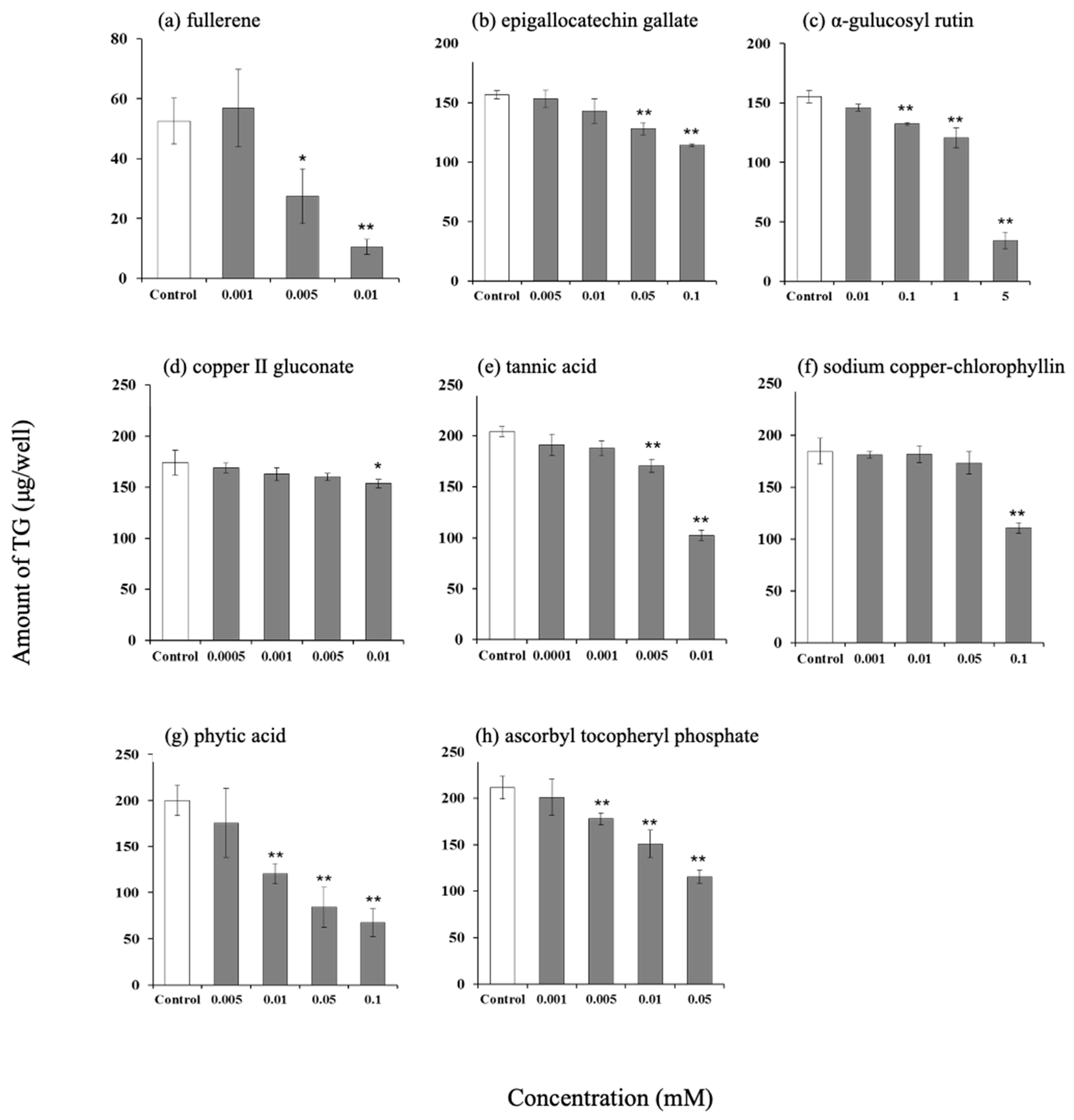
3.3. Effect of Antioxidants on Intracellular Triglyceride Content in Hamster Sebaceous Gland Cells
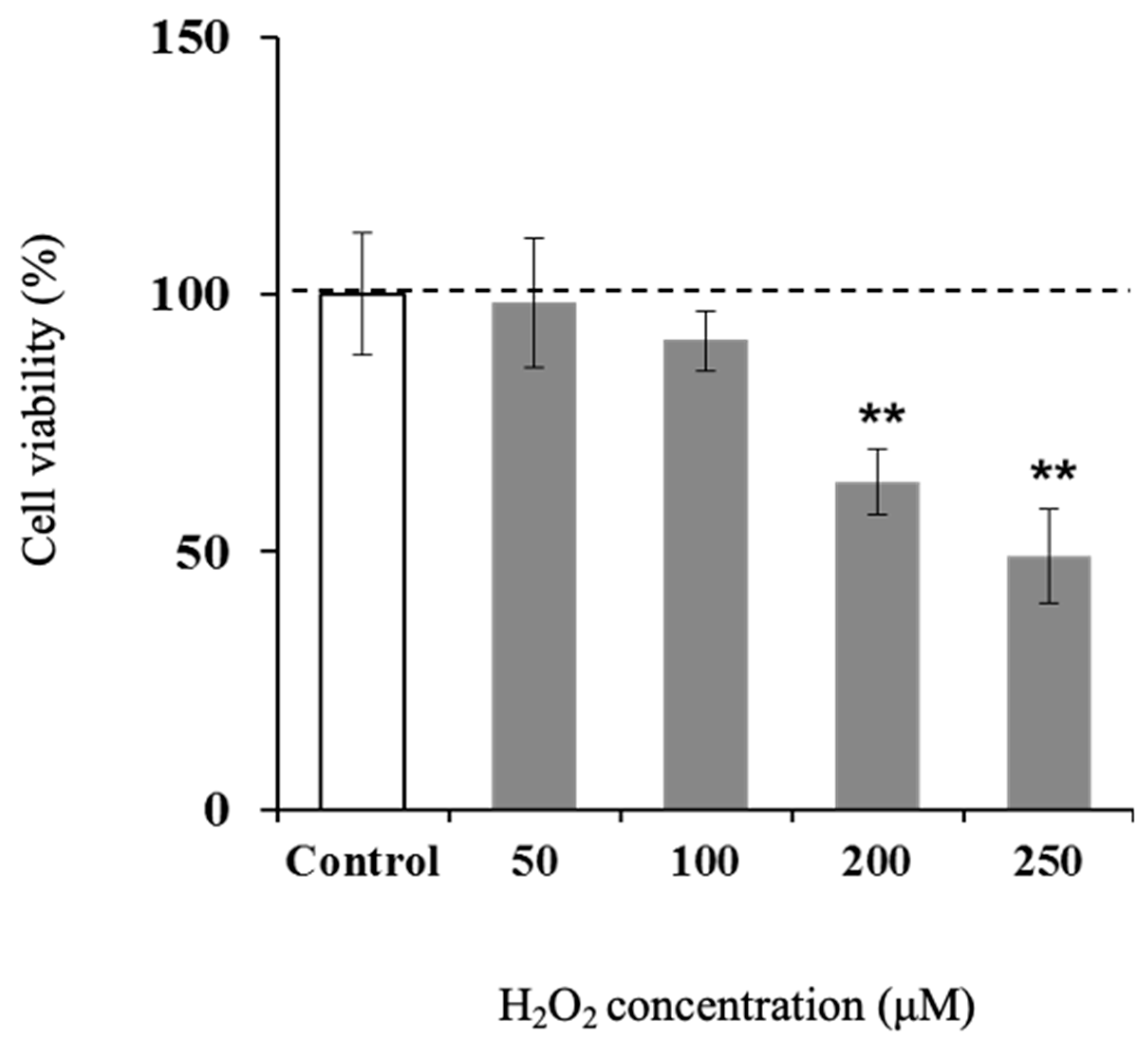
3.4. Effect of H2O2 on Viability of Hamster Sebaceous Gland Cells
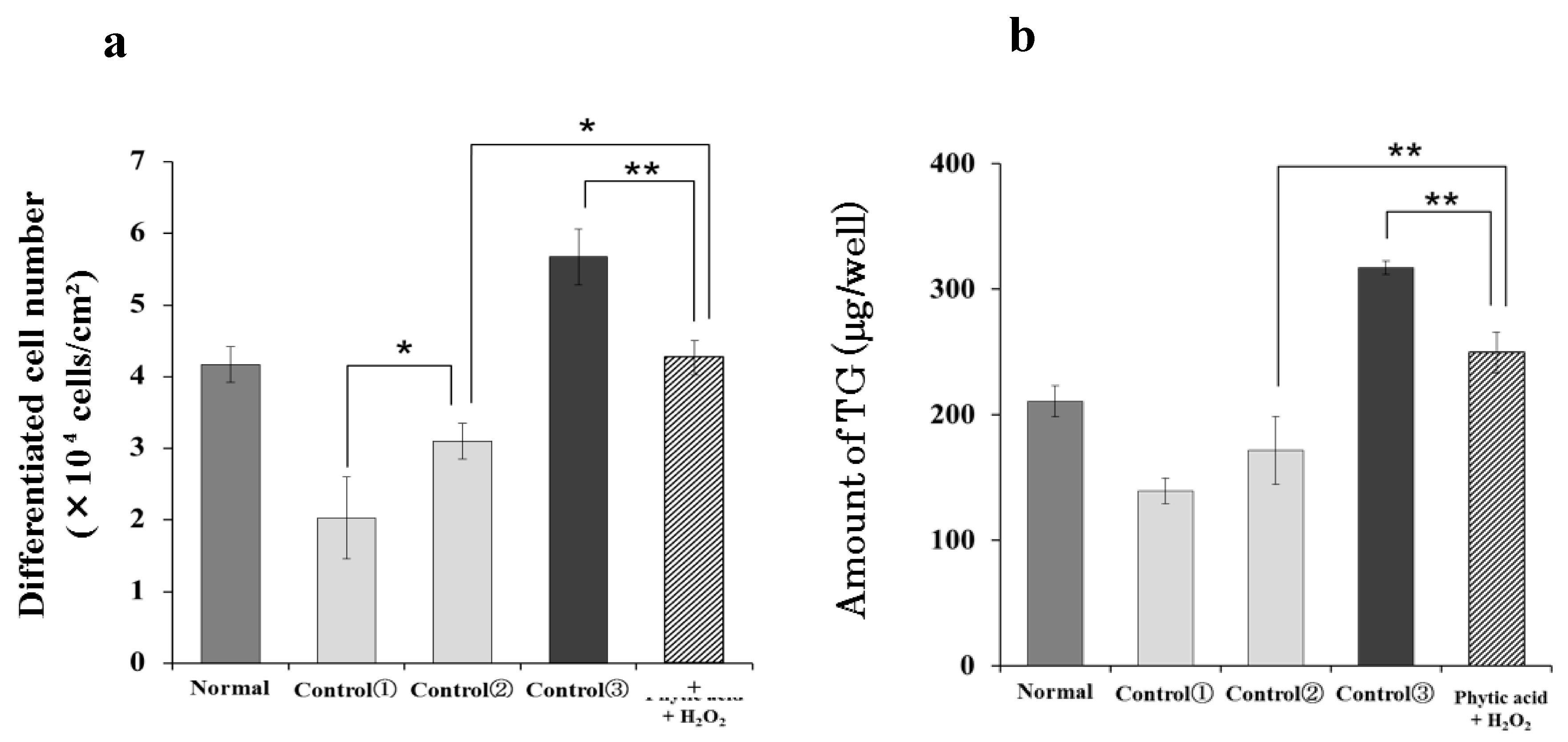
3.5. Effect of H2O2 on the Number of Differentiated Cells and Intracellular Triglyceride Content of Hamster Sebaceous Gland Cells
3.6. Effect of Radical Induction on the Number of Differentiated Cells and Intracellular Triglyceride Content of Hamster Sebaceous Gland Cells
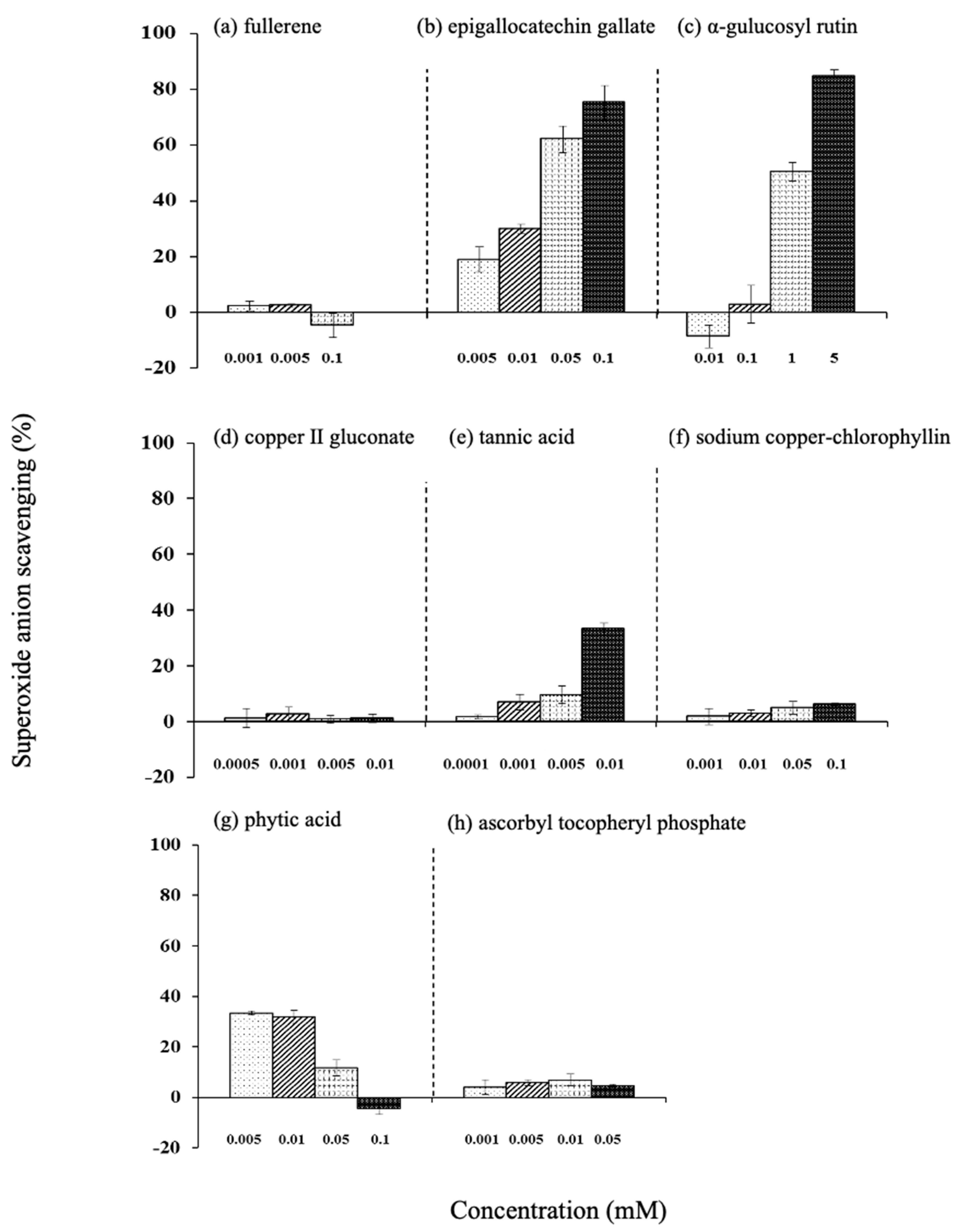
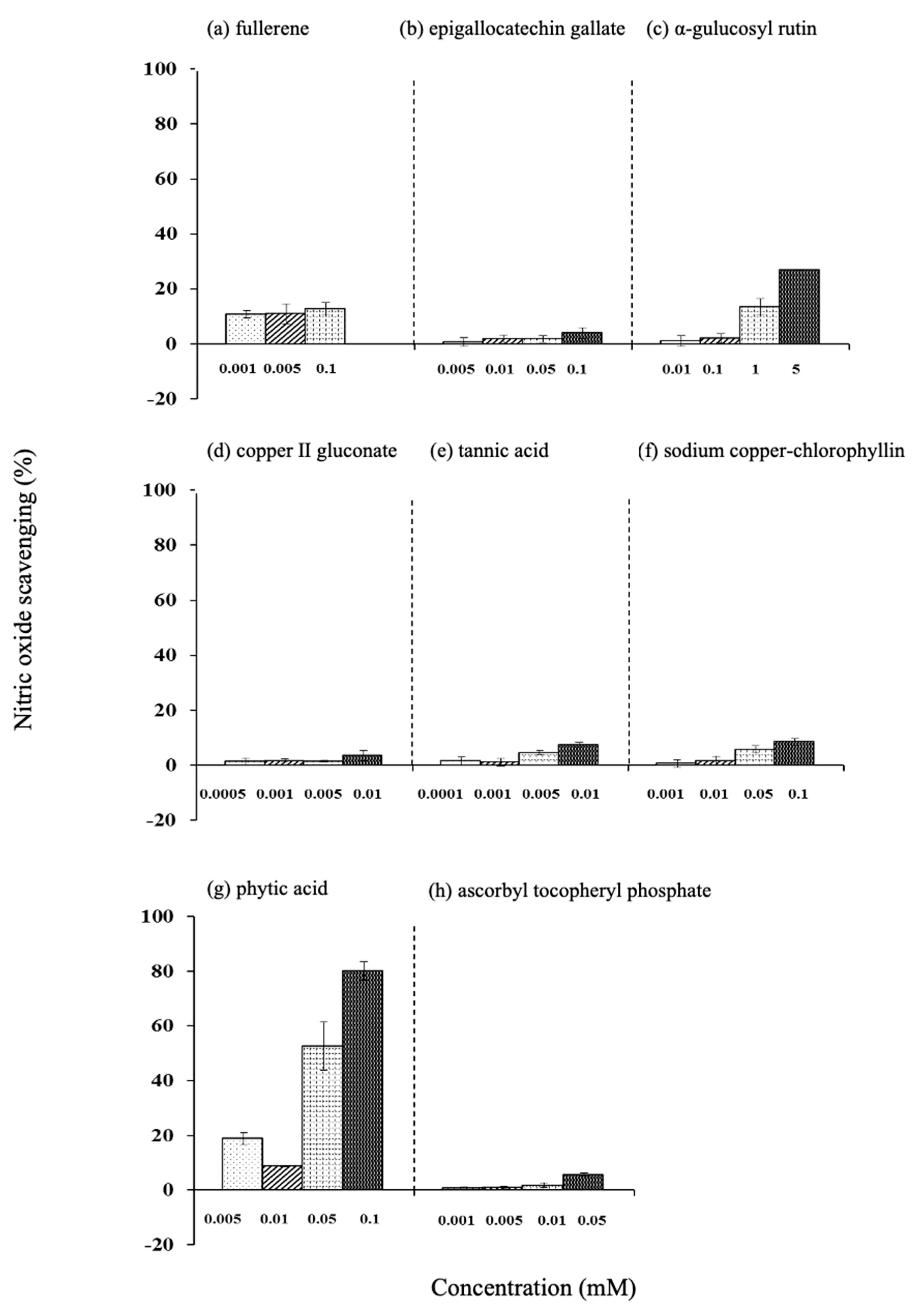
3.7. Efficiency of Various Antioxidants
3.7.1. Superoxide Anion Radical Scavenging
3.7.2. Nitric Oxide Scavenging Efficiency
3.7.3. H2O2 Scavenging Efficiency
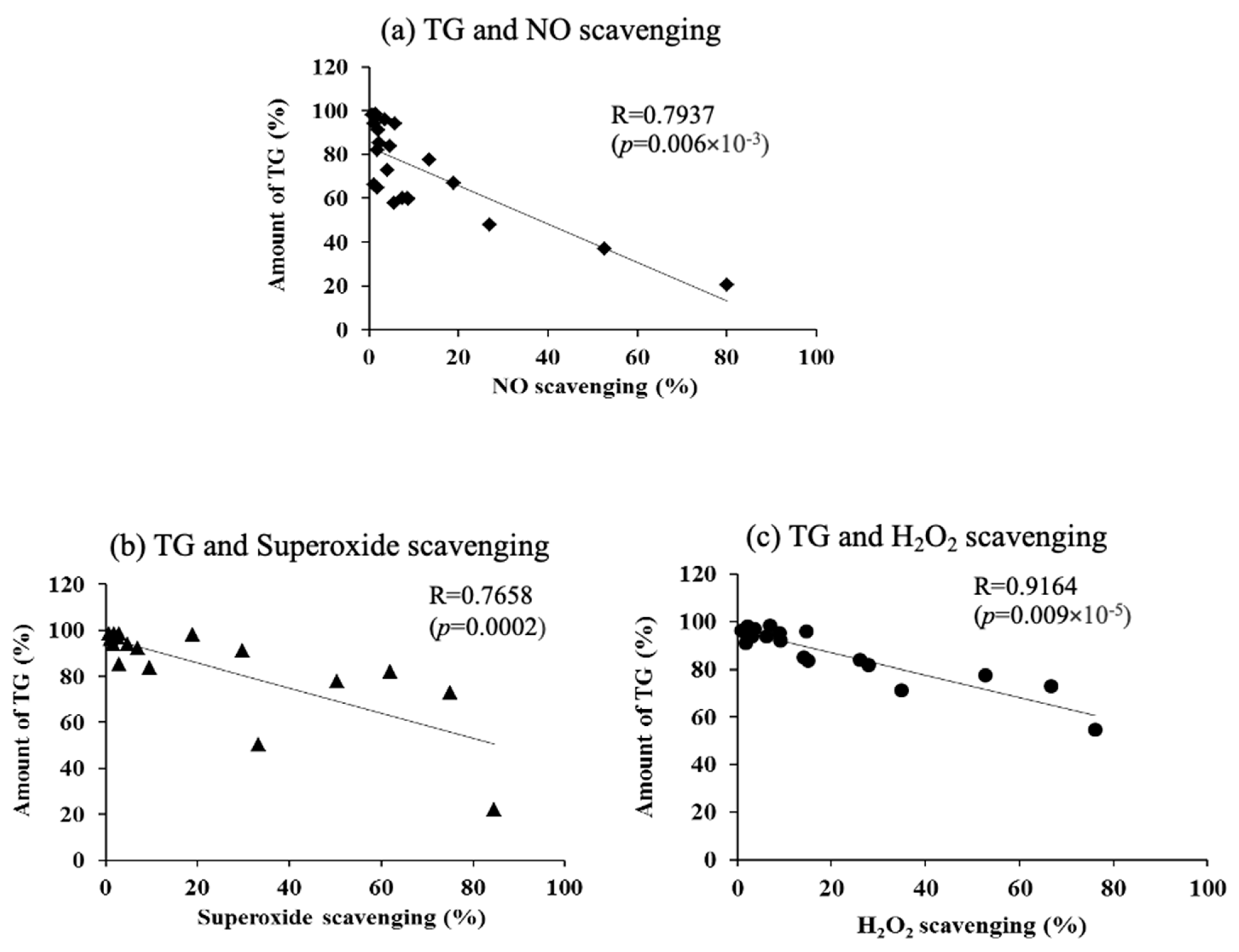
3.8. Correlation Between Sebum Production and Reactive Oxygen Species (ROS) Scavenging Efficiency in Hamster Sebaceous Gland Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Drake, D.R.; Brogden, K.A.; Dawson, D.V.; Wertz, P.W. Thematic review series: Skin lipids—Antimicrobial lipids at the skin surface. J. Lipid Res. 2008, 49, 4–11. [Google Scholar] [CrossRef]
- Shamloul, G.; Khachemoune, A. An updated review of the sebaceous gland and its role in health and diseases Part 1: Embryology, evolution, structure, and function of sebaceous glands. Dermatol. Ther. 2021, 34, e14695. [Google Scholar] [CrossRef] [PubMed]
- Thody, A.J.; Shuster, S. Control and function of sebaceous glands. Physiol. Rev. 1989, 69, 383–416. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, Y.; Akamatsu, H.; Okano, Y.; Masaki, H.; Horio, T. Effects of UV irradiation on the sebaceous gland and sebum secretion in hamsters. J. Dermatol. Sci. 2003, 31, 151–159. [Google Scholar]
- Makrantonaki, E.; Ganceviciene, R.; Zouboulis, C. An update on the role of the sebaceous gland in the pathogenesis of acne. Derm.-Endocrinol. 2011, 3, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Ebling, F.J.; Ebling, E.; McCaffery, V.; Skinner, J. The responses of the sebaceous glands of the hypophysectomized-castrated male rat to 5-androstanedione and 5-androstane-3,17–diol. J. Investig. Dermatol. 1973, 64, 183–187. [Google Scholar] [CrossRef]
- Szöllősi, A.G.; Oláh, A.; Bíró, T.; Tóth, B.I. Recent advances in the endocrinology of the sebaceous gland. Dermatoendocrinology 2017, 9, e1361576. [Google Scholar] [CrossRef] [PubMed]
- Thody, A.J.; Cooper, M.F.; Bowden, P.E.; Meddis, D.; Shuster, S. Effect of alpha-melanocyte-stimulating hormone and testosterone on cutaneous and modified sebaceous glands in the rat. J. Endocrinol. 1976, 71, 279–288. [Google Scholar] [CrossRef]
- Sansone, G.; Davidson, W.; Cummings, B.; Reisner, R.M. Sebaceous gland lipogenesis induced by testosterone: Early metabolic events. J. Investig. Dermatol. 1971, 57, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Seltmann, H.; Hiroi, N.; Chen, W.; Young, M.; Oeff, M.; Scherbaum, W.A.; Orfanos, C.E.; McCann, S.M.; Bornstein, S.R. Corticotropin-releasing hormone: An autocrine hormone that promotes lipogenesis in human sebocytes. Proc. Natl. Acad. Sci. USA 2002, 99, 7148–7153. [Google Scholar] [CrossRef]
- Kurokawa, I.; Danby, F.W.; Ju, Q.; Wang, X.; Xiang, L.F.; Xia, L.; Chen, W.; Nagy, I.; Picardo, M.; Suh, D.H.; et al. New developments in our understanding of acne pathogenesis and treatment. Exp. Dermatol. 2009, 10, 821–832. [Google Scholar] [CrossRef]
- Smith, T.M.; Cong, Z.; Gilliland, K.L.; Clawson, G.A.; Thiboutot, D.M. Insulin-like growth factor-1 induces lipid production in human SEB-1 sebocytes via sterol response element-binding protein-1. J. Investig. Dermatol. 2006, 126, 1226–1232. [Google Scholar] [CrossRef]
- Smith, K.R.; Thiboutot, D.M. Thematic review series: Skin lipids. Sebaceous gland lipids: Friend or foe? J. Lipid Res. 2008, 49, 271–281. [Google Scholar] [CrossRef]
- Mastrofrancesco, A.; Ottaviani, M.; Cardinali, G.; Flori, E.; Briganti, S.; Ludovici, M.; Zouboulis, C.C.; Lora, V.; Camera, E.; Picardo, M. Pharmacological PPARγ modulation regulates sebogenesis and inflammation in SZ95 human sebocytes. Biochem. Pharmacol. 2017, 138, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Iwata, C.; Akimoto, N.; Sato, T.; Morokuma, Y.; Ito, A. Augmentation of lipogenesis by 15-deoxy-Delta12,14-prostaglandin J2 in hamster sebaceous glands: Identification of cytochrome P-450-mediated 15-deoxy-Delta12,14-prostaglandin J2 production. J. Investig. Dermatol. 2005, 125, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Alestas, T.; Ganceviciene, R.; Fimmel, S.; Müller-Decker, K.; Zouboulis, C.C. Enzymes involved in the biosynthesis of leukotriene B4 and prostaglandin E2 are active in sebaceous glands. J. Mol. Med. 2006, 84, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Li, X.; Zhao, S.; Zheng, X.; Zhu, W.; Zhang, J.; Meng, H.; Dong, Y. Impact of skin sensitivity mechanisms on sebum secretion: Management strategies for oily sensitive skin. J. Dermatol. Sci. Cosmet. Technol. 2024, 1, 100017. [Google Scholar] [CrossRef]
- Inui, S.; Aoshima, H.; Nishiyama, A.; Itami, S. Improvement of acne vulgaris by topical fullerene application: Unique impact on skin care. Nanomedicine 2011, 7, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Hu, X.; He, Y.; Huang, D. Lipidomics demonstrates the association of sex hormones with sebum. J. Cosmet. Dermatol. 2021, 20, 2015–2019. [Google Scholar] [CrossRef]
- Landthaler, M.; Kummermehr, J.; Wagner, A.; Plewig, G. Inhibitory effects of 13cis-reinoic acid on human sebaceous glands. Arch. Dermatol. Res. 1980, 269, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.S.; Stranieri, A.M.; Farrell, L.N.; Downing, D.T. The effect of marked inhibition of sebum production with 13cis-retinoic acid on skin surface lipid composition. J. Investig. Dermatol. 1980, 74, 66–67. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.A.; Socha-Szott, A.; Thomsen, R.J.; Pochi, P.E.; Shalita, A.R.; Strauss, J.S. Comparative effect of isotretinoin and etretinate on acne and sebaceous gland secretion. J. Am. Acad. Dermatol. 1982, 6, 760–765. [Google Scholar] [CrossRef] [PubMed]
- Guy, R.; Ridden, C.; Kealey, T. The improved organ maintenance of the human sebaceous gland: Modeling in vitro the effects of epidermal growth factor, androgens, estrogens, 13-cis retinoic acid, and phenol red. J. Investig. Dermatol. 1996, 106, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, N.; Sato, T.; Sakiguchi, T.; Kitamura, K.; Kohno, Y.; Ito, A. Cell proliferation and lipid formation in hamster sebaceous gland cells. Dermatology 2002, 204, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Kitano, T.; Koiwai, T.; Fujikawa, K.; Mori, S.; Matsumoto, T.; Sato, T. Ozenoxacin suppresses sebum production by inhibiting mTORC1 activation in differentiated hamster sebocytes. J. Dermatol. 2024, 51, 1187–1198. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar]
- Bielski, B.H.J.; Shiue, G.G.; Bajuk, S. Reduction of nitro blue tetrazolium by CO2- and O2-radicals. J. Phys. Chem. 1980, 84, 830–833. [Google Scholar] [CrossRef]
- Hsiu-Ping, L.; Daniel, B.; Creeley, D.; Grandbois, R.; Zhang, S.; Xu, C.; Yi-Fang, H.; Kathy, A.S.; Daniel, I.K.; Peter, H.S.; et al. Superoxide production by a manganese-oxidizing bacterium facilitates iodide oxidation. Appl. Environ. Microbiol. 2014, 80, 2693–2699. [Google Scholar]
- Tsikas, D. Analysis of nitrite and nitrate in biological fluids by assays based on the Griess reaction: Appraisal of the Griess reaction in the L-arginine/nitric oxide area of research. J. Chromatogr. B 2007, 851, 51–70. [Google Scholar] [CrossRef]
- Zhao, B.; Summers, F.A.; Mason, R.P. Photooxidation of Amplex Red to resorufin: Implications of exposing the Amplex Red assay to light. Free Radic. Biol. Med. 2012, 5, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Michael, F.S.; Yitong, L. A fluorescence assay for measuring acetylcholinesterase activity in rat blood and a human neuroblastoma cell line (SH-SY5Y). J. Pharmacol. Toxicol. Methods 2015, 76, 15–22. [Google Scholar]
- Basak, P.Y.; Gultekin, F.; Kilinc, I. The role of the antioxidative defense system in papulopustular acne. J. Dermatol. 2001, 28, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, A.R.; Brasaemle, D.L.; McAndrews-Hill, M.; Sztalryd, C.; Londos, C. Adoption of PERILIPIN as a unifying nomenclature for the mammalian PAT-family of intracellular lipid storage droplet proteins. J. Lipid Res. 2010, 51, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Nagai, S.; Shimizu, C.; Umetsu, M.; Taniguchi, S.; Endo, M.; Miyoshi, H.; Yoshioka, N.; Kubo, M.; Koike, T. Identification of a functional peroxisome proliferator-activated receptor responsive element within the murine perilipin gene. Endocrinology 2004, 145, 2346–2356. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, L.; Dalen, K.; Dorward, H.; Marcinkiewicz, A.; Russell, D.; Gong, D.; Londos, C.; Yamaguchi, T.; Holm, C.; et al. Activation of hormone-sensitive lipase requires two steps, protein phosphorylation and binding to the PAT-1 domain of lipid droplet coat proteins. J. Biol. Chem. 2009, 284, 32116–32125. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Takahashi, A.; Kojima, M.; Akimoto, N.; Yano, M.; Ito, A. A citrus polymethoxy flavonoid, nobiletin inhibits sebum production and sebocyte proliferation, and augments sebum excretion in hamsters. J. Investig. Dermatol. 2007, 127, 2740–2748. [Google Scholar] [CrossRef] [PubMed]
- Shimano, H. Sterol regulatory element-binding proteins (SREBPs): Transcriptional regulators of lipid synthetic genes. Prog. Lipid Res. 2001, 40, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Libby, A.E.; Bales, E.; Orlicky, D.J.; McManaman, J.L. Perilipin-2 deletion impairs hepatic lipid accumulation by interfering with sterol regulatory element-binding protein (SREBP) activation and altering the hepatic lipidome. J. Biol. Chem. 2016, 291, 24231–24246. [Google Scholar] [CrossRef]
- Tebay, L.E.; Robertson, H.; Durant, S.T.; Vitale, S.R.; Penning, T.M.; Dinkova-Kostova, A.T.; Hayes, J.D. Mechanisms of activation of the transcription factor Nrf2 by redox stressors, nutrient cues, and energy status and the pathways through which it attenuates degenerative disease. Free Radic. Biol. Med. 2015, 88 Pt B, 108–146. [Google Scholar] [CrossRef]
- Brieger, K.; Schiavone, S.; Miller, F.J., Jr.; Krause, K.-H. Reactive oxygen species: From health to disease. Swiss Med. Wkly. 2012, 142, w13659. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | 1–5 Days | 6–10 Days |
|---|---|---|
| Normal | None | None |
| Control (1) | 0.1 mM phytic acid | 0.1 mM phytic acid |
| Control (2) | 0.1 mM phytic acid | None |
| Control (3) | None | H2O2 |
| Antioxidant + H2O2 | 0.1 mM phytic acid | H2O2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokudome, Y.; Sasaki, A. Influence of Radical Generation and Elimination on Sebum Production of Hamster Sebaceous Gland Cells. Life 2025, 15, 165. https://doi.org/10.3390/life15020165
Tokudome Y, Sasaki A. Influence of Radical Generation and Elimination on Sebum Production of Hamster Sebaceous Gland Cells. Life. 2025; 15(2):165. https://doi.org/10.3390/life15020165
Chicago/Turabian StyleTokudome, Yoshihiro, and Asami Sasaki. 2025. "Influence of Radical Generation and Elimination on Sebum Production of Hamster Sebaceous Gland Cells" Life 15, no. 2: 165. https://doi.org/10.3390/life15020165
APA StyleTokudome, Y., & Sasaki, A. (2025). Influence of Radical Generation and Elimination on Sebum Production of Hamster Sebaceous Gland Cells. Life, 15(2), 165. https://doi.org/10.3390/life15020165