
Sex-Dependent Changes in Risk-Taking Predisposition of Rats Following Space Radiation Exposure

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Regulatory Compliance
2.2. Rat Demographics, Husbandry, Experimental Timeline, and Exercise Regimen
2.3. RTP Performance Screening
2.3.1. Stimulus Response Training (STR)
2.3.2. Risk-Taking Propensity (RTP) Task
2.4. Irradiation Procedure
2.5. Statistical Methods
3. Results
3.1. Pre-Exposure (Task Engagement and Learning Proficiency)
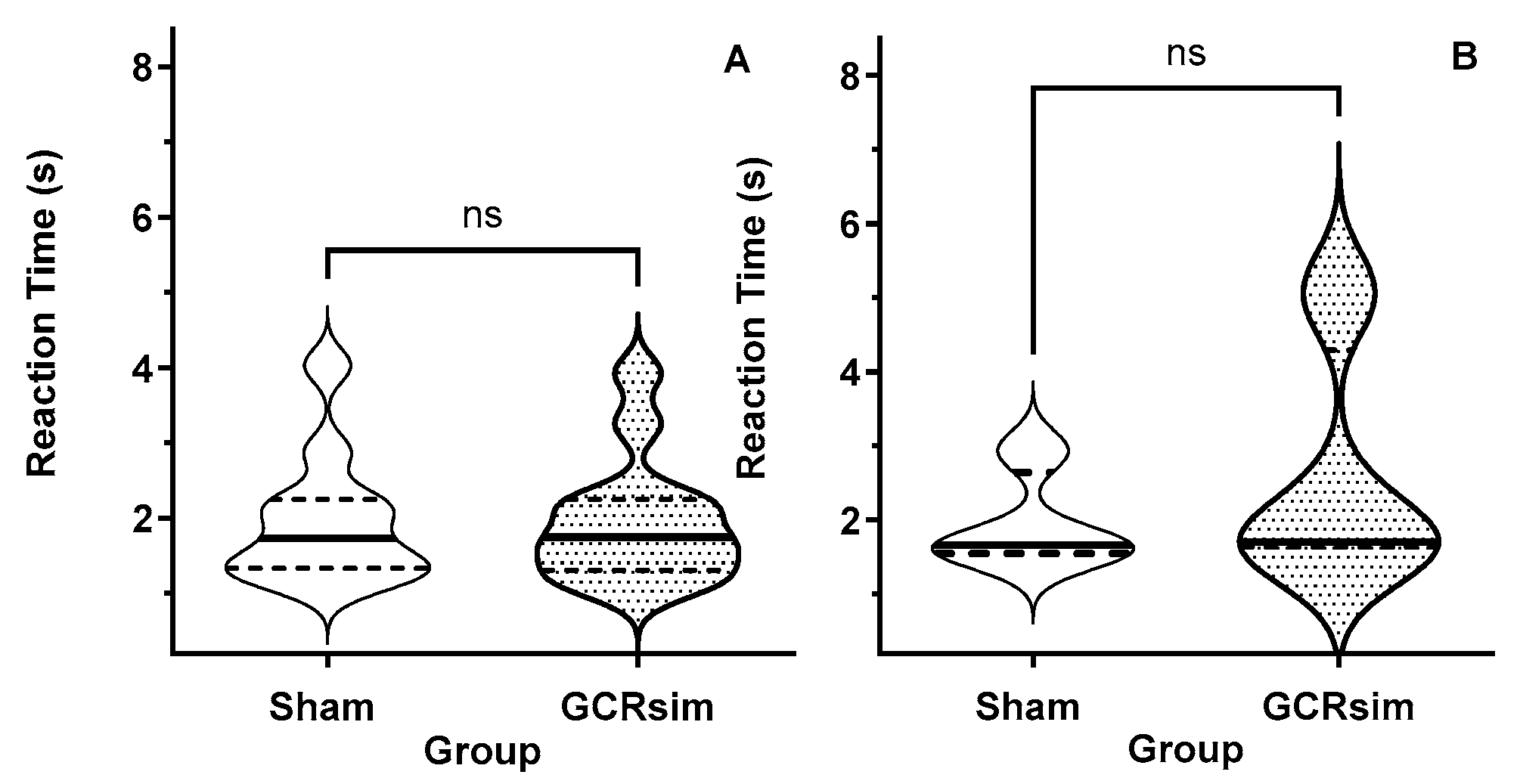
3.2. Pre-Exposure Processing Speed (Reaction Time)
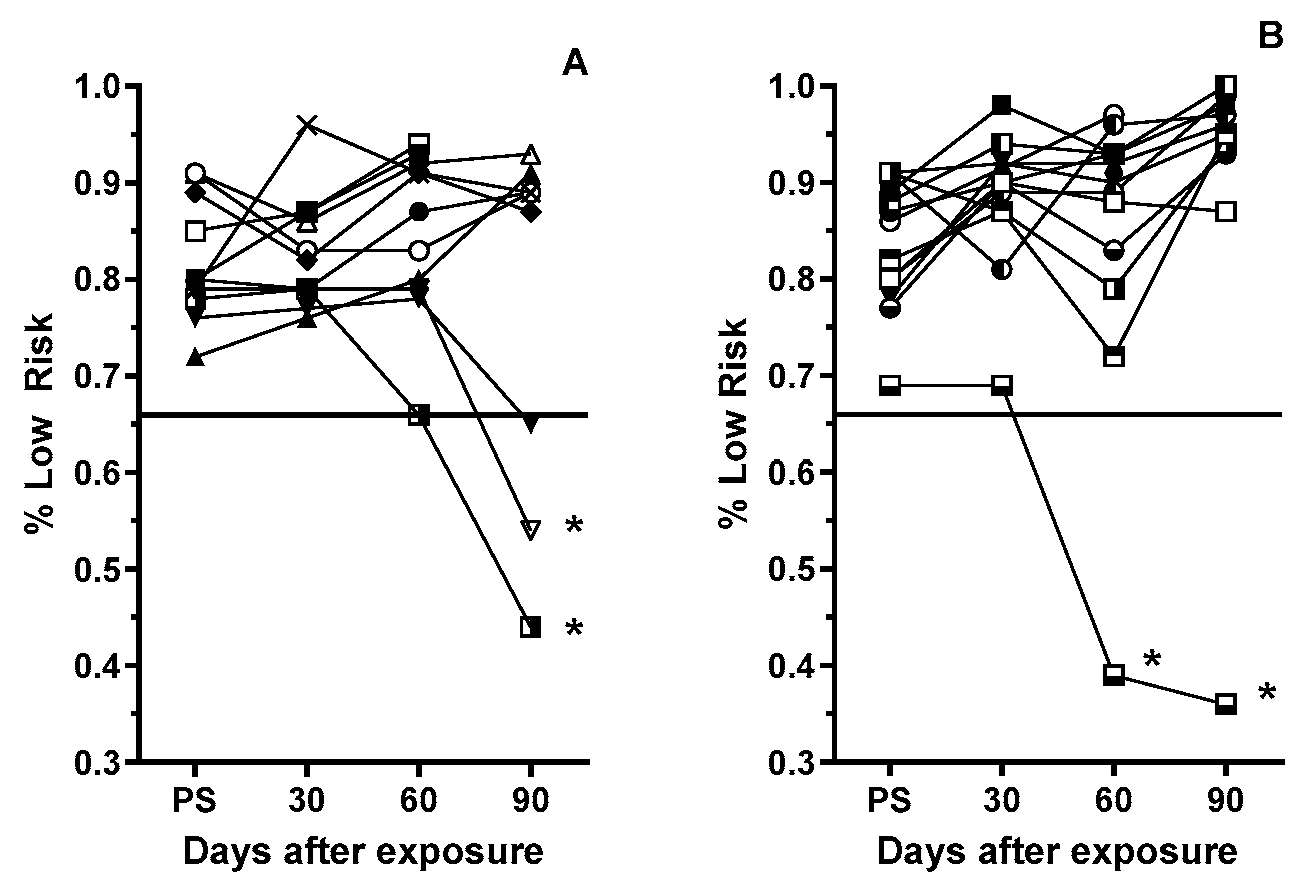
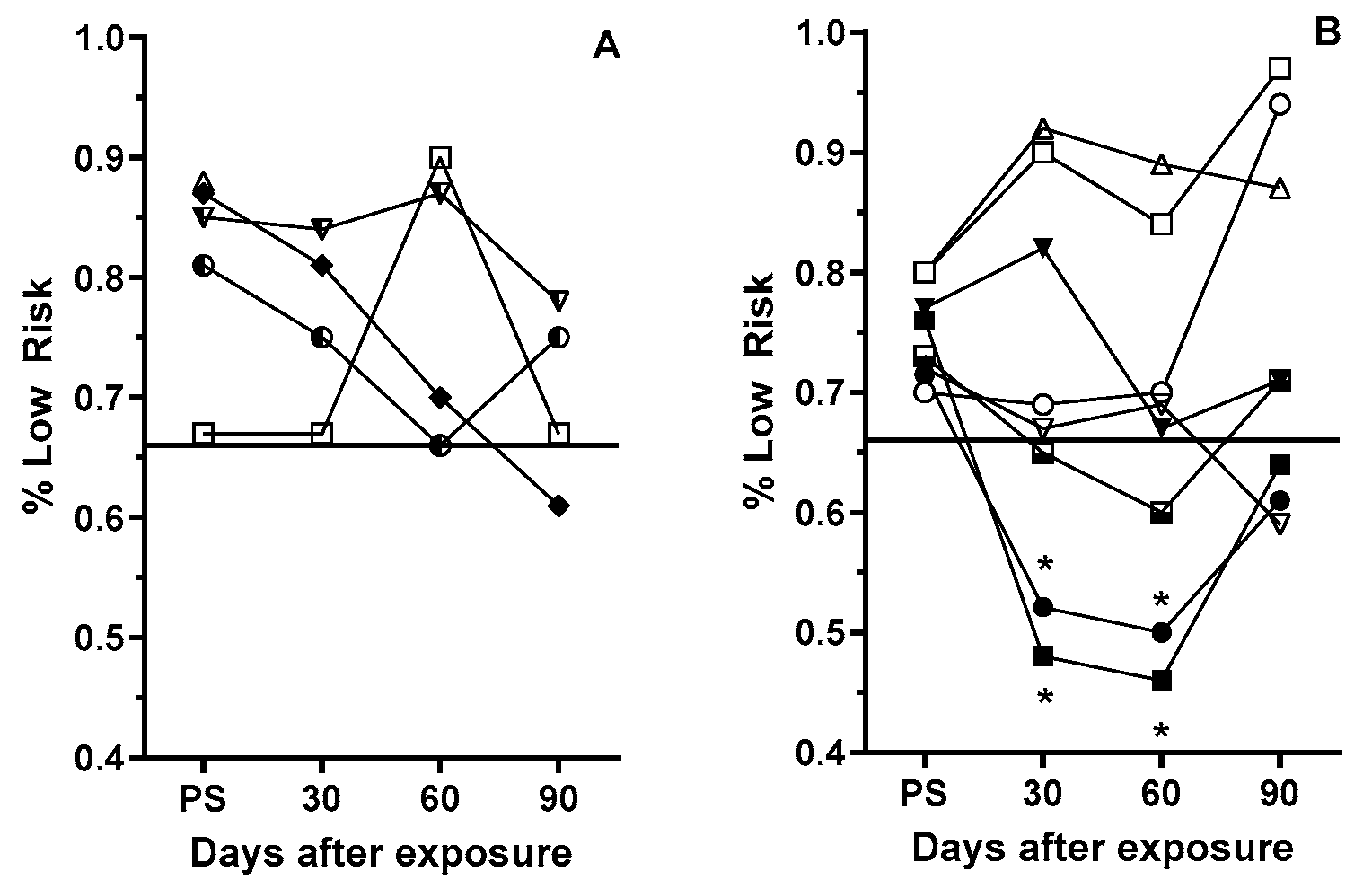
3.3. Post-Exposure Selection Choices
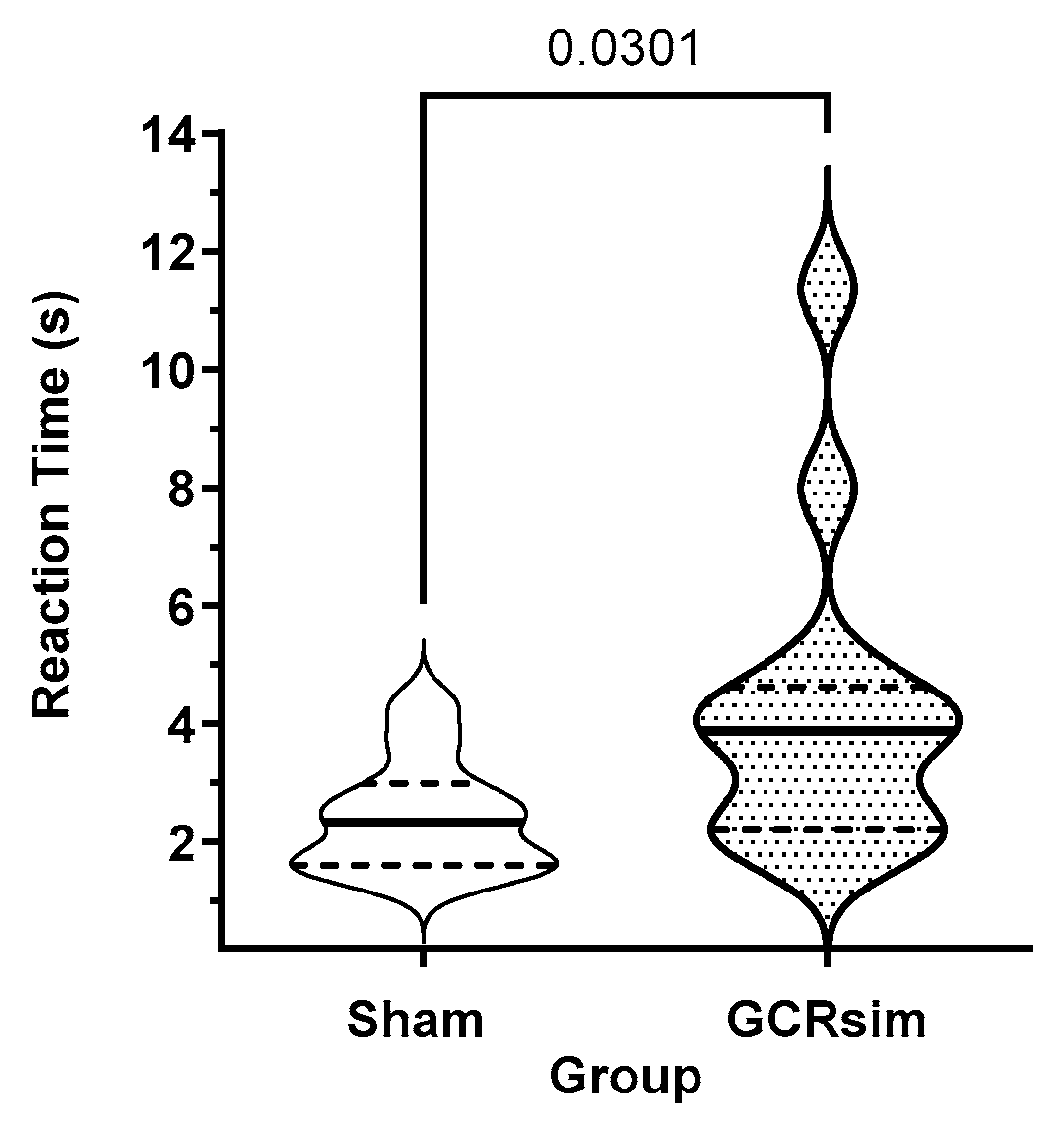
3.4. Post-Exposure Processing Speed
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nelson, G.A. Space Radiation and Human Exposures, A Primer. Radiat. Res. 2016, 185, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Wellman, L.L.; Sanford, L.D. Progressive increase in the complexity and translatability of rodent testing to assess space-radiation induced cognitive impairment. Neurosci. Biobehav. Rev. 2021, 126, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Cekanaviciute, E.; Rosi, S.; Costes, S.V. Central nervous system responses to simulated galactic cosmic rays. Int. J. Mol. Sci. 2018, 19, 3669. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, F.A.; Cacao, E. Risks of cognitive detriments after low dose heavy ion and proton exposures. Int. J. Radiat. Biol. 2019, 95, 985–998. [Google Scholar] [CrossRef]
- Kiffer, F.; Boerma, M.; Allen, A. Behavioral effects of space radiation: A comprehensive review of animal studies. Life Sci. Space Res. 2019, 21, 1–21. [Google Scholar] [CrossRef]
- Whoolery, C.W.; Yun, S.; Reynolds, R.P.; Lucero, M.J.; Soler, I.; Tran, F.H.; Ito, N.; Redfield, R.L.; Richardson, D.R.; Shih, H.Y.; et al. Multi-domain cognitive assessment of male mice reveals whole body exposure to space radiation is not detrimental to high-level cognition and actually improves pattern separation. BioRxiv 2019. [Google Scholar] [CrossRef]
- Britten, R.A.; Fesshaye, A.; Tidmore, A.; Blackwell, A.A. Similar Loss of Executive Function Performance after Exposure to Low (10 cGy) Doses of Single (4He) Ions and the Multi-Ion GCRSim Beam. Radiat. Res. 2022, 198, 375–383. [Google Scholar] [CrossRef]
- Britten, R.A.; Fesshaye, A.; Tidmore, A.; Liu, A.; Blackwell, A.A. Loss of Cognitive Flexibility Practice Effects in Female Rats Exposed to Simulated Space Radiation. Radiat. Res. 2023, 200, 256–265. [Google Scholar] [CrossRef]
- Parihar, V.K.; Allen, B.D.; Caressi, C.; Kwok, S.; Chu, E.; Tran, K.K.; Chmielewski, N.N.; Giedzinski, E.; Acharya, M.M.; Britten, R.A.; et al. Cosmic radiation exposure and persistent cognitive dysfunction. Sci. Rep. 2016, 6, 34774. [Google Scholar] [CrossRef]
- Jewell, J.S.; Duncan, V.D.; Fesshaye, A.; Tondin, A.; Macadat, E.; Britten, R.A. Exposure to ≤15 cgy of 600 mev/n 56 fe particles impairs rule acquisition but not long-term memory in the attentional set-shifting assay. Radiat. Res. 2018, 190, 565–575. [Google Scholar] [CrossRef]
- Britten, R.A.; Fesshaye, A.S.; Duncan, V.D.; Wellman, L.L.; Sanford, L.D. Sleep Fragmentation Exacerbates Executive Function Impairments Induced by Low Doses of Si Ions. Radiat. Res. 2020, 194, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Burket, J.A.; Matar, M.; Fesshaye, A.; Pickle, J.C.; Britten, R.A. Exposure to Low (≤10 cGy) Doses of 4He Particles Leads to Increased Social Withdrawal and Loss of Executive Function Performance. Radiat. Res. 2021, 196, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, S.; Liu, A.; Blackwell, A.A.; Britten, R.A. Multiple decrements in switch task performance in female rats exposed to space radiation. Behav. Brain Res. 2023, 449, 114465. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, S.; Britten, R. Simulated Space Radiation Exposure Effects on Switch Task Performance in Rats. Aerosp. Med. Hum. Perform. 2022, 93, 673–680. [Google Scholar] [CrossRef]
- Bechara, A.; Martin, E.M. Impaired decision making related to working memory deficits in individuals with substance addictions. Neuropsychology 2004, 18, 152–162. [Google Scholar] [CrossRef]
- Brand, M.; Kalbe, E.; Labudda, K.; Fujiwara, E.; Kessler, J.; Markowitsch, H.J. Decision-making impairments in patients with pathological gambling. Psychiatry Res. 2005, 133, 91–99. [Google Scholar] [CrossRef]
- Romer, D.; Betancourt, L.; Giannetta, J.M.; Brodsky, N.L.; Farah, M.; Hurt, H. Executive cognitive functions and impulsivity as correlates of risk taking and problem behavior in preadolescents. Neuropsychologia 2009, 47, 2916–2926. [Google Scholar] [CrossRef]
- Smith, M.J.; Cobia, D.J.; Wang, L.; Alpert, K.I.; Cronenwett, W.J.; Goldman, M.B.; Mamah, D.; Barch, D.M.; Breiter, H.C.; Csernansky, J.G. Cannabis-related working memory deficits and associated subcortical morphological differences in healthy individuals and schizophrenia subjects. Schizophr. Bull. 2014, 40, 287–299. [Google Scholar] [CrossRef]
- Li, N.; Phuyal, S.; Smits, E.; Reid, F.E.; Tamgue, E.N.; Arriaga, P.A.; Britten, R.A. Exposure to low (10 cGy) doses of (4)He ions leads to an apparent increase in risk taking propensity in female rats. Behav. Brain Res. 2024, 474, 115182. [Google Scholar] [CrossRef]
- Lejuez, C.W.; Read, J.P.; Kahler, C.W.; Richards, J.B.; Ramsey, S.E.; Stuart, G.L.; Strong, D.R.; Brown, R.A. Evaluation of a behavioral measure of risk taking: The Balloon Analogue Risk Task (BART). J. Exp. Psychol. Appl. 2002, 8, 75–84. [Google Scholar] [CrossRef]
- Hunt, M.K.; Hopko, D.R.; Bare, R.; Lejuez, C.W.; Robinson, E.V. Construct validity of the Balloon Analog Risk Task (BART): Associations with psychopathy and impulsivity. Assessment 2005, 12, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Lejuez, C.W.; Aklin, W.M.; Jones, H.A.; Richards, J.B.; Strong, D.R.; Kahler, C.W.; Read, J.P. The Balloon Analogue Risk Task (BART) differentiates smokers and nonsmokers. Exp. Clin. Psychopharmacol. 2003, 11, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Lejuez, C.W.; Aklin, W.M.; Zvolensky, M.J.; Pedulla, C.M. Evaluation of the Balloon Analogue Risk Task (BART) as a predictor of adolescent real-world risk-taking behaviours. J. Adolesc. 2003, 26, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Pickering, A.D.; Gray, J.A. Dopamine, appetitive reinforcement, and the neuropsychology of human learning: An individual differences approach. In Advances in Research on Temperament; Pabst Science Publishers: Lengerich, Germany, 2001; pp. 113–149. [Google Scholar]
- Slaba, T.C.; Blattnig, S.R.; Norbury, J.W.; Rusek, A.; La Tessa, C. Reference field specification and preliminary beam selection strategy for accelerator-based GCR simulation. Life Sci. Space Res. 2016, 8, 52–67. [Google Scholar] [CrossRef]
- Andreollo, N.A.; Santos, E.F.D.; de Araujo, M.R.; Lopes, L.R. Rat’s age versus human’s age: What is the relationship? Arq. Bras. Cir. Dig. 2012, 25, 49–51. [Google Scholar] [CrossRef]
- Barrus, M.M.; Winstanley, C.A. Dopamine D3 Receptors Modulate the Ability of Win-Paired Cues to Increase Risky Choice in a Rat Gambling Task. J. Neurosci. 2016, 36, 785–794. [Google Scholar] [CrossRef]
- Alaghband, Y.; Klein, P.M.; Kramár, E.A.; Cranston, M.N.; Perry, B.C.; Shelerud, L.M.; Kane, A.E.; Doan, N.-L.; Ru, N.; Acharya, M.M.; et al. Galactic cosmic radiation exposure causes multifaceted neurocognitive impairments. Cell. Mol. Life Sci. 2023, 80, 29. [Google Scholar] [CrossRef]
- Villasana, L.E.; Benice, T.S.; Raber, J. Long-term effects of 56Fe irradiation on spatial memory of mice: Role of sex and apolipoprotein E isoform. Int. J. Radiat. Oncol. Biol. Phys. 2011, 80, 567–573. [Google Scholar] [CrossRef]
- Villasana, L.; Rosenberg, J.; Raber, J. Sex-dependent effects of 56Fe irradiation on contextual fear conditioning in C57BL/6J mice. Hippocampus 2010, 20, 19–23. [Google Scholar] [CrossRef]
- Krukowski, K.; Grue, K.; Frias, E.S.; Pietrykowski, J.; Jones, T.; Nelson, G.; Rosi, S. Female mice are protected from space radiation-induced maladaptive responses. Brain Behav. Immun. 2018, 74, 106–120. [Google Scholar] [CrossRef]
- Kiffer, F.; Alexander, T.; Anderson, J.E.; Groves, T.; Wang, J.; Sridharan, V.; Boerma, M.; Allen, A.R. Late effects of 16 O-particle radiation on female social and cognitive behavior and hippocampal physiology. Radiat. Res. 2019, 191, 278–294. [Google Scholar] [CrossRef]
- Parihar, V.K.; Angulo, M.C.; Allen, B.D.; Syage, A.; Usmani, M.T.; de la Chapelle, E.P.; Amin, A.N.; Flores, L.; Lin, X.; Giedzinski, E.; et al. Sex-Specific Cognitive Deficits Following Space Radiation Exposure. Front. Behav. Neurosci. 2020, 14, 535885. [Google Scholar] [CrossRef]
- Hinkle, J.J.; Olschowka, J.A.; Love, T.M.; Williams, J.P.; O’Banion, M.K. Cranial irradiation mediated spine loss is sex-specific and complement receptor-3 dependent in male mice. Sci. Rep. 2019, 9, 18899. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Hinshaw, R.G.; Le, K.X.; Park, M.-A.; Wang, S.; Belanger, A.P.; Dubey, S.; Frost, J.L.; Shi, Q.; Holton, P.; et al. Space-like (56)Fe irradiation manifests mild, early sex-specific behavioral and neuropathological changes in wildtype and Alzheimer’s-like transgenic mice. Sci. Rep. 2019, 9, 12118. [Google Scholar] [CrossRef]
- Gaillard, A.; Rossell, S.L.; Carruthers, S.P.; Sumner, P.J.; Michie, P.T.; Woods, W.; Neill, E.; Phillipou, A.; Toh, W.L.; Hughes, M.E. Greater activation of the response inhibition network in females compared to males during stop signal task performance. Behav. Brain Res. 2020, 386, 112586. [Google Scholar] [CrossRef]
- Bolla, K.I.; Eldreth, D.A.; Matochik, J.A.; Cadet, J.L. Sex-related Differences in a Gambling Task and Its Neurological Correlates. Cereb. Cortex 2004, 14, 1226–1232. [Google Scholar] [CrossRef]
- Grissom, N.M.; Reyes, T.M. Let’s call the whole thing off: Evaluating gender and sex differences in executive function. Neuropsychopharmacology 2019, 44, 86–96. [Google Scholar] [CrossRef]
- Clark, D.K. Casualties as a Measure of the Loss of Combat Effectiveness of an Infantry Battalion; Operations Research Office, Johns Hopkins University: Baltimore, MD, USA, 1954. [Google Scholar]
- Kepecs, A.; Uchida, N.; Zariwala, H.A.; Mainen, Z.F. Neural correlates, computation and behavioural impact of decision confidence. Nature 2008, 455, 227–231. [Google Scholar] [CrossRef]
- Muller, M.D.; Fountain, S.B. Concurrent cognitive processes in rat serial pattern learning: Item memory, serial position, and pattern structure. Learn. Motiv. 2010, 41, 252–272. [Google Scholar] [CrossRef]
- Chang, H.H.V.; Fesshaye, A.S.; Tidmore, A.; Sanford, L.D.; Britten, R.A. Sleep Fragmentation Results in Novel Set-Shifting Decrements in GCR-Exposed Male and Female Rats. Radiat. Res. 2024, 203, 18–25. [Google Scholar] [CrossRef]
- Kahalley, L.S.; Peterson, R.; Ris, M.D.; Janzen, L.; Okcu, M.F.; Grosshans, D.R.; Ramaswamy, V.; Paulino, A.C.; Hodgson, D.; Mahajan, A.; et al. Superior Intellectual Outcomes After Proton Radiotherapy Compared with Photon Radiotherapy for Pediatric Medulloblastoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2020, 38, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Huynh-Le, M.-P.; Tibbs, M.D.; Karunamuni, R.; Salans, M.; Tringale, K.R.; Yip, A.; Connor, M.; Simon, A.B.; Vitzthum, L.K.; Reyes, A.; et al. Microstructural Injury to Corpus Callosum and Intrahemispheric White Matter Tracts Correlate with Attention and Processing Speed Decline After Brain Radiation. Int. J. Radiat. Oncol. 2021, 110, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Moretti, L.; Semenza, C.; Vallesi, A. General Slowing and Education Mediate Task Switching Performance Across the Life-Span. Front. Psychol. 2018, 9, 630. [Google Scholar] [CrossRef] [PubMed]
- Salthouse, T.A.; Fristoe, N.; McGuthry, K.E.; Hambrick, D.Z. Relation of task switching to speed, age, and fluid intelligence. Psychol. Aging 1998, 13, 445–461. [Google Scholar] [CrossRef]
- Jacobson, L.A.; Ryan, M.; Martin, R.B.; Ewen, J.; Mostofsky, S.H.; Denckla, M.B.; Mahone, E.M. Working memory influences processing speed and reading fluency in ADHD. Child Neuropsychol. 2011, 17, 209–224. [Google Scholar] [CrossRef]
- Shaw, P.; Eckstrand, K.; Sharp, W.; Blumenthal, J.; Lerch, J.P.; Greenstein, D.; Clasen, L.; Evans, A.; Giedd, J.; Rapoport, J.L. Attention-deficit/hyperactivity disorder is characterized by a delay in cortical maturation. Proc. Natl. Acad. Sci. USA 2007, 104, 19649–19654. [Google Scholar] [CrossRef]
- Shaw, P.; Sharp, W.S.; Morrison, M.; Eckstrand, K.; Greenstein, D.K.; Clasen, L.S.; Evans, A.C.; Rapoport, J.L. Psychostimulant treatment and the developing cortex in attention deficit hyperactivity disorder. Am. J. Psychiatry 2009, 166, 58–63. [Google Scholar] [CrossRef]
- Shaw, P.; Lalonde, F.; Lepage, C.; Rabin, C.; Eckstrand, K.; Sharp, W.; Greenstein, D.; Evans, A.; Giedd, J.N.; Rapoport, J. Development of cortical asymmetry in typically developing children and its disruption in attention-deficit/hyperactivity disorder. Arch. Gen. Psychiatry 2009, 66, 888–896. [Google Scholar] [CrossRef]
- Soler, I.; Yun, S.; Reynolds, R.P.; Whoolery, C.W.; Tran, F.H.; Kumar, P.L.; Rong, Y.; DeSalle, M.J.; Gibson, A.D.; Stowe, A.M.; et al. Multi-Domain Touchscreen-Based Cognitive Assessment of C57BL/6J Female Mice Shows Whole-Body Exposure to (56)Fe Particle Space Radiation in Maturity Improves Discrimination Learning yet Impairs Stimulus-Response Rule-Based Habit Learning. Front. Behav. Neurosci. 2021, 15, 722780. [Google Scholar] [CrossRef]
- Lu, P.H.; Lee, G.J.; Raven, E.P.; Tingus, K.; Khoo, T.; Thompson, P.M.; Bartzokis, G. Age-related slowing in cognitive processing speed is associated with myelin integrity in a very healthy elderly sample. J. Clin. Exp. Neuropsychol. 2011, 33, 1059–1068. [Google Scholar] [CrossRef]
- Lu, P.H.; Lee, G.J.; Tishler, T.A.; Meghpara, M.; Thompson, P.M.; Bartzokis, G. Myelin breakdown mediates age-related slowing in cognitive processing speed in healthy elderly men. Brain Cogn. 2013, 81, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Dickstein, D.L.; Talty, R.; Bresnahan, E.; Varghese, M.; Perry, B.; Janssen, W.G.M.; Sowa, A.; Giedzinski, E.; Apodaca, L.; Baulch, J.; et al. Alterations in synaptic density and myelination in response to exposure to high-energy charged particles. J. Comp. Neurol. 2018, 526, 2845–2855. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Event | Time Relative to SR Exposure | Rat Age |
|---|---|---|
| Arrival at EVMS | ~12 weeks pre | ~3 months |
| Exercise | ~11–3 weeks pre | |
| Ship to BNL | 1 week pre | |
| SR exposure | - | ~6 months |
| Ship to EVMS | 1 week post | |
| Exercise | 1–14 weeks post | |
| RTP testing | 14–18 weeks post | ~10 months |
| Stage 1 | Response Window | # Correct Selections | Completion Rate | Permitted # Sessions | Days at Criterion |
|---|---|---|---|---|---|
| STR15 | N/A | ≥30 | ≥60% | 8 | 1 |
| STR4 | N/A | ≥30 | ≥60% | 8 | 1 |
| STR1-Timed | 30 s | ≥30 | ≥75% | 8 | 2 |
| STR1-Fast | 10 s | ≥30 | ≥75% | 10 | 2 |
| Hole | Reward Size 1 | Win Rate | Loss Time Penalty | Possible Reward 2 |
|---|---|---|---|---|
| 6 | 1 | 90% | 5 s | 216 |
| 7 | 4 | 40% | 35 s | 102 |
| 9 | 2 | 80% | 10 s | 320 |
| 10 | 3 | 50% | 20 s | 158 |
| Male | Female | |||||||
|---|---|---|---|---|---|---|---|---|
| Sham | GCR | Sham | GCR | |||||
| TEST 1 | STRC 2 | Trials 3 | STRC | Trials | STRC | Trials | STRC | Trials |
| Pre | 3.09 ± 0.41 | 56.9 ± 2.9 | 2.69 ± 0.45 | 55.1 ± 4.4 | 2.2 ± 0.20 | 44.3 ± 2.43 | 3.12 ± 0.29 | 49.7 ± 1.51 |
| Post + 30 | 2.63 ± 0.31 | 55.6 ± 3.0 | 2.69 ± 0.33 | 51.7 ± 2.9 | 3.00 ± 0.31 | 31.2 ± 2.59 | 2.51 ± 0.19 | 35.2 ± 2.9 |
| Post + 60 | 2.45 ± 0.28 | 55.5 ± 2.5 | 2.38 ± 0.27 | 54.6 ± 2.6 | 3.1 ± 0.28 | 30.6 ± 2.5 | 2.75 ± 0.16 | 31.3 ± 2.25 |
| Post + 90 | 3.11 ± 0.48 | 48.9 ± 3.9 | 2.27 ± 0.27 | 53.2 ± 2.6 | 3.5 ± 0.50 | 30.7 ± 3.5 | 3.03 ± 0.23 | 35.8 ± 2.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smits, E.; Reid, F.E.; Tamgue, E.N.; Alvarado Arriaga, P.; Nguyen, C.; Britten, R.A. Sex-Dependent Changes in Risk-Taking Predisposition of Rats Following Space Radiation Exposure. Life 2025, 15, 449. https://doi.org/10.3390/life15030449
Smits E, Reid FE, Tamgue EN, Alvarado Arriaga P, Nguyen C, Britten RA. Sex-Dependent Changes in Risk-Taking Predisposition of Rats Following Space Radiation Exposure. Life. 2025; 15(3):449. https://doi.org/10.3390/life15030449
Chicago/Turabian StyleSmits, Elliot, Faith E. Reid, Ella N. Tamgue, Paola Alvarado Arriaga, Charles Nguyen, and Richard A. Britten. 2025. "Sex-Dependent Changes in Risk-Taking Predisposition of Rats Following Space Radiation Exposure" Life 15, no. 3: 449. https://doi.org/10.3390/life15030449
APA StyleSmits, E., Reid, F. E., Tamgue, E. N., Alvarado Arriaga, P., Nguyen, C., & Britten, R. A. (2025). Sex-Dependent Changes in Risk-Taking Predisposition of Rats Following Space Radiation Exposure. Life, 15(3), 449. https://doi.org/10.3390/life15030449