
Sucrose in Cyanobacteria: From a Salt-Response Molecule to Play a Key Role in Nitrogen Fixation

Abstract
:1. Introduction
2. Sucrose Metabolism in Cyanobacteria
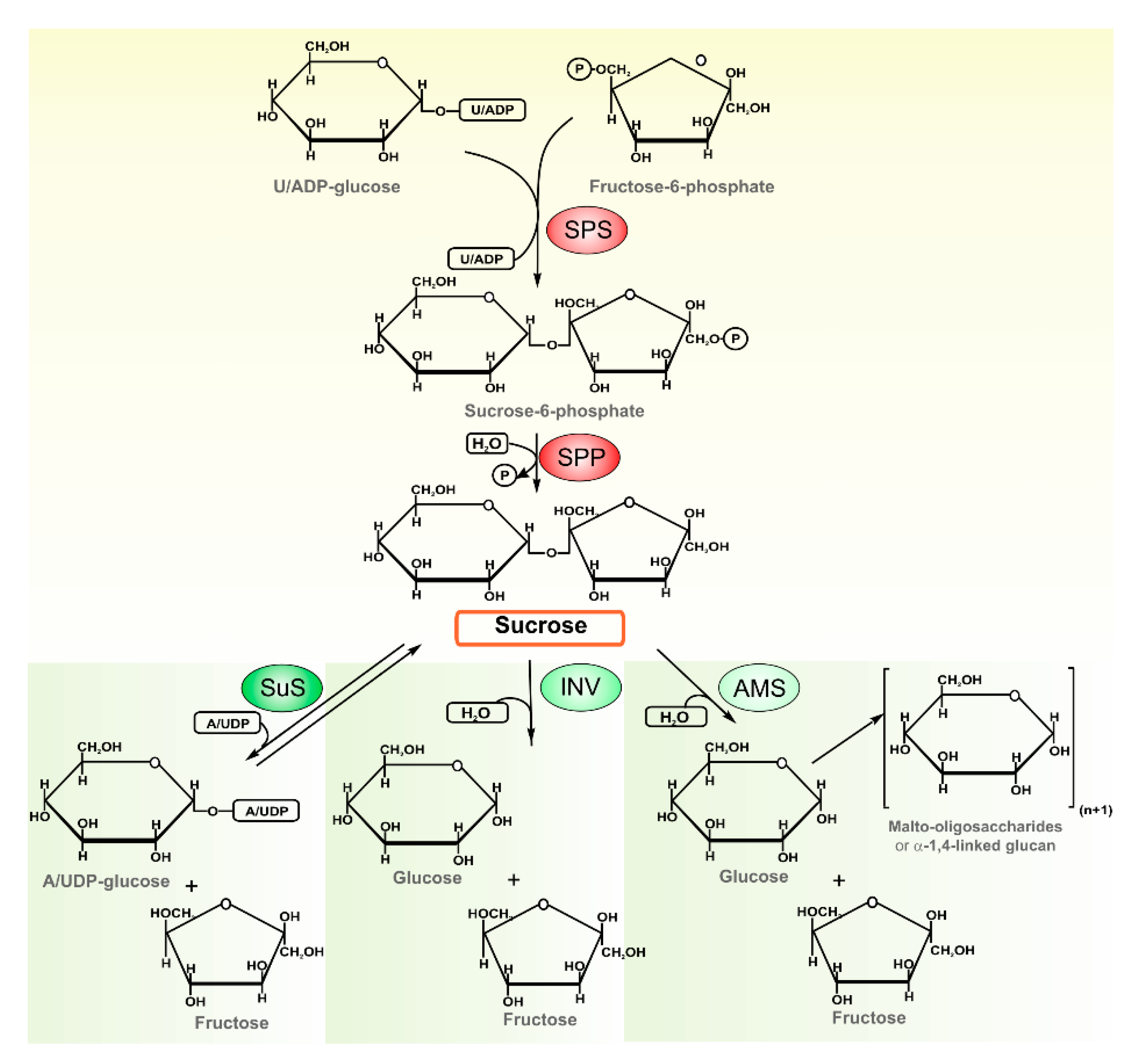
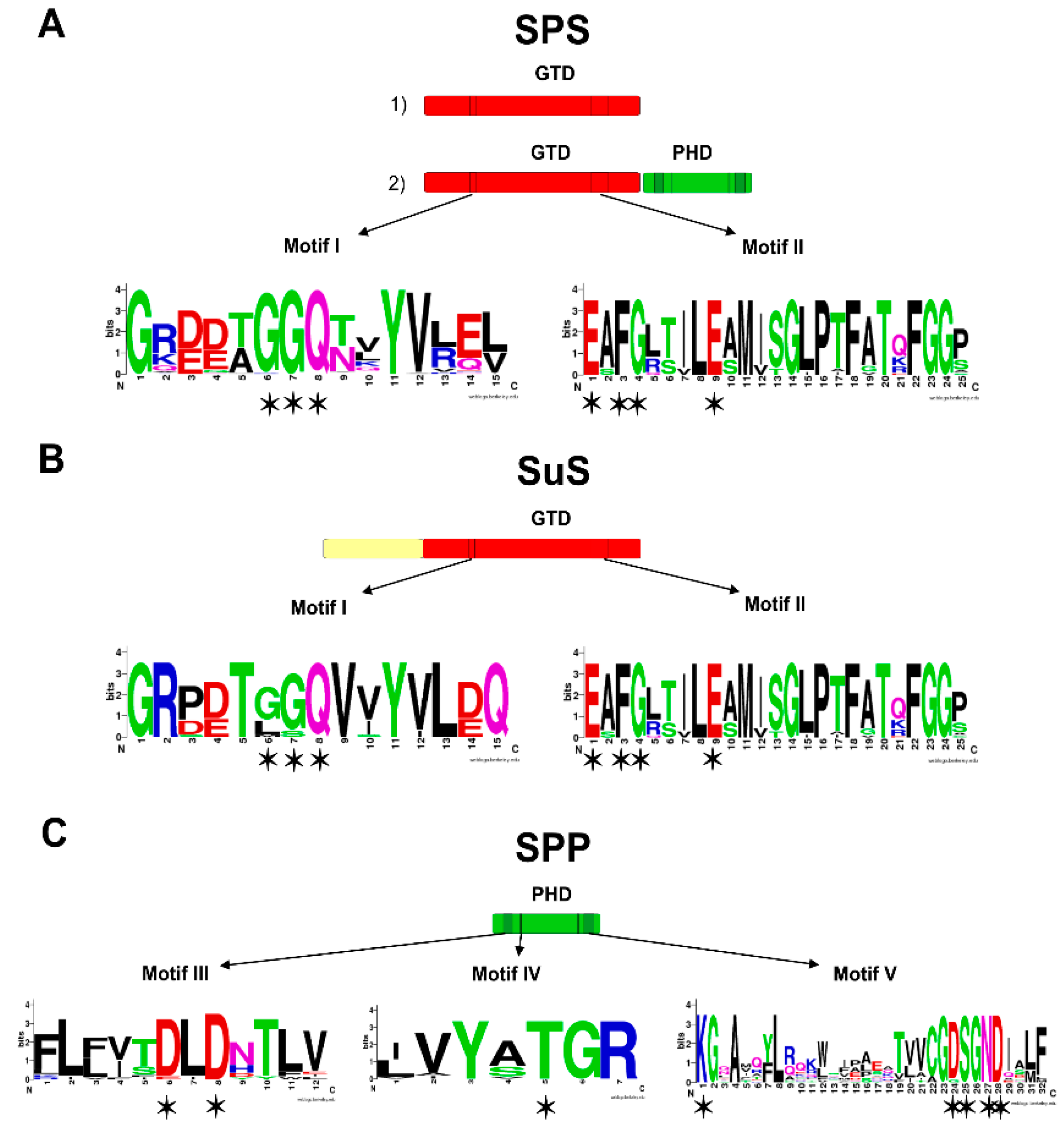
2.1. Sucrose Enzymes


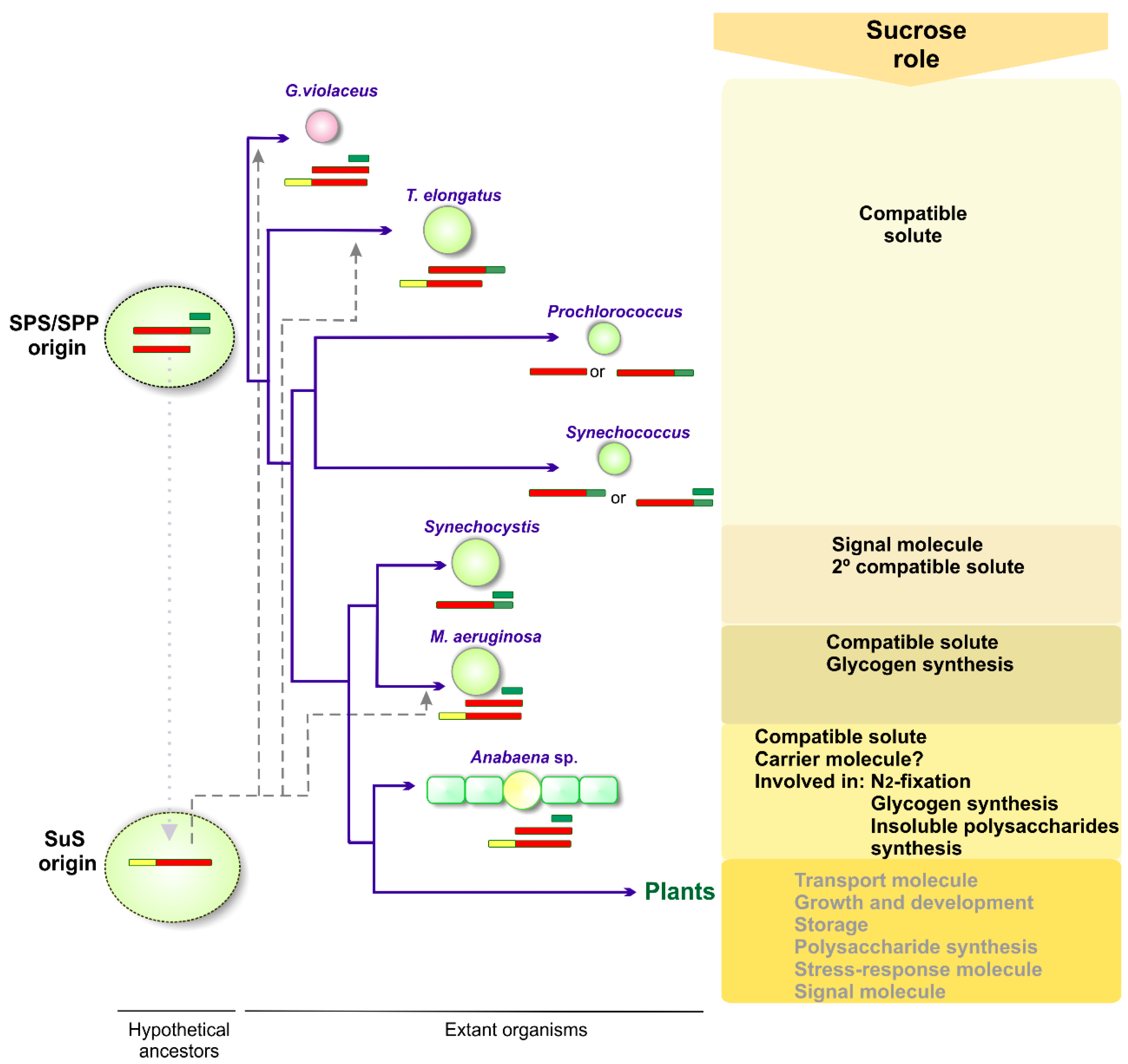
2.2. The Ancestral Origin of Sucrose Biosynthesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Strain | Habitat | Place of origin | SPS (GTD) 1 | SPS (GTD-PHD) 2 | SPP 3 | SuS 4 | A/N-Inv 5 | AMS 6 |
|---|---|---|---|---|---|---|---|---|---|
| Chroococcales | Acaryochloris marina MBIC11017 | Marine | Pacific Ocean | + | - | + | + | + | - |
| Acaryochloris sp. CCMEE 5410 | Aquatic | USA | + | - | + | + | + | - | |
| Chamaesiphon minutus PCC 6605 | Aquatic | USA | 2+ | - | + | - | - | - | |
| Crocosphaera watsonii WH 0003 | Marine | Pacific Ocean | - | - | - | - | - | - | |
| Crocosphaera watsonii WH 0003 | Marine | Pacific Ocean | - | - | - | - | - | - | |
| Crocosphaera watsonii WH 0401 | Marine | Atlantic Ocean | - | - | - | - | - | - | |
| Crocosphaera watsonii WH 8501 | Marine | Atlantic Ocean | - | - | - | - | - | - | |
| Crocosphaera watsonii WH 8501 | Marine | Atlantic Ocean | - | - | - | - | - | - | |
| Cyanobacterium aponinum PCC 10605 | Freshwater | Italy | - | - | - | - | - | - | |
| Cyanobacterium sp. UCYN-A | Marine | Pacific Ocean | - | - | - | - | - | - | |
| Cyanobacterium stanieri PCC 7202 | Freshwater | Chad | - | - | - | - | + | - | |
| Cyanobium gracile PCC 6307 | Aquatic | USA | + | + | 2+ | - | - | + | |
| Cyanobium sp. PCC 7001 | Marine | USA | + | + | + | - | + | - | |
| Cyanothece sp. BH63E, ATCC 51472 | Aquatic | USA | - | - | + | - | - | - | |
| Cyanothece sp. BH68, ATCC 51142 | Marine | USA | - | - | + | - | - | - | |
| Cyanothece sp. CCY 0110 | Marine | Tanzania | - | - | + | - | - | - | |
| Cyanothece sp. PCC 7424 | Freshwater | Senegal | 2+ | - | + | + | + | - | |
| Cyanothece sp. PCC 7425 | Freshwater | Senegal | 2+ | - | + | 2+ | - | - | |
| Cyanothece sp. PCC 7822 | Freshwater | India | + | - | + | - | + | - | |
| Cyanothece sp. PCC 8801 | Freshwater | Taiwan | - | - | - | - | - | - | |
| Cyanothece sp. PCC 8802 | Freshwater | Taiwan | - | - | - | - | - | - | |
| Dactylococcopsis salina PCC 8305 | Aquatic | Israel | + | - | + | + | + | - | |
| Geminocystis herdmanii PCC 6308 | Aquatic | USA | - | - | - | - | - | - | |
| Gloeobacter kilaueensis JS1 | Freshwater | Hawaii | + | - | + | + | + | - | |
| Gloeobacter violaceus PCC 7421 | Freshwater | Switzerland | + | - | + | + | - | - | |
| Gloeocapsa sp. PCC 73106 | Freshwater | Switzerland | + | - | - | - | - | - | |
| Gloeocapsa sp. PCC 7428 | Freshwater | Sri Lanka | - | - | + | 2+ | + | - | |
| Halothece sp. PCC 7418 | Freshwater | Israel | 2+ | - | + | + | - | - | |
| Microcystis aeruginosa DIANCHI905/PCC7806 | Freshwater | China/ Netherlands | + | - | + | + | - | - | |
| Microcystis aeruginosa NIES-843 | Freshwater | Japan | - | - | - | - | - | - | |
| Microcystis aeruginosa PCC 7941 | Freshwater | Canada | - | - | - | - | - | - | |
| Microcystis aeruginosa PCC 9432 | Freshwater | Canada | - | - | - | - | - | - | |
| Microcystis aeruginosa PCC 9443 | Freshwater | Africa | - | - | - | - | - | - | |
| Microcystis aeruginosa PCC 9701 | Freshwater | France | - | - | - | - | - | - | |
| Microcystis aeruginosa PCC 9717 | Freshwater | France | - | - | - | - | - | - | |
| Microcystis aeruginosa PCC 9806 | Freshwater | USA | - | - | - | - | - | - | |
| Microcystis aeruginosa PCC 9807 | Freshwater | South Africa | - | - | - | - | - | - | |
| Microcystis aeruginosa PCC 9808 | Freshwater | Australia | - | - | - | - | - | - | |
| Microcystis aeruginosa PCC 9809 | Freshwater | USA | - | - | - | - | - | - | |
| Microcystis aeruginosa SPC777 | Freshwater | Brazil | - | - | - | - | - | - | |
| Microcystis aeruginosa TAIHU98 | Freshwater | China | - | - | - | - | - | - | |
| Microcystis sp. T1-4 | Freshwater | Thailand | - | - | - | - | - | - | |
| Prochlorococcus marinus AS9601 | Marine | Arabian Sea | + | - | - | - | - | - | |
| Prochlorococcus marinus CCMP 1375 | Marine | Atlantic Ocean | + | - | - | - | + | - | |
| Prochlorococcus marinus CCMP 1986 | Marine | Mediterranean Sea | + | - | - | - | + | - | |
| Prochlorococcus marinus MIT 9211 | Marine | Pacific Ocean | + | - | - | - | + | - | |
| Prochlorococcus marinus MIT 9215 | Marine | Pacific Ocean | + | - | - | - | + | - | |
| Prochlorococcus marinus MIT 9301 | Aquatic | Sargasso Sea | + | - | - | - | + | - | |
| Prochlorococcus marinus MIT 9303 | Aquatic | Sargasso Sea | - | + | - | - | + | - | |
| Prochlorococcus marinus MIT 9312 | Marine | Gulf Stream | + | - | - | - | + | - | |
| Prochlorococcus marinus MIT 9313 | Marine | Gulf Stream | - | + | - | - | + | - | |
| Prochlorococcus marinus MIT 9515 | Marine | Pacific Ocean | + | - | - | - | + | - | |
| Prochlorococcus marinus MIT9202 | Marine | Pacific Ocean | + | - | - | - | + | - | |
| Prochlorococcus marinus NATL1A | Marine | Atlantic Ocean | - | + | - | - | + | - | |
| Prochlorococcus marinus NATL2A | Marine | Atlantic Ocean | - | + | - | - | + | - | |
| Prochlorococcus sp. CC9311 | Marine | USA | - | + | - | - | + | - | |
| Prochlorococcus sp. CC9605 | Marine | USA | - | + | - | - | + | - | |
| Prochlorococcus sp. CC9902 | Marine | USA | - | + | - | - | + | - | |
| Prochlorococcus sp. W10 | Marine | Pacific Ocean | - | - | - | - | + | - | |
| Prochlorococcus sp. W11 | Marine | Pacific Ocean | - | - | - | - | + | - | |
| Prochlorococcus sp. W12 | Marine | Pacific Ocean | - | - | - | - | - | - | |
| Prochlorococcus sp. W2 | Marine | Pacific Ocean | + | - | - | - | - | - | |
| Prochlorococcus sp. W3 | Marine | Pacific Ocean | - | - | - | - | - | - | |
| Prochlorococcus sp. W4 | Marine | Pacific Ocean | - | - | - | - | + | - | |
| Prochlorococcus sp. W5 | Marine | Pacific Ocean | - | - | - | - | - | - | |
| Prochlorococcus sp. W6 | Marine | Pacific Ocean | - | - | - | - | - | - | |
| Prochlorococcus sp. W7 | Marine | Pacific Ocean | + | - | - | - | + | - | |
| Prochlorococcus sp. W8 | Marine | Pacific Ocean | + | - | - | - | + | - | |
| Prochlorococcus sp. W9 | Marine | Pacific Ocean | - | - | - | - | + | - | |
| Prochlorococcus sp. WH 7803 | Marine | Sargasso Sea | - | + | - | - | + | - | |
| Prochlorococcus sp. WH8102 | Marine | Atlantic Ocean | - | + | - | - | + | - | |
| Prochlorothrix hollandica PCC 9006 | Freshwater | Netherlands | - | - | - | - | - | - | |
| Rubidibacter lacunae KORDI 51-2 | Marine | Micronesia | + | - | + | + | - | - | |
| Synechococcus elongatus PCC 6301 | Freshwater | USA | - | + | - | - | + | - | |
| Synechococcus elongatus PCC 7942 | Freshwater | USA | - | + | - | - | + | - | |
| Synechococcus sp. BL107 | Marine | Mediterranean Sea | - | + | - | - | + | - | |
| Synechococcus sp. CB0101 | Marine | USA | - | + | + | - | + | - | |
| Synechococcus sp. CB0205 | Marine | USA | - | + | + | - | + | - | |
| Synechococcus sp. CC9616 | Marine | Pacific Ocean | - | - | - | - | + | - | |
| Synechococcus sp. JA-2-3B'a(2-13) | Freshwater | USA | - | - | + | - | + | - | |
| Synechococcus sp. JA-3-3Ab | Freshwater | USA | - | - | + | - | + | - | |
| Synechococcus sp. KORDI-100 | Marine | South Korea | + | - | - | - | + | -- | |
| Synechococcus sp. KORDI-49 | Marine | South Korea | + | - | - | - | + | - | |
| Synechococcus sp. KORDI-52 | Marine | South Korea | - | + | - | - | + | - | |
| Synechococcus sp. PCC 6312 | Marine | USA | - | + | - | - | + | - | |
| Synechococcus sp. PCC 7002 | Marine | Atlantic Ocean | - | + | - | - | - | + | |
| Synechococcus sp. PCC 7003 | Marine | USA | - | + | + | - | - | + | |
| Synechococcus sp. PCC 7117 | Marine | Asustralia | - | + | + | - | - | + | |
| Synechococcus sp. PCC 73109 | Marine | USA | - | + | + | - | - | + | |
| Synechococcus sp. PCC 7335 | Marine | Mexico | - | - | - | - | - | - | |
| Synechococcus sp. PCC 7336 | Marine | USA | - | - | - | - | - | - | |
| Synechococcus sp. PCC 7502 | Freshwater | Switzerland | + | - | + | - | + | - | |
| Synechococcus sp. PCC 8807 | Freshwater | Gabon | - | + | - | - | - | + | |
| Synechococcus sp. RCC 307 | Marine | Mediterranean Sea | + | - | + | - | + | - | |
| Synechococcus sp. RS9916 | Marine | Israel | + | + | - | - | + | - | |
| Synechococcus sp. RS9917 | Marine | Israel | - | + | - | - | + | - | |
| Synechococcus sp. WH 8016 | Marine | USA | - | + | - | - | + | - | |
| Synechococcus sp. WH 8109 | Marine | Sargasso Sea | - | + | - | - | + | - | |
| Synechococcus sp. WH5701 | Marine | USA | + | - | + | - | + | + | |
| Synechococcus sp. WH7805 | Marine | Sargasso Sea | - | + | - | - | - | - | |
| Synechocystis sp. PCC 6308 | Freshwater | USA | - | + | + | - | - | - | |
| Synechocystis sp. PCC 6803 | Freshwater | USA | - | + | + | - | + | - | |
| Synechocystis sp. PCC 7509 | Freshwater | Switzerland | - | - | + | + | + | - | |
| Thermosynechococcus elongatus BP-1 | Freshwater | Japan | - | + | + | + | - | - | |
| Oscillatoriales | Arthrospira maxima CS-328 | Freshwater | Chad | - | - | - | + | - | - |
| Arthrospira platensis C1 | Freshwater | Chad | - | - | - | + | - | - | |
| Arthrospira platensis NIES-39 | Freshwater | Chad | - | - | - | + | - | - | |
| Arthrospira platensis Paraca | Freshwater | Peru | - | - | - | + | - | - | |
| Arthrospira sp. PCC 8005 | Freshwater | India, Kenya, Mexico or Peru | - | - | - | 2+ | - | - | |
| Crinalium epipsammum PCC 9333 | Unknown | Unknown | 2+ | - | + | + | + | - | |
| Cyanobacterium sp. ESFC-1 | Freshwater | USA | 2+ | - | + | + | - | - | |
| Geitlerinema sp. PCC 7105 | Freshwater | USA | 2+ | - | + | + | - | - | |
| Geitlerinema sp. PCC 7407 | Freshwater | Unknown | + | - | 2+ | + | + | - | |
| Leptolyngbya boryana PCC 6306 | Freshwater | USA | 2+ | - | + | 2+ | 2+ | - | |
| Leptolyngbya sp. 2LT21S03 | Desert soil | Israel | - | - | + | - | - | - | |
| Leptolyngbya sp. PCC 6406 | Freshwater | USA | - | - | - | - | - | - | |
| Leptolyngbya sp. PCC 7375 | Freshwater | USA | - | - | - | - | - | - | |
| Leptolyngbya sp. PCC 7376 | Freshwater | USA | - | - | - | - | - | - | |
| Lyngbya majuscula 3L | Marine | Netherlands Antilles | + | - | + | 2+ | + | - | |
| Lyngbya sp. CCY 8106 | Freshwater | Germany | - | - | - | - | - | - | |
| Microcoleus chthonoplastes PCC 7420 | Marine | USA | + | - | + | + | + | - | |
| Microcoleus sp. PCC 7113 | Soil | USA | 4+? | - | + | 2+ | 2+ | - | |
| Microcoleus vaginatus FGP-2 | Desert soil | USA | + | - | + | + | - | - | |
| Microcoleus vaginatus PCC 9802 | Soil crusts | USA | + | - | + | + | - | - | |
| Nodosilinea nodulosa PCC 7104 | Soil | USA | - | - | - | - | - | - | |
| Oscillatoria acuminata PCC 6304 | Soil | USA | 2+ | + | + | + | 2+ | - | |
| Oscillatoria formosa PCC 6407 | Freshwater | USA | - | - | - | - | - | - | |
| Oscillatoria nigro-viridis PCC 7112 | Soil | USA | + | - | + | + | - | - | |
| Oscillatoria sp. PCC 10802 | Freshwater | Unknown | - | - | - | - | - | - | |
| Oscillatoria sp. PCC 6506 | Freshwater | Unknown | - | - | - | - | - | - | |
| Oscillatoriales sp. JSC-1 | Freshwater | USA | - | - | - | - | - | - | |
| Oscillatoriales sp. JSC-12 | Freshwater | USA | 2+ | - | + | + | 2+ | - | |
| Planktothrix agardhii NIVA-CYA 126/8 | Freshwater | Finland | - | - | - | - | - | - | |
| Planktothrix agardhii NIVA-CYA 34 | Freshwater | Norway | - | - | - | - | - | - | |
| Planktothrix agardhii NIVA-CYA 56/3 | Freshwater | Norway | - | - | - | - | - | - | |
| Planktothrix NIVA-CYA405 | Freshwater | Norway | - | - | - | - | - | - | |
| Planktothrix NIVA-CYA406 | Freshwater | Norway | - | - | - | - | - | - | |
| Planktothrix prolifica NIVA-CYA 540 (Draft1) | Freshwater | Norway | - | - | - | - | - | - | |
| Planktothrix rubescens NIVA-CYA 98 | Freshwater | Norway | - | - | - | - | - | - | |
| Planktothrix sp. 585 | Freshwater | Germany | - | - | - | - | - | - | |
| Planktothrix sp. NIVA CYA 15 | Freshwater | Norway | - | - | - | - | - | - | |
| Planktothrix sp. NIVA-CYA 407 | Freshwater | Norway | - | - | - | - | - | - | |
| Planktothrix sp. st147 | Freshwater | Germany | - | - | - | - | - | - | |
| Pseudanabaena sp. PCC 6802 | Freshwater | USA | + | - | + | + | + | - | |
| Pseudanabaena sp. PCC 7367 | Marine | USA | - | - | + | + | - | - | |
| Pseudanabaena sp. PCC 7429 | Freshwater | Switzerland | - | - | + | - | - | - | |
| Spirulina major PCC 6313 | Brackish water | USA | - | - | - | - | - | - | |
| Spirulina subsalsa PCC 9445 | Freshwater | Italy | - | - | - | - | - | - | |
| Trichodesmium erythraeum IMS101 | Marine | USA | + | - | - | - | - | - | |
| Pleurocapsales | Chroococcidiopsis sp. PCC 6712 | Freshwater | USA | - | - | + | - | - | - |
| Chroococcidiopsis thermalis PCC 7203 | Soil | Germany | 2+ | - | + | + | 2+ | - | |
| Pleurocapsa sp. PCC 7319 | Freshwater | Mexico | + | - | + | - | - | - | |
| Pleurocapsa sp. PCC 7327 | Freshwater | USA | + | - | + | 2+ | 2+ | - | |
| Stanieria cyanosphaera PCC 7437 | Freshwater | Cuba | 2+ | - | - | + | 2+ | - | |
| Xenococcus sp. PCC 7305 | Marine | USA | - | - | + | - | - | - | |
| Nostocales | Anabaena circinalis AWQC131C | Freshwater | Australia | + | - | + | - | - | - |
| Anabaena cylindrica PCC 7122 | Freshwater | United Kingdom | + | + | + | 2+ | + | - | |
| Anabaena sp. 90 | Freshwater | Finland | + | + | 2+ | + | + | - | |
| Anabaena sp. PCC 7108 | Freshwater | USA | + | + | + | + | + | - | |
| Anabaena variabilis ATCC 29413 | Freshwater | USA | 2+ | - | + | 2+ | + | - | |
| Calothrix desertica PCC 7102 | Sand | Chile | + | + | + | 2+ | 4+ | + | |
| Calothrix sp. PCC 6303 | Freshwater | USA | + | + | + | + | + | - | |
| Calothrix sp. PCC 7103 | Freshwater | USA | + | + | + | 2+ | 4+ | + | |
| Calothrix sp. PCC 7507 | Freshwater | Switzerland | 2+ | - | + | 2+ | + | - | |
| Cylindrospermopsis raciborskii CS-505 | Freshwater | Australia | + | - | + | - | + | - | |
| Cylindrospermopsis raciborskii CS-506 | Freshwater | Australia | + | - | + | - | + | - | |
| Cylindrospermopsis raciborskii CS-509 | Freshwater | Australia | + | - | + | - | + | - | |
| Cylindrospermum stagnale PCC 7417 | Soil | Sweden | + | + | + | 2+ | + | - | |
| Fremyella diplosiphon UTEX 481 | Freshwater | USA | - | + | 2+ | 3+ | 2+ | - | |
| Microchaete sp. PCC 7126 | Freshwater | USA | 2+ | + | 2+ | + | - | ||
| Nodularia spumigena CCY9414 | Brackish water | Denmark | + | + | + | 2+ | 2+ | - | |
| Nostoc azollae 0708 | Symbiont | Unkown | 2+ | - | + | + | + | - | |
| Nostoc punctiforme PCC 73102 | Freshwater | Australia | 2+ | - | 2+ | 2+ | 2+ | - | |
| Nostoc sp. PCC 7107 | Freshwater | USA | + | 2+ | 2+ | 2+ | - | ||
| Nostoc sp. PCC 7120 | Freshwater | USA | 2+ | - | + | 2+ | 2+ | - | |
| Nostoc sp. PCC 7524 | Freshwater | Sri Lanka | + | + | + | 2+ | 2+ | - | |
| Raphidiopsis brookii D9 | Freshwater | Brazil | - | - | - | - | - | - | |
| Rivularia sp. PCC 7116 | Freshwater | USA | + | - | + | 2+ | + | - | |
| Scytonema hofmanni PCC 7110 | Freshwater | Bermuda | 2+ | - | + | 2+ | 2+ | - | |
| Scytonema hofmanni UTEX 2349 | Freshwater | USA | + | - | + | 2+ | + | - | |
| Stigonematales | Chlorogloeopsis fritschii PCC 6912 | Freshwater | India | + | - | + | 2+ | 2+ | - |
| Chlorogloeopsis fritschii PCC 7702 | Soil | India | + | - | + | 2+ | - | ||
| Chlorogloeopsis sp. PCC 9212 | Freshwater | Spain | + | - | + | 2+ | 2+ | - | |
| Fischerella muscicola PCC 7414 | Freshwater | New Zealand | + | - | 2+ | 3+ | 2+ | - | |
| Fischerella muscicola SAG 1427-1 | Freshwater | India | - | - | - | 2+ | + | - | |
| Fischerella sp. JSC-11 | Freshwater | USA | + | - | + | 2+ | + | - | |
| Fischerella sp. PCC 9339 | Freshwater | Unknown | + | - | + | - | 2+ | - | |
| Fischerella sp. PCC 9431 | Freshwater | Unknown | 2+ | - | 2+ | + | + | - | |
| Fischerella sp. PCC 9605 | Freshwater | Israel | + | - | 2+ | 3+ | 2+ | - | |
| Fischerella thermalis PCC 7521 | Freshwater | USA | + | - | + | 3+ | + | - | |
| Mastigocladopsis repens PCC 10914 | Soil | Spain | 2+ | - | + | 2+ | + | - | |
| Mastigocoleus testarum BC008 | Marine | Puerto Rico | + | - | + | + | 2+ | - |
3. Sucrose Roles in Cyanobacteria
3.1. Sucrose as a Compatible Solute
3.2. Sucrose and Nitrogen Fixation
4. Final Remarks

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Whitton, B.A.; Potts, M. Introduction to the cyanobacteria. In Ecology of Cyanobacteria II; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 1–13. [Google Scholar]
- Reed, R.H.; Stewart, W.D.P. The responses of cyanobacteria to salt stress. In Biochemistry of the Algae and Cyanobacteria; Rogers, L.J., Gallon, J.R., Eds.; Clarendon Press: Oxford, UK, 1988; Chapter 12; pp. 217–231. [Google Scholar]
- Hagemann, M. Molecular biology of cyanobacterial salt acclimation. FEMS Microbiol. Rev. 2011, 35, 87–123. [Google Scholar] [CrossRef] [PubMed]
- Blumwald, E.; Tel-Or, E. Osmoregulation and cell composition in salt-adaptation of Nostoc. Muscorum. Arch. Microbiol. 1982, 132, 168–172. [Google Scholar] [CrossRef]
- Reed, R.H.; Richardson, D.L.; Warr, S.R.C.; Stewart, W.D.P. Carbohydrate accumulation and osmotic stress in cyanobacteria. J. Gen. Microbiol. 1984, 130, 1–4. [Google Scholar]
- Reed, R.H.; Chudek, J.A.; Foster, R.; Stewart, W.D.P. Osmotic adjustment in cyanobacteria from hypersaline environments. Arch. Microbiol. 1984, 138, 333–337. [Google Scholar] [CrossRef]
- Schilling, N.; Ehrnsperger, K. Cellular differentiation of sucrose metabolism in Anabaena variabilis. Z. Naturforsch. 1985, 40, 776–779. [Google Scholar]
- Salerno, G.L.; Curatti, L. Origin of sucrose metabolism in higher plants: When, how and why? Trends Plant Sci. 2003, 8, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Cumino, A.C.; Perez-Cenci, M.; Giarrocco, L.E.; Salerno, G.L. The proteins involved in sucrose synthesis in the marine cyanobacterium Synechococcus sp. PCC 7002 are encoded by two genes transcribed from a gene cluster. FEBS Lett. 2010, 584, 4655–4660. [Google Scholar] [CrossRef] [PubMed]
- Kolman, M.A.; Torres, L.L.; Martin, M.L.; Salerno, G.L. Sucrose synthase in unicellular cyanobacteria and its relationship with salt and hypoxic stress. Planta 2012, 235, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Noel, G.; Cumino, A.C.; Kolman, M.A.; Salerno, G.L. First evidence of sucrose biosynthesis by single cyanobacterial bimodular proteins. FEBS Lett. 2013, 587, 1669–1674. [Google Scholar] [CrossRef] [PubMed]
- Winter, H.; Huber, S.C. Regulation of sucrose metabolism in higher plants: Localization and regulation of activity of key enzymes. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 253–289. [Google Scholar] [CrossRef] [PubMed]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant. Biol. 2004, 7, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Porchia, A.C.; Salerno, G.L. Sucrose biosynthesis in a prokaryotic organism: Presence of two sucrose-phosphate synthases in Anabaena with remarkable differences compared with the plant enzymes. Proc. Natl. Acad. Sci. USA 1996, 93, 13600–13604. [Google Scholar] [CrossRef] [PubMed]
- Curatti, L.; Folco, E.; Desplats, P.; Abratti, G.; Limones, V.; Herrera-Estrella, L.; Salerno, G. Sucrose-phosphate synthase from Synechocystis sp. strain PCC 6803: Identification of the spsA gene and characterization of the enzyme expressed in Escherichia coli. J. Bacteriol. 1998, 180, 6776–6779. [Google Scholar] [PubMed]
- Lunn, J.E.; Price, G.D.; Furbank, R.T. Cloning and expression of a prokaryotic sucrose-phosphate synthase gene from the cyanobacterium Synechocystis sp. PCC 6803. Plant Mol. Biol. 1999, 40, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Cumino, A.; Ekeroth, C.; Salerno, G.L. Sucrose-phosphate phosphatase from Anabaena sp. strain PCC 7120: Isolation of the protein and gene revealed significant structural differences from the higher-plant enzyme. Planta 2001, 214, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Cumino, A.; Curatti, L.; Giarrocco, L.; Salerno, G.L. Sucrose metabolism: Anabaena sucrose-phosphate synthase and sucrose-phosphate phosphatase define minimal functional domains shuffled during evolution. FEBS Lett. 2002, 517, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Cumino, A.C.; Marcozzi, C.; Barreiro, R.; Salerno, G.L. Carbon cycling in Anabaena sp. PCC 7120. Sucrose synthesis in the heterocysts and possible role in nitrogen fixation. Plant Phys. 2007, 143, 1385–1397. [Google Scholar] [CrossRef]
- Curatti, L.; Giarrocco, L.E.; Cumino, A.C.; Salerno, G.L. Sucrose synthase is involved in the conversion of sucrose to polysaccharides in filamentous nitrogen-fixing cyanobacteria. Planta 2008, 228, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Vargas, W.A.; Salerno, G.L. The Cinderella story of sucrose hydrolysis: Alkaline/neutral invertases, from cyanobacteria to unforeseen roles in plant cytosol and organelles. Plant Sci. 2010, 178, 1–8. [Google Scholar] [CrossRef]
- Perez-Cenci, M.; Salerno, G.L. Functional characterization of Synechococcus amylosucrase and fructokinase encoding genes discovers two novel actors on the stage of cyanobacterial sucrose metabolism. Plant Sci. 2014, 224, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Chua, T.K.; Bujnicki, J.M.; Tan, T.C.; Huynh, F.; Patel, B.K.; Sivaraman, J. The structure of sucrose phosphate synthase from Halothermothrix orenii reveals its mechanism of action and binding mode. Plant Cell 2008, 20, 1059–1072. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Anderson, S.; Zhang, Y.; Garavito, R.M. The structure of sucrose synthase-1 from Arabidopsis thaliana and its functional implications. J. Biol. Chem. 2010, 286, 36108–36118. [Google Scholar] [CrossRef]
- Fieulaine, S.; Lunn, J.E.; Borel, F.; Ferrer, J. The structure of a cyanobacterial sucrose-phosphatase reveals the sugar tongs that release free sucrose in the cell. Plant Cell 2005, 17, 2049–2058. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. Weblogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Blank, C.E. Phylogenetic distribution of compatible solute synthesis genes support a freshwater origin for cyanobacteria. J. Phycol. 2013, 49, 880–895. [Google Scholar] [CrossRef]
- Nagao, M.; Uemura, M. Sucrose phosphate phosphatase in the green alga Klebsormidium flaccidum (Streptophyta) lacks an extensive C-terminal domain and differs from that of land plants. Planta 2012, 235, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acid Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Makarova, K.S.; Aravind, L. Horizontal gene transfer in prokaryotes: Quantification and classification. Ann. Rev. Microbiol. 2001, 55, 709–742. [Google Scholar] [CrossRef]
- Barratt, D.H.P.; Derbyshire, P.; Findlay, K.; Pike, M.; Wellner, N.; Lunn, J.; Feil, R.; Simpson, C.; Maule, A.J.; Smith, A.M. Normal growth of Arabidopsis requires cytosolic invertase but not sucrose synthase. Proc. Natl. Acad. Sci. USA 2009, 106, 13124–13129. [Google Scholar] [CrossRef] [PubMed]
- Kolman, M.A.; Salerno, G.L. Sucrose in bloom-forming cyanobacteria: Gain and loss of genes involved in its biosynthesis. Environ. Microbiol. 2015. submitted for publication. [Google Scholar]
- Hagemann, M.; Erdmann, N. Environmental stresses. In Cyanobacterial Nitrogen Metabolism and Environmental Biotechnology; Rai, A.K., Ed.; Springer Verlag Narosa Publishing House: New Delhi, India, 1997; pp. 155–221. [Google Scholar]
- Oren, A. Salts and Brines. In Ecology of Cyanobacteria II; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 401–426. [Google Scholar]
- Higo, A.; Katoh, H.; Ohmori, K.; Ikeuchi, M.; Ohmori, M. The role of a gene cluster for trehalose metabolism in dehydration tolerance of the filamentous cyanobacterium Anabaena sp. PCC 7120. Microbiology 2006, 152, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Reed, R.H.; Stewart, W.D.P. Osmotic adjustment and organic solute accumulation in unicellular cyanobacteria from freshwater and marine habitats. Mar. Biol. 1985, 88, 1–9. [Google Scholar] [CrossRef]
- Klahn, S.; Hagemann, M. Compatible solute biosynthesis in cyanobacteria. Environ. Microbiol. 2011, 13, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Potts, M. Desiccation tolerance: A simple process? Trends Microbiol. 2001, 9, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Blumwald, E.; Mehlhorn, R.J.; Packer, L. Studies of osmoregulation in salt adaptation of cyanobacteria with ESR spin-probe techniques. Proc. Natl. Acad. Sci. USA 1983, 80, 2599–2602. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, N. Organic osmoregulatory solutes in blue-green algae. Z. Pflanzenphysiol. 1983, 110, 147–155. [Google Scholar] [CrossRef]
- Page-Sharp, M.; Behm, C.A.; Smith, G.D. Involvement of the compatible solutes trehalose and sucrose in the response to salt stress of a cyanobacterial Scytonema species isolated from desert soils. Biochim. Biophys. Acta 1999, 1472, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, M.; Marin, K. Salt-induced sucrose accumulation is mediated by sucrose-phosphate-synthase in cyanobacteria. J. Plant. Phys. 1999, 155, 424–430. [Google Scholar] [CrossRef]
- Salerno, G.L.; Instituto de Investigaciones en Biodiversidad y Biotecnología (INBIOTEC-CONICET) and Fundación para Investigaciones Biológicas Aplicadas (FIBA), Mar del Plata, Argentina. Personal Communication, 2014.
- Salerno, G.L.; Porchia, A.C.; Vargas, W.; Abdian, P.L. Fructose-containing oligosaccharides: Novel compatible solutes in Anabaena cells exposed to salt stress. Plant Sci. 2004, 167, 1003–1008. [Google Scholar] [CrossRef]
- Desplats, P.; Folco, E.; Salerno, G.L. Sucrose may play an additional role to that of an osmolyte in Synechocystis sp. PCC 6803 salt-shocked cells. Plant Phys. Biochem. 2005, 43, 133–138. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Black, T.A.; Jager, K.; Panoff, J.-M.; Wolk, C.P. Regulation of an osmoticum-responsive gene in Anabaena sp. strain PCC 7120. J. Bacteriol. 1998, 180, 6332–6337. [Google Scholar] [PubMed]
- Ehira, S.; Kimura, S.; Miyazaki, S.; Ohmori, M. Sucrose synthesis is controlled by the two-component response regulator OrrA in the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120. App. Environ. Microbiol. 2014, 5672–5679. [Google Scholar]
- Pontis, H.G.; Vargas, W.A.; Salerno, G.L. Structural characterization of the members of a polymer series, compatible solutes in Anabaena cells exposed to salt stress. Plant Sci. 2007, 172, 29–35. [Google Scholar] [CrossRef]
- Miao, X.; Wu, Q.; Wu, G.; Zhao, N. Sucrose accumulation in salt-stressed cells of agp gene deletion mutant in cyanobacterium Synechocystis sp. PCC 6803. FEMS Microbiol. Lett. 2003, 218, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Ludden, P.; Barris, R.H. Biochemical basis of plant breeding. In Nitrogen Metabolism; Neyra, C.A., Ed.; CRC Press: Boca Raton, FL, USA, 1986; Volume 2, pp. 41–59. [Google Scholar]
- Berman-Frank, I.; Lundgren, P.; Falkowski, P. Nitrogen fixation and photosynthetic oxygen evolution in cyanobacteria. Res. Microbiol. 2003, 154, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Buikema, W.J.; Haselkorn, R. Isolation and complementation of nitrogen fixation mutants of the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 1991, 173, 1879–1885. [Google Scholar] [PubMed]
- Wolk, P.C. Heterocyst formation in Anabaena. In Prokaryotic Development; Brun, Y.V., Shimkets, L.J., Eds.; American Society for Microbiology: Washington, DC, USA, 2000; pp. 83–104. [Google Scholar]
- Yoon, H.-S.; Golden, J.W. PatS and products of nitrogen fixation control heterocyst pattern. J. Bacteriol. 2001, 183, 2605–2613. [Google Scholar] [CrossRef] [PubMed]
- Golden, J.W.; Yoon, H.-S. Heterocyst development in Anabaena. Curr. Opin. Microbiol. 2003, 6, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Wolk, C.P. Movement of carbon from vegetative cells to heterocysts in Anabaena cylindrica. J. Bacteriol. 1968, 96, 2138–2143. [Google Scholar] [PubMed]
- Wolk, C.P.; Ernst, A.; Elhai, J. Heterocyst metabolism and development. In The Molecular Biology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 769–823. [Google Scholar]
- Zhang, C.C.; Laurent, S.; Sakr, S.; Peng, L.; Bedu, S. Heterocyst differentiation and pattern formation in cyanobacteria: A chorus of signals. Mol. Biol. 2006, 59, 367–375. [Google Scholar]
- Privalle, L.S.; Burris, R.H. Adenine nucleotide levels in and nitrogen fixation by the cyanobacterium Anabaena sp. strain 7120. J. Bacteriol. 1983, 154, 351–355. [Google Scholar] [PubMed]
- Curatti, L.; Giarrocco, L.; Salerno, G.L. Sucrose synthase and RuBisCo expression is similarly regulated by the nitrogen source in the nitrogen-fixing cyanobacterium Anabaena sp. Planta 2006, 223, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Curatti, L.; Flores, E.; Salerno, G. Sucrose is involved in the diazotrophic metabolism of the heterocyst-forming cyanobacterium Anabaena sp. FEBS Lett. 2002, 513, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Vargas, W.; Cumino, A.; Salerno, G.L. Cyanobacterial alkaline/neutral invertases. Origin of sucrose hydrolysis in the plant cytosol? Planta 2003, 216, 951–960. [Google Scholar] [PubMed]
- Vargas, W.A.; Nishi, C.N.; Giarrocco, L.E.; Salerno, G.L. Differential roles of alkaline/neutral invertases in Nostoc sp. PCC 7120: Inv-B isoform is essential for diazotrophic growth. Planta 2011, 233, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Igual, R.; Flores, E.; Herrero, A. Inactivation of a heterocyst-specific invertase indicates a principal role of sucrose catabolism in heterocysts of Anabaena sp. J. Bacteriol. 2011, 192, 5526–5533. [Google Scholar] [CrossRef]
- Fay, P. Factors influencing dark nitrogen fixation in a blue-green alga. App. Environ. Microbiol. 1976, 31, 376–379. [Google Scholar]
- Lockau, W.; Peterson, R.B.; Wolk, C.P.; Burris, R.H. Modes of reduction of nitrogenase in heterocysts isolated from Anabaena species. Biochimica et Biophysica Acta (BBA)—Bioenergetics 1978, 502, 298–308. [Google Scholar] [CrossRef]
- Ernst, A.; Boger, P. Glycogen accumulation and the induction of nitrogenase activity in the heterocyst-forming cyanobacterium Anabaena variabilis. J. Gen. Microbiol. 1985, 131, 3147–3153. [Google Scholar]
- Ernst, A.; Reich, S.; Boger, P. Modification of dinitrogenase reductase in the cyanobacterium Anabaena variabilis due to C starvation and ammonia. J. Bacteriol. 1990, 172, 748–755. [Google Scholar] [PubMed]
- Vargas, W.; Martin, M.L.; Salerno, G. Myths and Facts on Cytosolic Sucrose Hydrolysis. In Sucrose: Properties, Biosynthesis and Health Implications; Magazu, S., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2013; pp. 155–176. [Google Scholar]
- Jensen, B.B.; Cox, R.P.; Burris, R.H. Isolation of cyanobacterial heterocysts with high and sustained dinitrogen-fixation capacity supported by endogenous reductants. Arch. Microbiol. 1986, 145, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Luque, I.; Forchhammer, K. Nitrogen Assimilation and C/N Balance Sensing. In The Cyanobacteria: Molecular Biology, Genomics and Evolution; Herrero, A., Flores, E., Eds.; Caister Academic Press: Norfolk, UK, 2008; pp. 335–382. [Google Scholar]
- Marcozzi, C.; Cumino, A.C.; Salerno, G.L. Role of NtcA, a cyanobacterial global nitrogen regulator, in the regulation of sucrose metabolism gene expression in Anabaena sp. PCC 7120. Arch. Microbiol. 2009, 191, 255–263. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolman, M.A.; Nishi, C.N.; Perez-Cenci, M.; Salerno, G.L. Sucrose in Cyanobacteria: From a Salt-Response Molecule to Play a Key Role in Nitrogen Fixation. Life 2015, 5, 102-126. https://doi.org/10.3390/life5010102
Kolman MA, Nishi CN, Perez-Cenci M, Salerno GL. Sucrose in Cyanobacteria: From a Salt-Response Molecule to Play a Key Role in Nitrogen Fixation. Life. 2015; 5(1):102-126. https://doi.org/10.3390/life5010102
Chicago/Turabian StyleKolman, María A., Carolina N. Nishi, Macarena Perez-Cenci, and Graciela L. Salerno. 2015. "Sucrose in Cyanobacteria: From a Salt-Response Molecule to Play a Key Role in Nitrogen Fixation" Life 5, no. 1: 102-126. https://doi.org/10.3390/life5010102
APA StyleKolman, M. A., Nishi, C. N., Perez-Cenci, M., & Salerno, G. L. (2015). Sucrose in Cyanobacteria: From a Salt-Response Molecule to Play a Key Role in Nitrogen Fixation. Life, 5(1), 102-126. https://doi.org/10.3390/life5010102