
Adsorption of Nucleic Acid Bases, Ribose, and Phosphate by Some Clay Minerals

Abstract
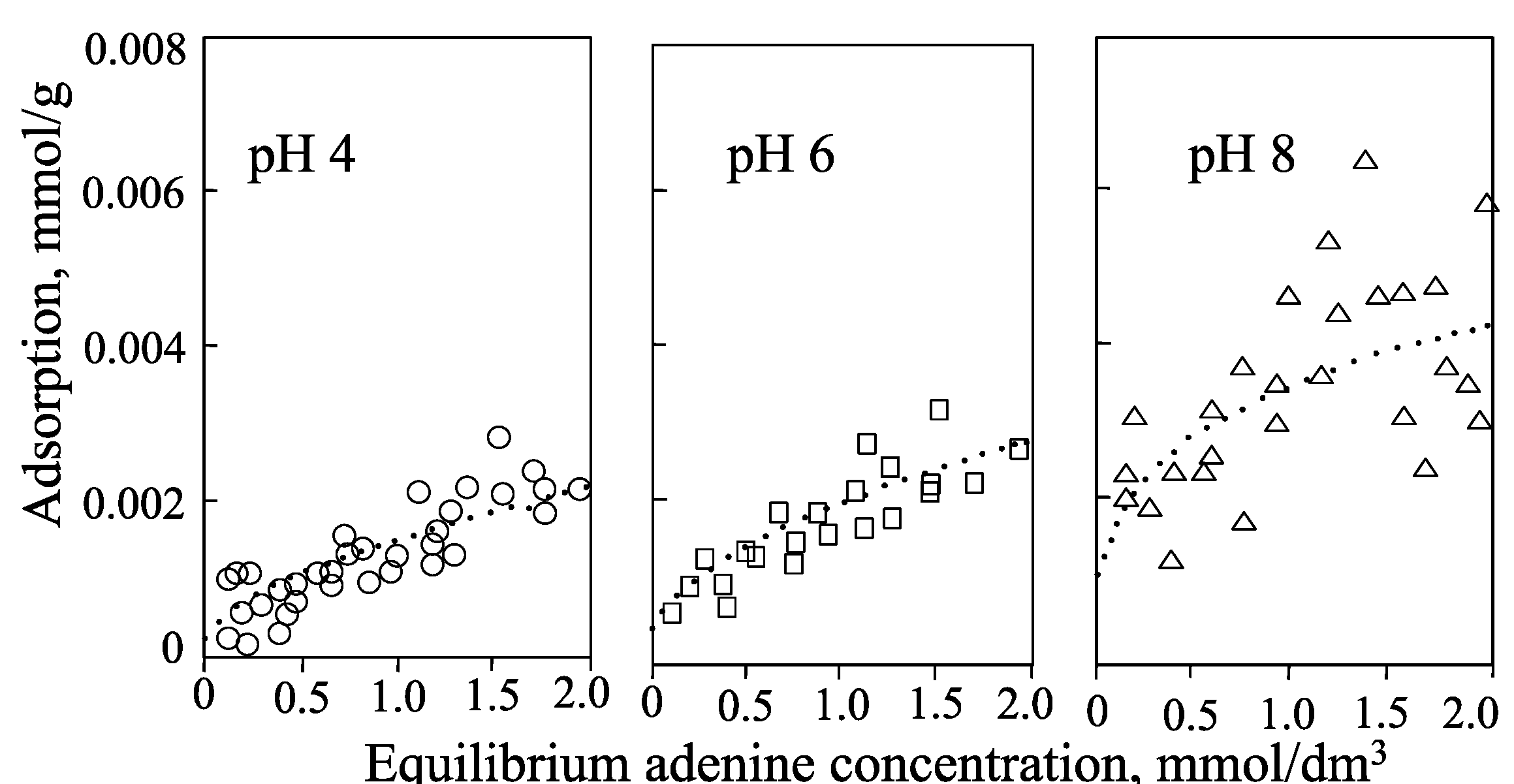
:1. Introduction
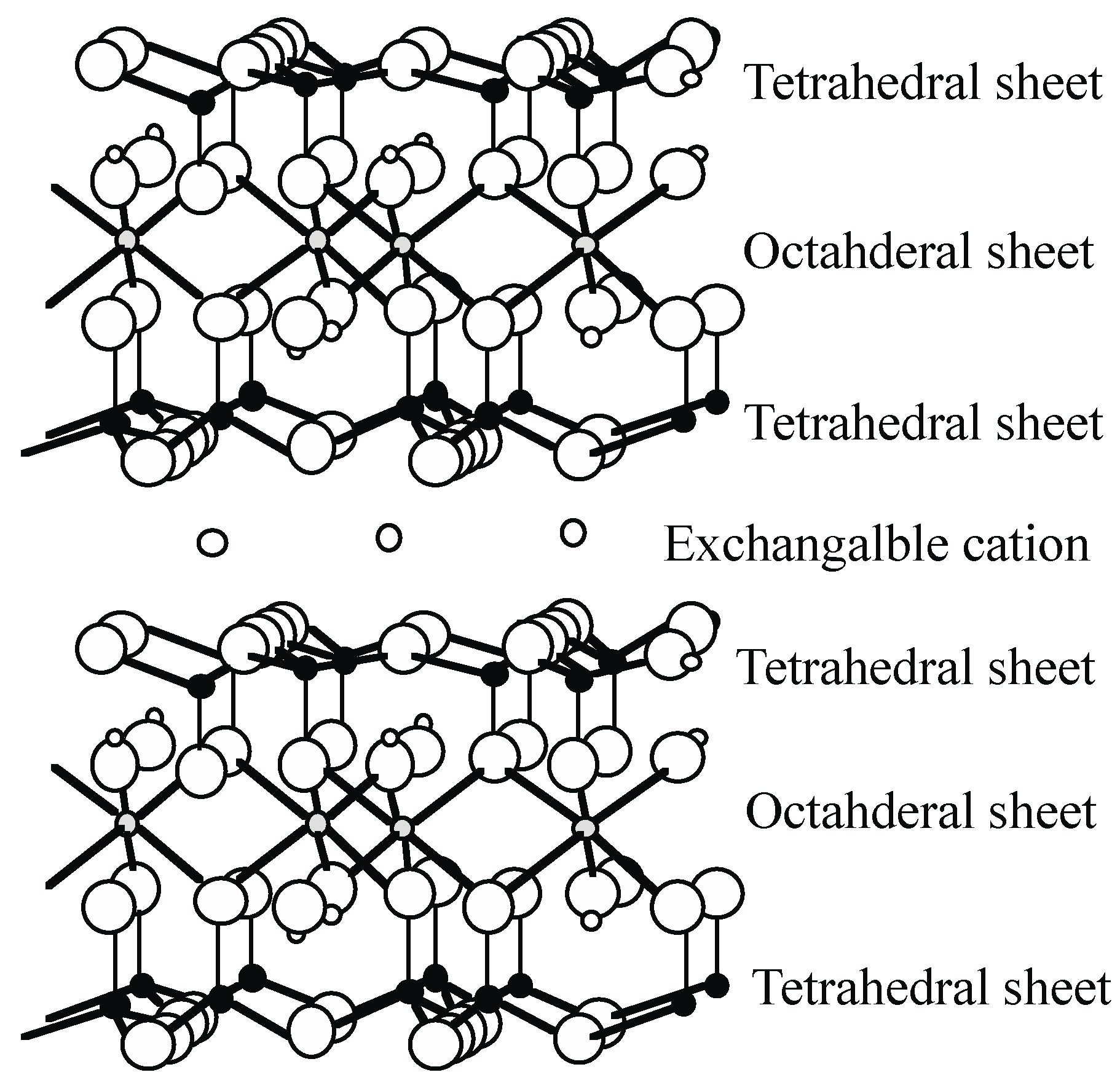
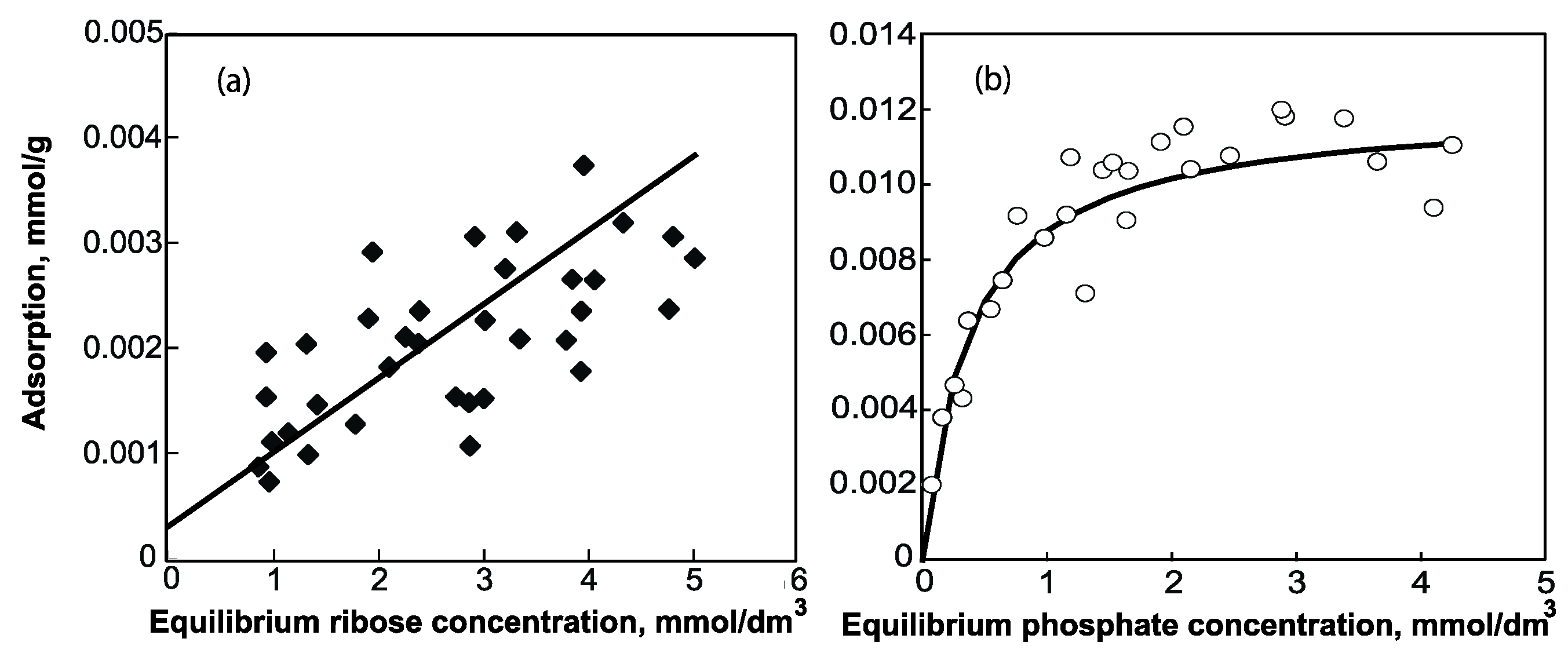
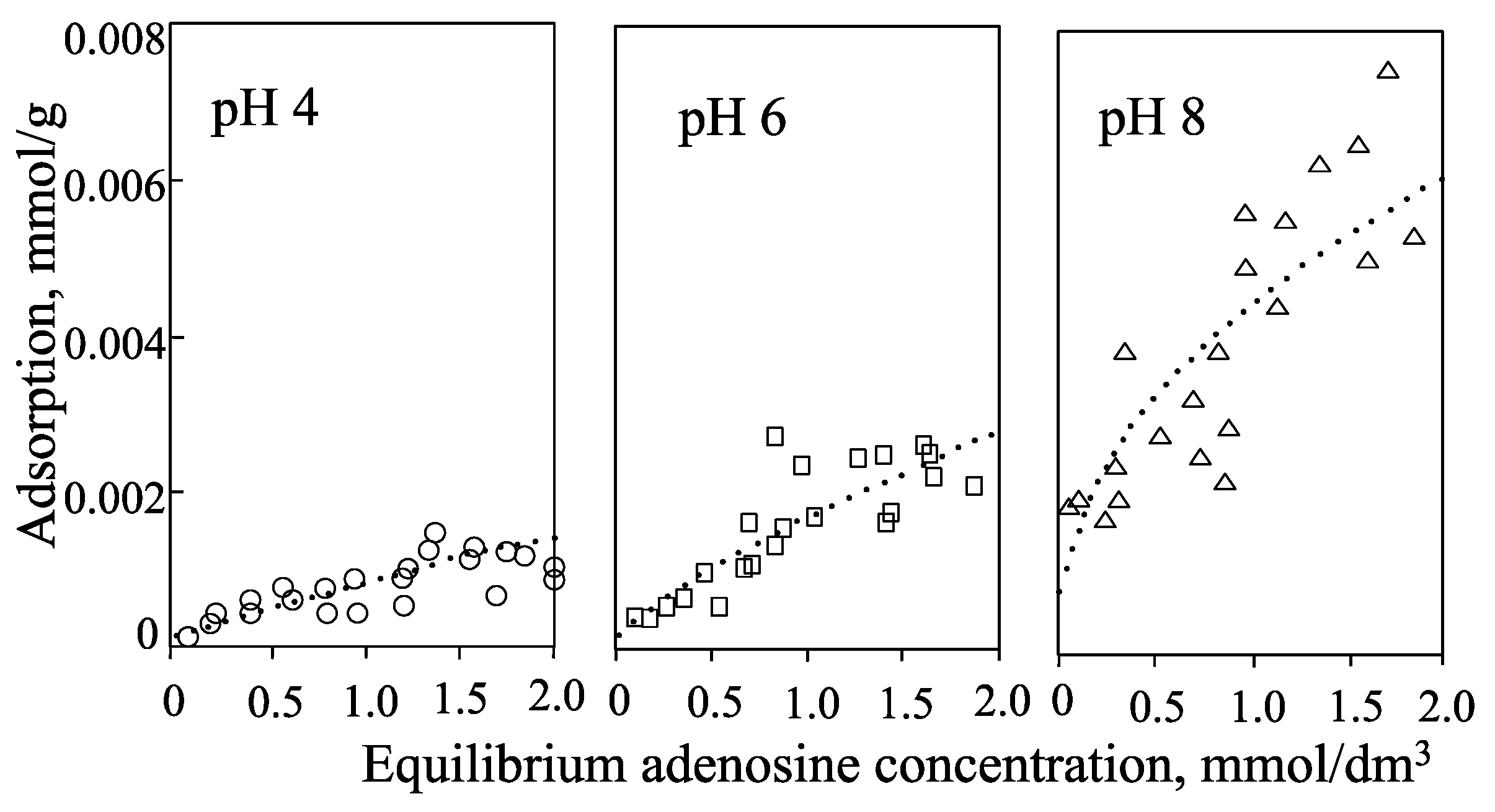
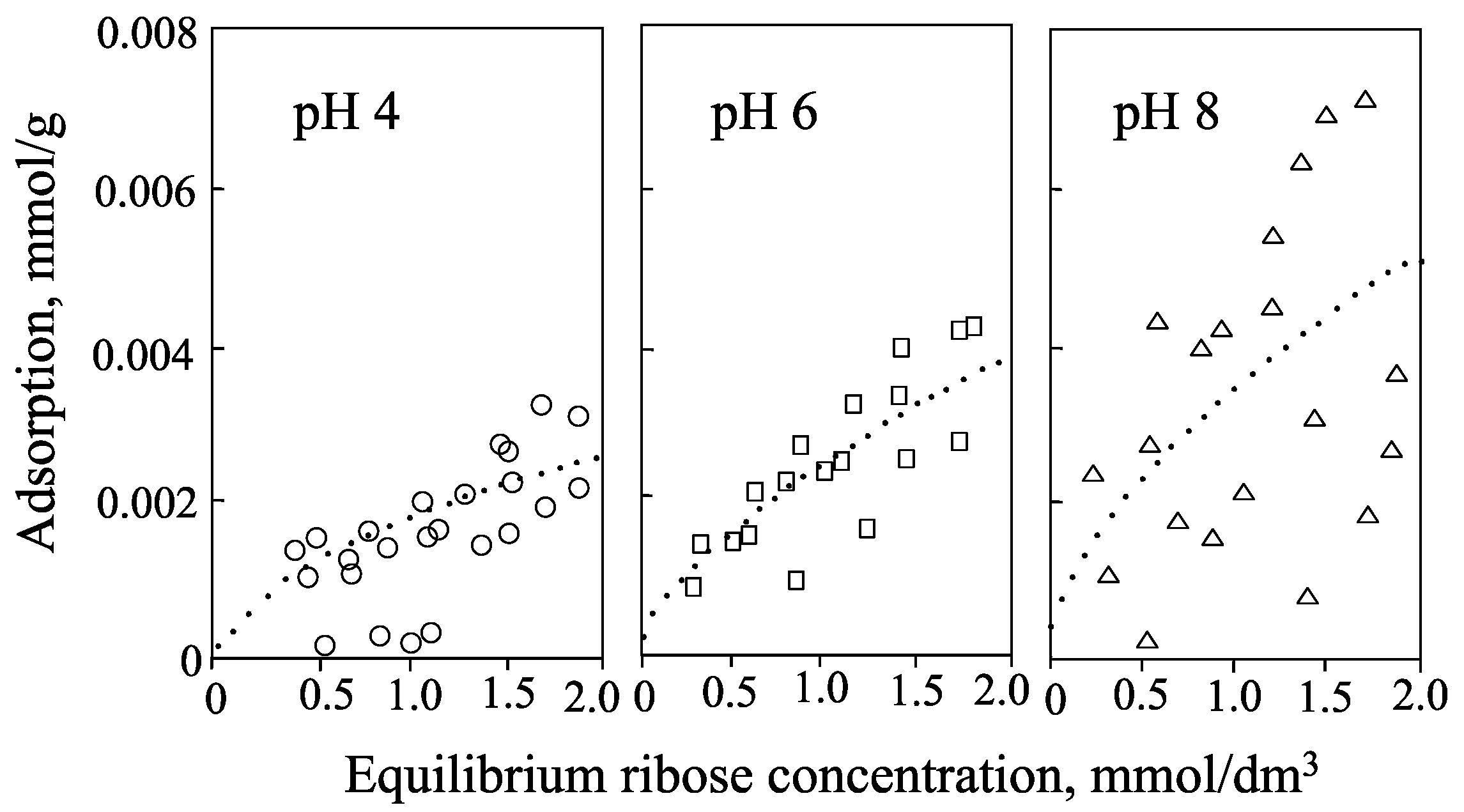
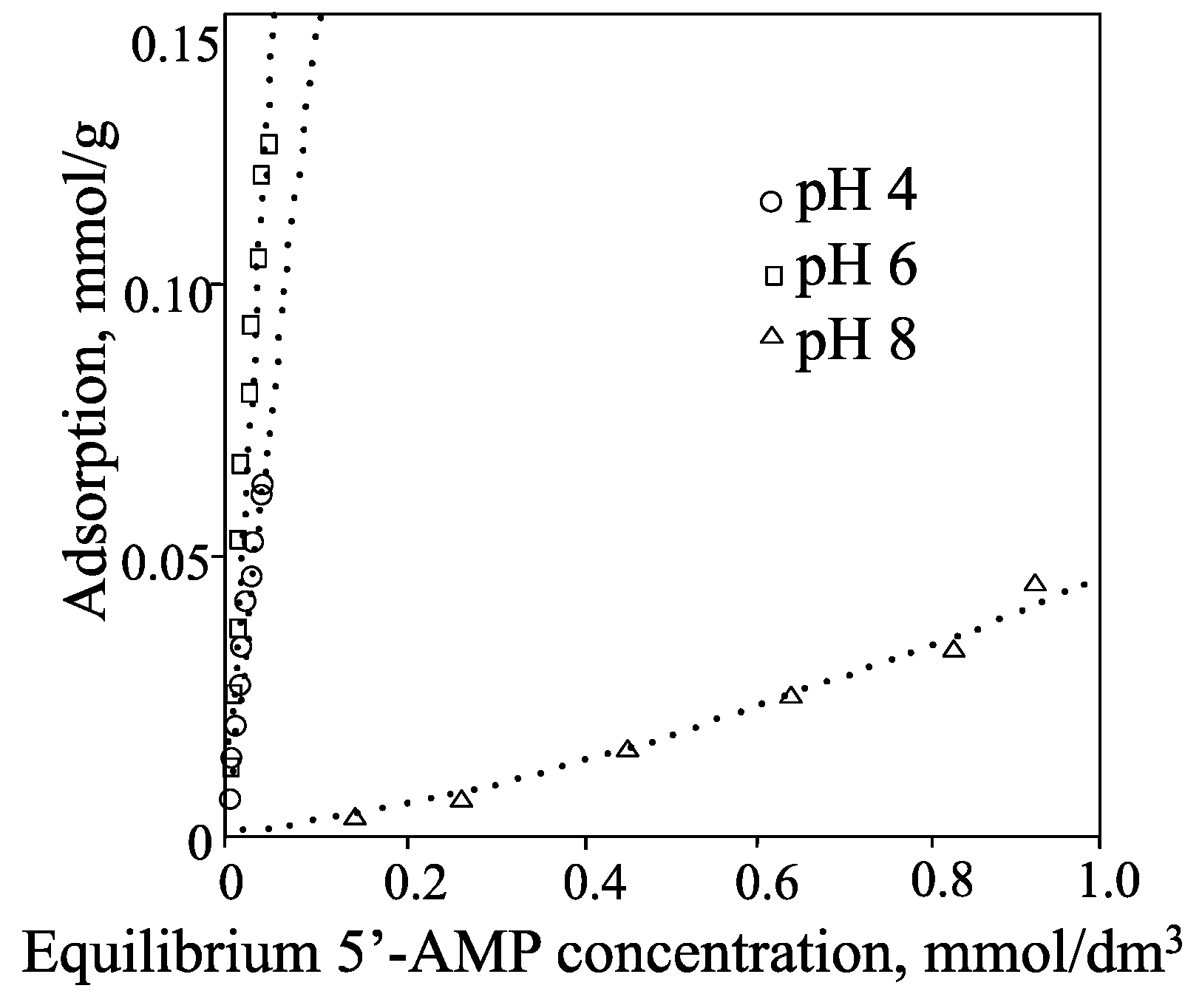
2. Smectite


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adenine | Cytosine | Uracil | Adenosine | Ribose | 5'-AMP | ||
|---|---|---|---|---|---|---|---|
| Dissociation constant | pKa1 | 4.15 * | 4.58 + | 9.48 + | 3.5 * | 3.80 # | |
| pKa2 | 9.8 * | 12.15 + | 12.5 * | 6.19 # | |||
| pKa3 | 13.06 # | ||||||
| Solubility (T) (g/100g) | 0.12 (25 °C) † | 0.743 (?)· | 0.36 (25 °C) † | 0.7 (?) § | 10 (?) †† | 97 (25 °C) ^ | |
| Molecular weight | 135 | 111 | 112 | 267 | 150 | 347 |

3. Kaoline Minerals
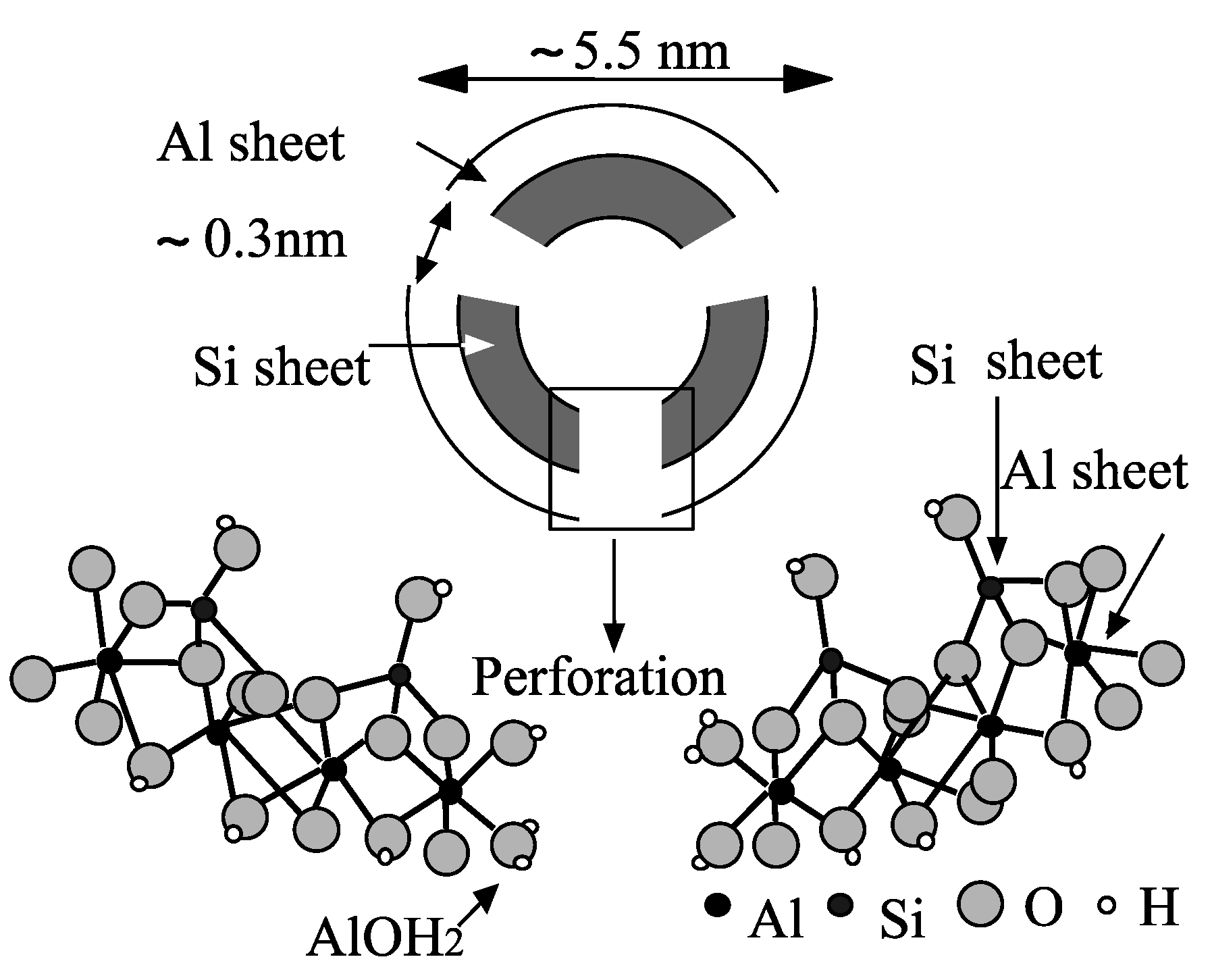
4. Allophane and Other Silicates


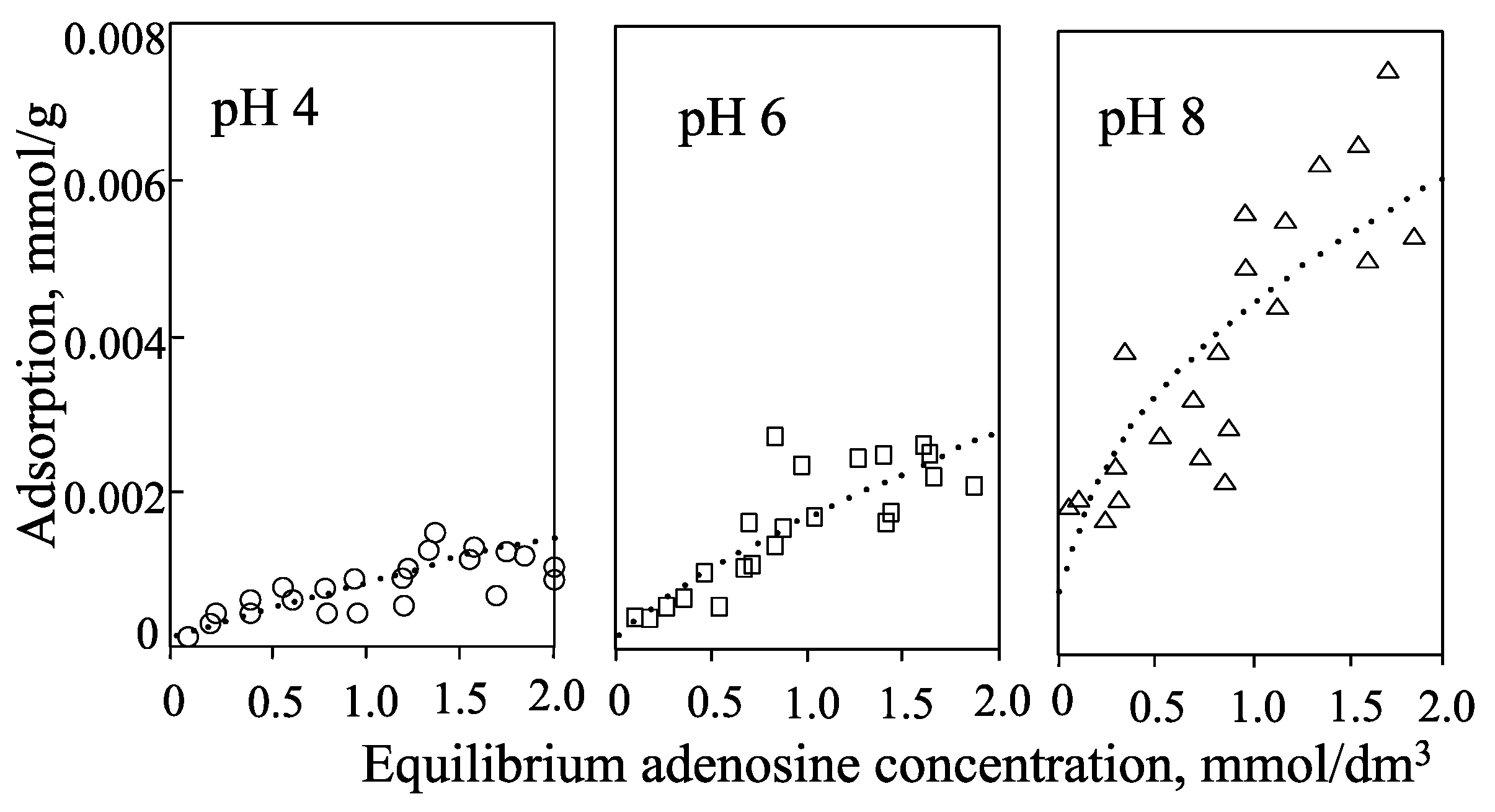


| pH 4 | pH 6 | pH 8 | |
|---|---|---|---|
| adenine | ++ | (+) N | N (+) |
| adenosine | ++ | (+) N | N |
| 5'-AMP | +− | − | −− |
| allophane | ++ | N | −− |
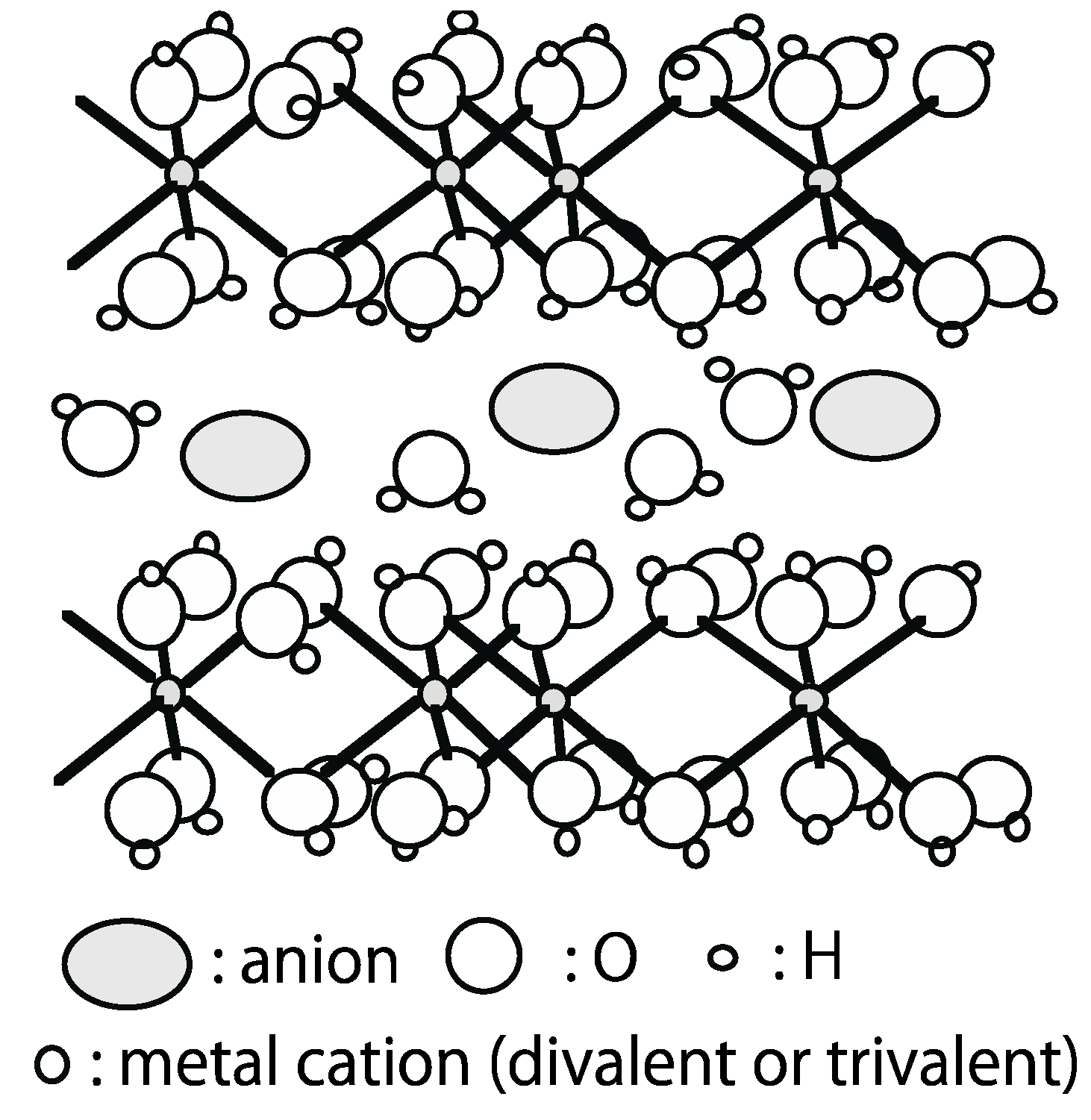
5. Layered Double Hydroxide (LDH)

| Mineral name | Cation | Anion | ||
|---|---|---|---|---|
| Rhombohedral | Hexagonal | M2+ | M3+ | |
| Hydrotalcite | Maasseite | Mg | Al | CO32− |
| Motukoreaite | Mg | Al | SO42−, CO32− | |
| Stichtite | Barbertonite | Mg | Cr | CO32− |
| Pyroaurite | Sjögrenite | Mg | Fe | CO32− |
| Iowaite | Mg | Fe | Cl− | |
| Chlormagaluminite | Mg | Fe | Cl−, CO32− | |
| Hydrocalmite | Ca | Al | OH− | |
| Green Rust 1 | Fe | Fe | CO32− | |
| Berthierine | Fe | Fe | SiO44− | |
| Takovite | Ni | Al | CO32− | |
| Reevesite | Ni | Fe | CO32− | |
| Honessite | Ni | Fe | SO42− | |
| Eardlyite | Zn, Ni | Al | CO32− | |
| Meixnerite | Mg | Al | OH− | |
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Theng, B.K.G. The Chemistry of Clay-Organic Reactions; Adam Hilger: London, UK, 1974; p. 343. [Google Scholar]
- Bernal, J.D. The Physical Basis of Life; Routledge and Kegan Paul: London, UK, 1951; p. 364. [Google Scholar]
- Ponnamperuma, C.; Shimoyama, A.; Friebele, E. Clay and the origin of life. Orig. Life 1982, 12, 9–49. [Google Scholar] [CrossRef] [PubMed]
- The Clay Mineral Society of Japan (Ed.) Handbook of Clays and Clay Minerals, 3rd ed.; Gihodoshuppan: Tokyo, Japan, 2009; pp. 11–17. (In Japanese)
- Cairns-Smith, A.G. Genetic Takeover and the Mineral Origins of Life; Cambridge University Press: Cambridge, UK, 1982; p. 133. [Google Scholar]
- Brack, A. Clay minerals and the origin of life. In Handbook of Clay Science; Bergaya, F., Theng, B.K.G., Lagaly, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 379–391. [Google Scholar]
- Cech, T.R. A model for the RNA-catalyzed replication of RNA. Proc. Nat. Acad. Sci. USA 1986, 83, 4360–4363. [Google Scholar] [CrossRef] [PubMed]
- Guerrier-Takada, C.; Altman, S. Catalytic activity of an RNA molecule prepared by transcription in vitro. Science 1984, 223, 285–286. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P.; Hill, A.R., Jr.; Liu, R.; Orgel, L.E. Synthesis of long prebiotic oligomers on mineral surfaces. Nature 1996, 381, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P. Mineral catalysis and prebiotic synthesis: Montmorillonite-Catalyzed formation of RNA. Elements 2005, 1, 145–149. [Google Scholar] [CrossRef]
- Ferris, J.P. Montmorillonite catalysis of 30−50 mer oligonucleotides: Laboratory demonstration of potential steps in the origin of the RNA world. Orig. Life Evol. Biosph. 2002, 32, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Lailach, G.E.; Thompson, T.D.; Brindley, G.W. Adsorption of pyrimidines, purines, and nucleosides by Li-, Na-, Mg-, and Ca-montmorillonite (Clay-organic studies XII). Clays Clay Miner. 1968, 16, 285–293. [Google Scholar] [CrossRef]
- Lailach, G.E.; Thompson, T.D.; Brindley, G.W. Absorption of pyrimidines, purines, and nucleosides by Co-, Ni-, Cu-, and Fe(III)- montmorillonite (Clay-organic studies XIII). Clays Clay Miner. 1968, 16, 295–301. [Google Scholar] [CrossRef]
- Benetoli, L.O.B.; de Santana, H.; Zaia, C.T.B.V.; Zaia, D.A.M. Adsorption of nucleic acid bases on clays: An investigation using Langmuir and Freundlich isotherms and FT-IR spectroscopy. Monatsh. Chem. 2008, 139, 753–761. [Google Scholar] [CrossRef]
- Pecini, E.M.; Avena, M.J. Measureing the isoelectric point of the edges of clay mineral particles: The case of montmorillonite. Langmuir 2013, 29, 14926–14934. [Google Scholar] [CrossRef] [PubMed]
- Perezgasga, L.; Serrato-Díaz, A.; Negron-Mendóza, A.; Galán, L.D.; Mosqeira, F.G. Sites of adsorption of adenine, uracil and their corresponding derivatives on sodium montmorillonite. Orig. Life Evol. Biosph. 2005, 35, 91–110. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.; Zubay, G. Binding of adenine and adenosine-related compounds to the clay montmorillonite and the mineral hydrocyapatite. Orig. Life Envol. Biosph. 1995, 25, 61–81. [Google Scholar] [CrossRef]
- Hashizume, H.; van der Gaast, S.; Theng, B.K.G. Adsorption of adenine, cytosine, uracil, ribose, and phosphate by Mg-exchanged montmorillonite. Clay Miner. 2010, 45, 469–475. [Google Scholar] [CrossRef]
- The chemical Society of Japan (Ed.) Kagakubinran; Maruzen: Tokyo, Japan, 1993; pp. II317–II321.
- Banin, A.; Lawless, J.G.; Church, F.M.; Margulies, L.; Orenberg, J.B. pH profile of the adsorption of nucleotides onto montmorillonite. II. Adsorption and desorption of 5'-AMP in iron-calcium montmorillonite systems. Orig. Life 1985, 15, 89–101. [Google Scholar] [CrossRef]
- The chemical Society of Japan. Kagakubinran; Maruzen: Tokyo, Japan, 1993; pp. II176–II178. [Google Scholar]
- Pub Chem. Available online: http://our.edu/cccda/sl/solubility/allsolvent (accessed on 17 December 2014).
- Product Information. Available online: http://www.sigmaaldrich.com/ (accessed on 17 December 2014).
- Lookchem. Available online: http://www.lookchem.com (accessed on 17 December 2014).
- Wang, D.; Li, Z.-J.; Ying, H.-J. Solubility of adenosine 5'-monophosphate in different solvents from (288.15 to 330.15)K. J. Chem. Eng. Data 2010, 55, 992–993. [Google Scholar] [CrossRef]
- Giles, C.H.; MacEwan, T.H.; Nakhwa, S.N.; Smith, D. Studies in adsorption. XI. A system of classification of solution adsorption isotherms and its use in diagnosis of adsorption mechanisms and in measurement of specific surface areas of solids. J. Chem. Soc. 1960, 3973–3993. [Google Scholar]
- Durán, J.D.G.; Ramos-Tejada, M.; Arroyo, F.J.; González-Caballero, F. Rheological and Electrokinetic properties of sodium montmorillonite suspensions. J. Colloid Interface Sci. 2000, 229, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Lawless, L.G.; Church, F.M.; Mazzurco, J.; Huff, R.; Kao, J.; Cook, A.; Lowe, T.; Orenberg, J.B.; Edelason, E. pH profiles of the adsorption of nucleotides onto montmorillontie. I. Selected homoionic clays. Orig. Life 1985, 15, 77–88. [Google Scholar] [CrossRef]
- Feuillie, C.; Daniel, I.; Michot, L.J.; Pedreira-Segada, U. Adsorption of nucleotides onto Fe-Mg-Al rich swelling clays. Geochim. Cosmoshim. Acta 2013, 120, 97–108. [Google Scholar] [CrossRef]
- Franchi, M.; Ferris, J.P.; Gallori, E. Cations as mediators of the adsorption of nucleic acids on clay surfaces in prebiotic environments. Orig. Life Evol. Biosph. 2003, 33, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Biondi, E.; Branciamore, S.; Maurel, M.-C.; Gallori, E. Motmorillonite protection of an UV-irradiated hairpin ribozyme: Evolution of the RNA world in a mineral environment. BMC Evol. Biol. 2007, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Ovando, E.; Negron-Medoza, A. Radiation chemistry approach to the study of sedimentary microenvironments as models for the protection of bio-organic molecules on the early earth. J. Radiational Nucleic Chem. 2010, 286, 637–642. [Google Scholar] [CrossRef]
- Mignon, P.; Ugleingo, P.; Sodupe, M. Theoretical study of the adsorption of RNA/DNA bases on the external surfaces of Na+-montorillonite. J. Phys. Chem. C 2009, 113, 13741–13749. [Google Scholar] [CrossRef]
- Mathew, D.C.; Luthey-Schulten, Z. Influence of montmorillonte on nucleotide oligomerization reactions: A molecular dyamics study. Orig. Life Evol. Biosph. 2010, 40, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Swadling, J.B.; Coveney, P.V.; Greenwell, H.C. Clay minerals mediate folding and regioselective interactions of RNA: A large-scale atmistic simulation study. J. Am. Chem. Soc. 2010, 132, 13750–13764. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.C.; Aldersley, M.F.; Delano, J.W.; Ferris, J.P. Mechanism of montmroillonite catalysis in the formation of RNA oligomers. J. Am. Chem. Soc. 2009, 131, 13369–13374. [Google Scholar] [CrossRef] [PubMed]
- Ertem, G.; Ferris, J.P. Sequence- and regio-selectivity in the montmorillonite-catalyzed systems of RNA. Orig. Life Evol. Biosph. 2000, 30, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Ertem, G.; Ferris, J.P. Synthesis of RNA ligomers on heterogenous templates. Nature 1996, 379, 238–240. [Google Scholar] [CrossRef] [PubMed]
- The clay mineral society of Japan. Available online: http://www.cssj2.org/seminar1/section04/ext.html (accessed on 16 January 2015). (In Japanese)
- Graf, G.; Lagaly, G. Interaction of clay minerals with adenosine -5-phosphates. Clays Clay Miner. 1980, 20, 12–18. [Google Scholar] [CrossRef]
- Robinson, T.L.; Michalkova, A.; Gorb, L.; Leszczynaski, J. Hydrogen bonding of thymine and uracil with surface of dickite: An ab intio study. J. Mol. Struc. 2007, 844–845, 48–58. [Google Scholar] [CrossRef]
- Michalkove, A.; Robinson, T.L.; Leszczynski, J. Adsorption of thymine and uracil on 1:1 clay mineral surfaces: Comprehensive ab initio study on influence of sodium cation and water. Phys. Chem. Chem. Phys. 2011, 13, 7862–7881. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, H.; Theng, B.K.G. Adsorption of dl-alanine by allophane: Effect of pH and unit particle aggregation. Clay Miner. 1999, 34, 233–238. [Google Scholar] [CrossRef]
- Hashizume, H.; Theng, B.K.G. Adenine, adenosine, ribose and 5'-AMP adsorption to allophane. Clays Clay Miner. 2007, 55, 599–605. [Google Scholar] [CrossRef]
- Theng, B.K.G.; Russell, M.; Churchman, G.J.; Parfitt, R.L. Surface properties of allophane, halloysite, and imogolite. Clays Clay Miner. 1982, 30, 143–149. [Google Scholar] [CrossRef]
- Rajan, S.S.S. Mechanism of phosphate adsorption by allophane clays. N. Zeal. J. Sci. 1975, 18, 93–101. [Google Scholar]
- Matsuura, Y.; yoda, F.; Arakawa, S.; John, B.; Okamoto, M.; Hayashi, H. DNA adsorption characteristics of hollow spherule allophane nano-particles. Mater. Sci. Eng. C 2013, 33, 5079–5083. [Google Scholar] [CrossRef]
- Aisawa, S.; Hirahara, H.; Ishiyama, K.; Ogasawara, W.; Umetsu, Y.; Narita, E. Sugar-anionic clay composite materials: Interaction of pentoses in layered double hydroxide. J. Solid State Chem. 2003, 174, 342–348. [Google Scholar] [CrossRef]
- Swadling, J.B.; Suter, J.L.; Christopher Greenwell, H.; Coveney, P.V. Infulence of surface chemistry on mineral-RNA inteactions. Langmuir 2013, 29, 1573–1583. [Google Scholar] [CrossRef] [PubMed]
- Swadling, J.B.; Coveney, P.V.; Christopher Greenwell, H. Stability of free and minerals-protected nucleic acids: Implications for the RNA world. Geochim. Cosmochim. Acta 2013, 83, 360–378. [Google Scholar] [CrossRef]
- Saladino, R.; Neri, V.; Crestini, C. Role of clays in the prebiotic synthesis of sugar derivatives from formamide. Philos. Mag. 2010, 90, 2329–2337. [Google Scholar] [CrossRef] [Green Version]
- Pitsch, S.; Eschenmoser, A.; Gedulin, B.; Hui, S.; Arrhenius, G. Mineral induced formation of sugar phosphates. Orig. Life Evol. Biosph. 1995, 25, 297–334. [Google Scholar] [CrossRef] [PubMed]
- The Clay Mineral Society of Japan (Ed.) Handbook of Clays and Clay Minerals, 3rd ed.; Gihodoshuppan: Tokyo, Japan, 2009; pp. 99–103. (In Japanese)
- Miller, S.L. A production of amino acids under possible primitive Earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L.; Urey, H.C. Organic compound synthesis on the primitive Earth. Science 1959, 130, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.T.; Cleaves, H.J.; Dwoekin, J.P.; Glavin, D.P.; Callahan, M.; Aubrey, A.; Lazcano, A.; Bada, J.L. Primordial synthesis of amines and amino acids in a 1958 Miller H2S-rich spark discharge experiment. Proc. Natl. Acad. Sci. USA 2011, 108, 5526–5531. [Google Scholar] [CrossRef]
- Powner, M.W.; Gerland, B.; Stherland, J.D. Syhthesis of activated pyrimidine ribonucleotides in prebiotically plausible coditions. Nature 2009, 459, 239–242. [Google Scholar]
- Ferris, J.P.; Erten, G.; Agarwel, V. Mineral catalysis of the formation of dimmers of 5'-AMP in aqueous soltion: The possible role of montmorillonite clays I the prebiotic synthesis of RNA. Orig. Life Evol. Biosph. 1989, 19, 165–178. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hashizume, H. Adsorption of Nucleic Acid Bases, Ribose, and Phosphate by Some Clay Minerals. Life 2015, 5, 637-650. https://doi.org/10.3390/life5010637
Hashizume H. Adsorption of Nucleic Acid Bases, Ribose, and Phosphate by Some Clay Minerals. Life. 2015; 5(1):637-650. https://doi.org/10.3390/life5010637
Chicago/Turabian StyleHashizume, Hideo. 2015. "Adsorption of Nucleic Acid Bases, Ribose, and Phosphate by Some Clay Minerals" Life 5, no. 1: 637-650. https://doi.org/10.3390/life5010637
APA StyleHashizume, H. (2015). Adsorption of Nucleic Acid Bases, Ribose, and Phosphate by Some Clay Minerals. Life, 5(1), 637-650. https://doi.org/10.3390/life5010637