
Arsenite as an Electron Donor for Anoxygenic Photosynthesis: Description of Three Strains of Ectothiorhodospira from Mono Lake, California and Big Soda Lake, Nevada


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Growth
2.2. Sequencing and Phylogenetic Analysis
2.3. Detection of arxA and aioA
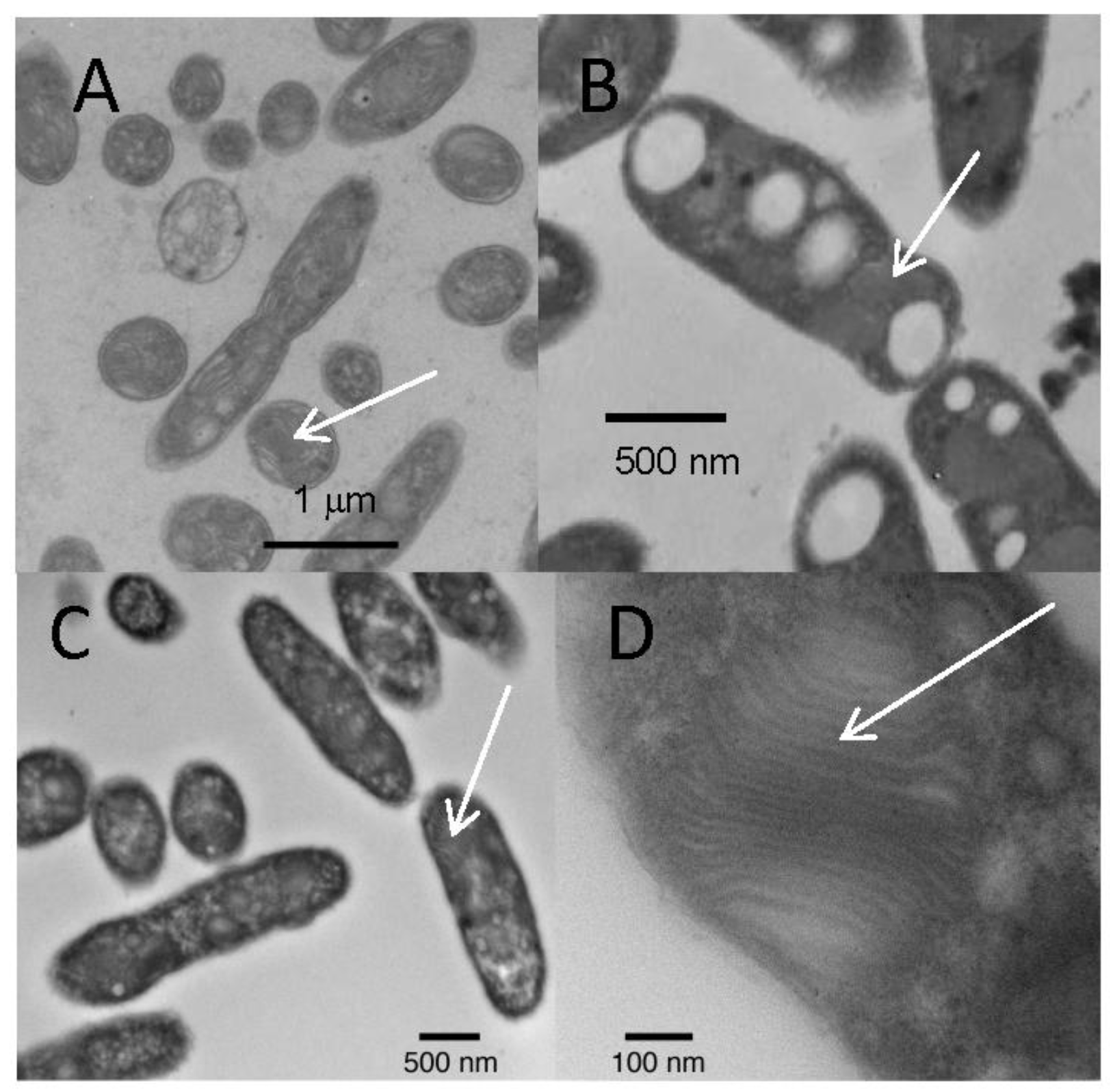
2.4. Electron Microscopy
2.5. Analytical Methods
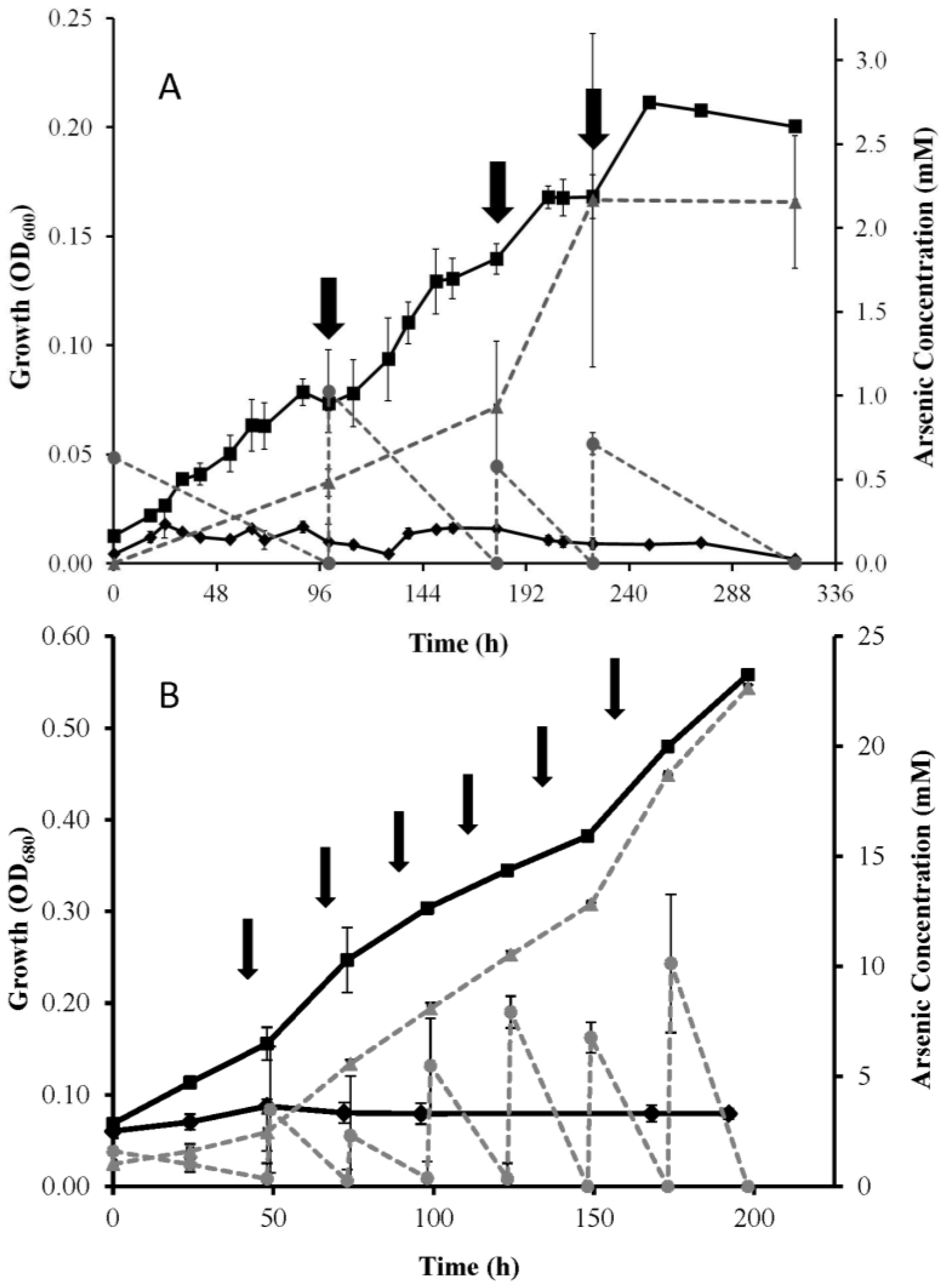
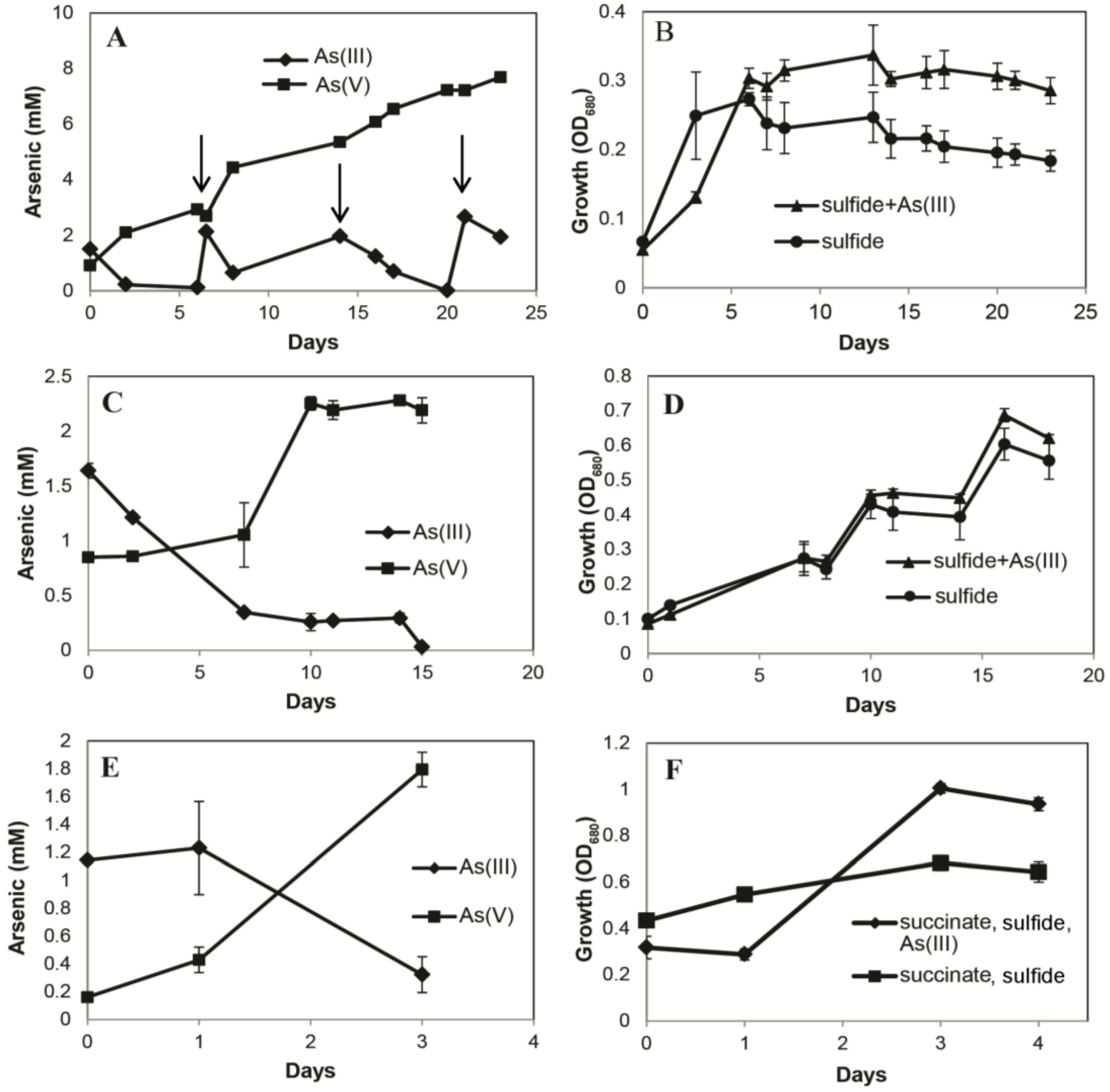
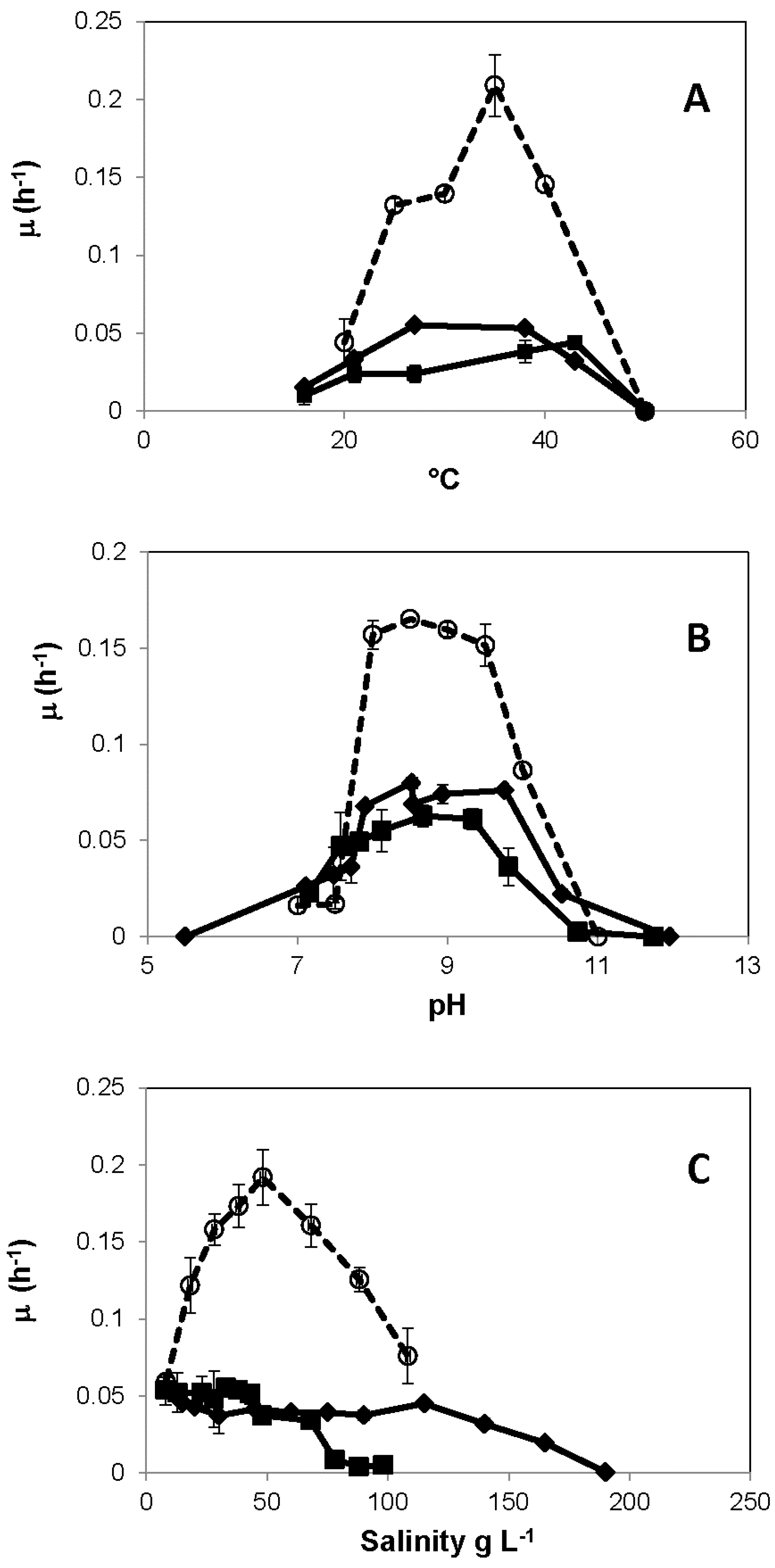
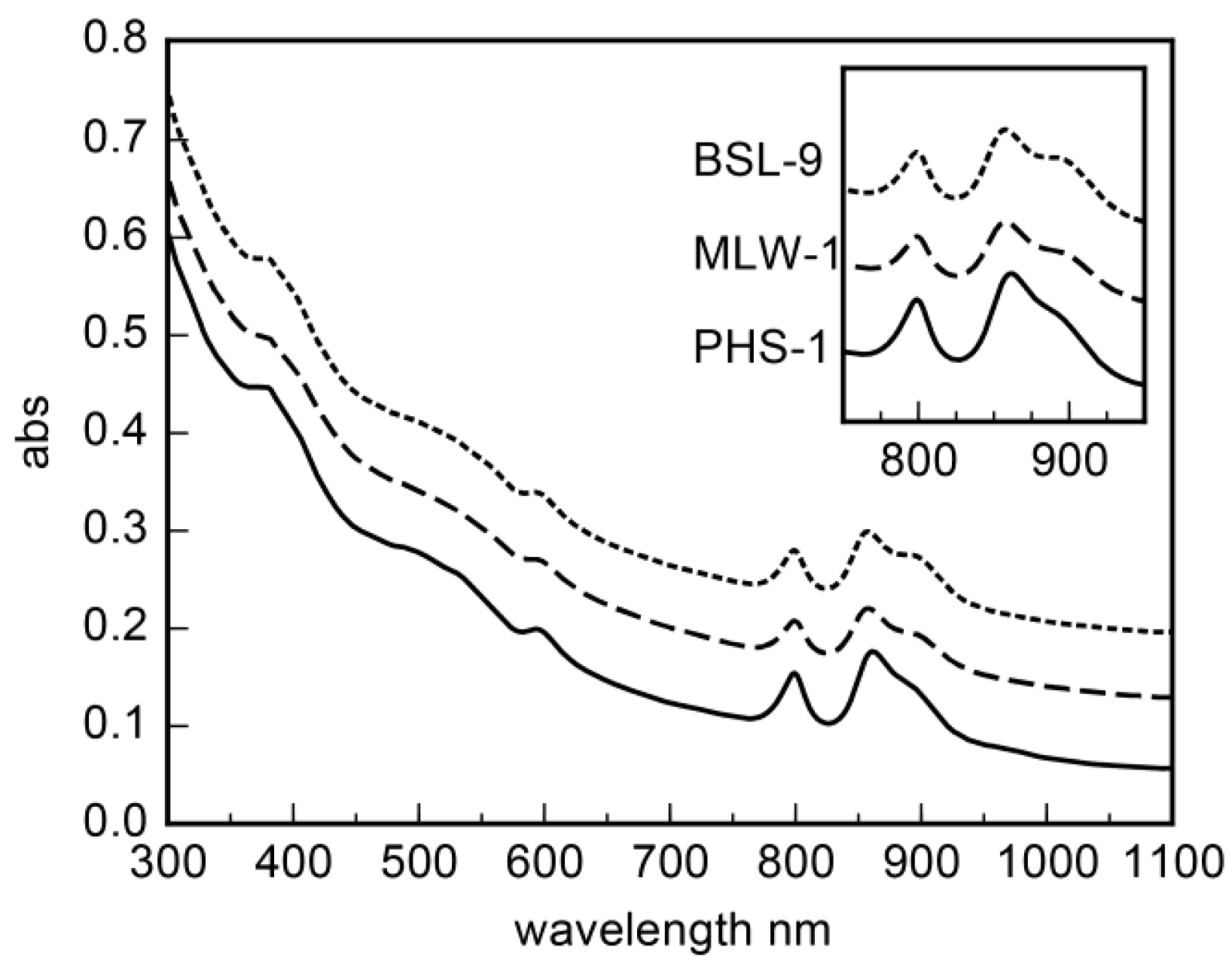
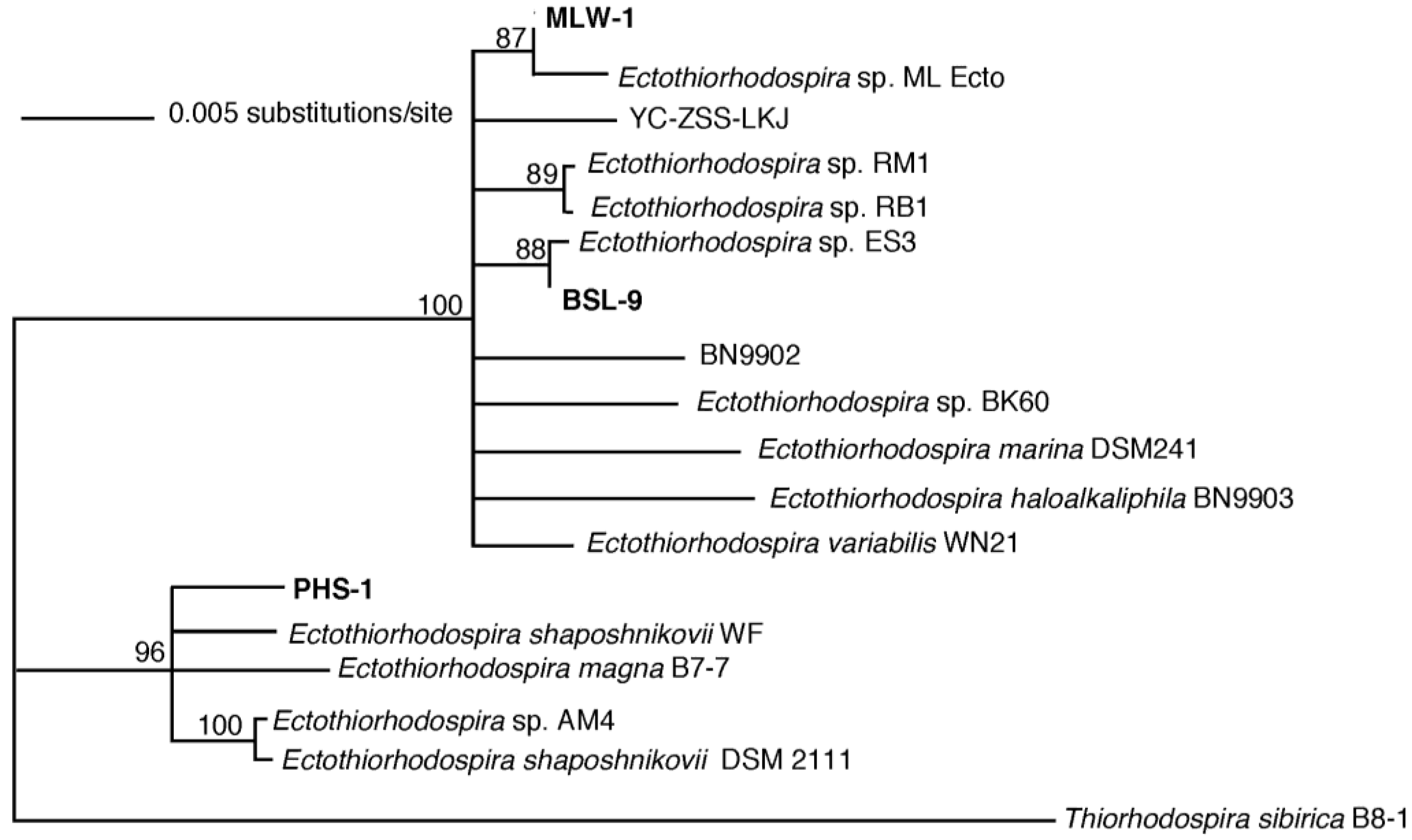
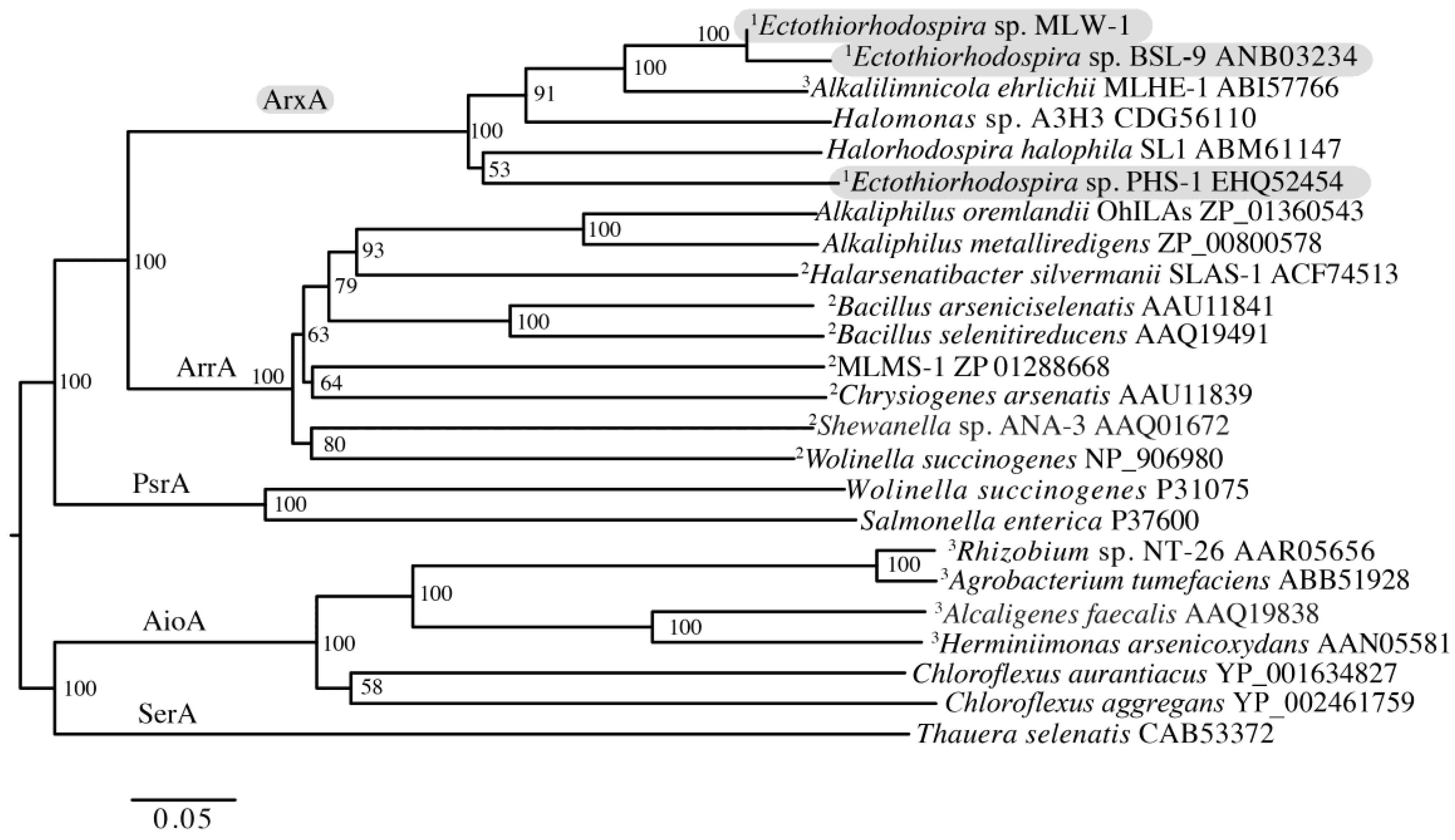
3. Results
3.1. Growth Experiments
3.2. Morphological Characteristics of the Isolates
3.3. Phylogenetic Analysis
3.4. Analysis of the Anaerobic Arsenite Oxidase Gene arxA
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Imhoff, J.F.; Süling, J. The phylogenetic relationship among Ectothiorhodospiraceae: A re-evalution of their taxonomy on the basis of 16S rDNA analyses. Arch. Microbiol. 1996, 165, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F. The Family Ectothiorhodospiraceae. In The Prokaryotes: A Handbook on the Biology of Bacteria, 3rd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 6, pp. 874–886. [Google Scholar]
- Budinoff, C.R.; Hollibaugh, J.T. Arsenite-dependent photoautotrophy by an Ectothiorhodospira-dominated consortium. ISME J. 2008, 2, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Kulp, T.R.; Hoeft, S.E.; Asao, M.; Madigan, M.T.; Hollibaugh, J.T.; Fisher, J.C.; Stolz, J.F.; Culbertson, C.W.; Miller, L.G.; Oremland, R.S. Arsenic (III) fuels anoxygenic photosynthesis in hot spring biofilms from Mono Lake, California. Science 2008, 321, 967–970. [Google Scholar] [CrossRef] [PubMed]
- Oremland, R.S. A random biogeochemical walk into three soda lakes of the western USA: With an introduction to a few of their microbial denizens. In Polyextremophiles, Life under Multiple Forms of Stress; Seckbach, J., Oren, A., Stan-Lotter, H., Eds.; Springer: New York, NY, USA, 2013; pp. 180–199. [Google Scholar]
- Oremland, R.S.; Stolz, J.F.; Hollibaugh, J.T. The microbial arsenic cycle in Mono Lake, California. FEMS Microbiol. Ecol. 2004, 48, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Oremland, R.S.; Dowdle, P.R.; Hoeft, S.; Sharp, J.O.; Schaefer, J.K.; Miller, L.G.; Switzer Blum, J.; Smith, R.L.; Bloom, N.S.; Wallschlaeger, D. Bacterial dissimilatory reduction of arsenate and sulfate in meromictic Mono Lake, California. Geochim. Cosmochim. Acta 2000, 64, 3073–3084. [Google Scholar] [CrossRef]
- Hoeft, S.E.; Kulp, T.R.; Stolz, J.F.; Hollibaugh, J.T.; Oremland, R.S. Dissimilatory arsenate reduction with sulfide as electron donor: Experiments with Mono Lake water and isolation of strain MLMS-1, a chemoautotrophic arsenate respirer. Appl. Environ. Microbiol. 2004, 70, 2741–2747. [Google Scholar] [CrossRef] [PubMed]
- Hoeft, S.E.; Lucas, F.; Hollibaugh, J.T.; Oremland, R.S. Characterization of bacterial arsenate reduction in the anoxic bottom waters of Mono Lake, California. Geomicrobiology 2002, 19, 1–19. [Google Scholar]
- Hoeft, S.E.; Blum, J.S.; Stolz, J.F.; Tabita, F.R.; Witte, B.; King, G.M.; Santini, J.M.; Oremland, R.S. Alkalilimnicola ehrlichii sp. nov., a novel, arsenite-oxidizing haloalkaliphilic gammaproteobacterium capable of chemoautotrophic or heterotrophic growth with nitrate or oxygen as the electron acceptor. Int. J. Syst. Evol. Microbiol. 2007, 57, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Richey, C.; Chovanec, P.; Hoeft, S.E.; Oremland, R.S.; Basu, P.; Stolz, J.F. Respiratory arsenate reductase as a bidirectional enzyme. Biochem. Biophys. Res. Commun. 2009, 382, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Zargar, K.; Hoeft, S.E.; Oremland, R.S.; Saltikov, C.S. Identification of a novel arsenite oxidase gene, arxA, in the haloalkaliphilic, arsenite-oxidizing bacterium Alkalilimnicola ehrlichii strain MLHE-1. J. Bacteriol. 2010, 192, 3755–3762. [Google Scholar] [CrossRef] [PubMed]
- Hoeft, S.E.; Kulp, T.R.; Han, S.; Lanoil, B.; Oremland, R.S. Coupled arsenotrophy in a hot spring photosynthetic biofilm at Mono Lake, California. Appl. Environ. Microbiol. 2010, 76, 4633–4639. [Google Scholar] [CrossRef] [PubMed]
- Zargar, K.; Conrad, A.; Bernick, D.L.; Lowe, T.M.; Stolc, V.; Hoeft, S.E.; Oremland, R.S.; Stolz, J.F.; Saltikov, C.W. ArxA, a new clade of arsenite oxidase within the DMSO reductase family of molybdenum oxidoreductases. Environ. Microbiol. 2012, 14, 1635–1645. [Google Scholar] [CrossRef] [PubMed]
- Hamamura, N.; Itai, T.; Liu, Y.; Reysenbach, A.-L.; Damdinsuren, N.; Inskeep, W.P. Identification of anaerobic arsenite-oxidizing and arsenate-reducing bacteria associated with an alkaline saline lake in Khovsgol, Mongolia. Environ. Microbiol. Rep. 2014, 6, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Widdel, F.; Koring, G.W.; Mayer, F. Studies on the dissimiliatory sulfate-reducing bacteria that decompose fatty acids. 3. Characterization of the filamentous gliding Desulfonema limicola, gen. nov. sp. nov., and Desulfonema magnum, sp. nov. Arch. Microbiol. 1983, 134, 286–294. [Google Scholar] [CrossRef]
- Oremland, R.S.; Blum, J.S.; Culbertson, C.W.; Visscher, P.T.; Miller, L.G.; Dowdle, P.; Strohmaier, F.E. Isolation, growth, and metabolism of an obligately anaerobic, selenite-respiring bacterium strain SES-3. Appl. Environ. Microbiol. 1994, 60, 3011–3019. [Google Scholar] [PubMed]
- Hermann, M.; Noll, K.M.; Wolfe, R.S. Improved agar bottle plate for isolation of methanogens or other anaerobes in a defined gas atmosphere. Appl. Environ. Microbiol. 1986, 51, 1124–1126. [Google Scholar] [PubMed]
- Widdel, F.; Pfennig, N. Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids. Arch. Microbiol. 1981, 129, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Kostka, J.; Nealson, K. Isolation, cultivation, and characterization of iron- and manganese-reducing bacteria. In Techniques in Microbial Ecology; Burlage, R.S., Atlas, R., Stahl, D., Geesey, G., Sayler, G., Eds.; Oxford University Press: Oxford, UK, 1998; pp. 58–78. [Google Scholar]
- Raymond, J.C.; Sistrom, W.R. Ectothiorhodospira halophila: A new species of the genus Ectothiorhodospira. Arch. Mikrobiol. 1969, 69, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Cashion, P.; Hodler-Franklin, M.A.; McCully, J.; Franklin, M. A rapid method for base ratio determination of bacterial DNA. Anal. Biochem. 1977, 81, 461–466. [Google Scholar] [CrossRef]
- De Ley, J.; Cattoir, H.; Reynaerts, A. The quantitative measurement of DNA hybridization from renaturation rates. Eur. J. Biochem. 1970, 12, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Huss, V.A.R.; Festl, H.; Schleifer, K.H. Studies on the spectrophotometric determination of DNA hybridization from renaturation rates. Syst. Appl. Microbiol. 1983, 4, 184–192. [Google Scholar] [CrossRef]
- Sultana, M.; Vogler, S.; Zargar, K.; Schmidt, A.C.; Saltikov, C.; Seifert, J.; Schlömann, M. New clusters of arsenite oxidase and unusual bacterial groups in enrichments from arsenic-contaminated soil. Arch. Microbiol. 2012, 194, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Maldonado, J.; Stoneburner, B.; Boren, A.; Miller, L.; Rosen, M.; Oremland, R.S.; Saltikov, C.W. Genome sequence of the photoarsenotrophic bacterium, Ectothiorhodospira str. sp. BSL-9, isolated from a saline alkaline arsenic rich extreme environment. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.L.; Strohmaier, F.S.; Oremland, R.S. Isolation of anaerobic oxalate degrading bacteria from freshwater lake sediments. Arch. Microbiol. 1985, 14, 8–13. [Google Scholar] [CrossRef]
- Switzer Blum, J.; Burns Bindi, A.; Buzzelli, J.; Stolz, J.; Oremland, R.S. Bacillus arsenicoselenatis sp. nov., and Bacillus selenitireducens sp. nov.: Two haloalkaliphiles from Mono Lake, California which respire oxyanions of selenium and arsenic. Arch. Microbiol. 1998, 171, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Cline, J.D. Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol. Oceanogr. 1969, 14, 454–459. [Google Scholar] [CrossRef]
- Stauder, S.; Raue, B.; Sacher, F. Thioarsenates in sulfidic waters. Environ. Sci. Technol. 2005, 39, 5933–5939. [Google Scholar] [CrossRef] [PubMed]
- Edwardson, C.F.; Planer-Friedrich, B.; Hollibaugh, J.T. Transformation of monothioarsenate by haloalkaliphilic, anoxygenic photosynthetic purple sulfur bacteria. FEMS Microbiol. Ecol. 2014, 90, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Maldonado, J.; Sanchez-Sedillo, B.; Stoneburner, B.; Boren, A.; Miller, L.G.; Hoeft McCann, S.; Rosen, M.; Oremland, R.S.; Saltikov, C.W. The genetic basis of anoxygenic photosynthetic arsenite oxidation. Environ. Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Andres, J.; Bertin, P.N. The microbial genomics of arsenic. FEMS Microbiol. Rev. 2016, 40, 299–322. [Google Scholar] [CrossRef] [PubMed]
- Edwardson, C.F. Microbially Mediated Transformation of Arsenic and Sulfur Compounds in Mono Lake, California, USA. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2015. [Google Scholar]
- Edwardson, C.F.; Hollibaugh, J.T. Metatranscriptomic analysis of the prokaryotic community active in sulfur and arsenic cycling in alkaline, hypersaline Mono Lake, California. ISME J. 2016. in revision. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MLW-1 | PHS-1 | BSL-9 | Ect. shaposhnikovii (DSM 2111) | Ect. shaposhnikovii (DSM 243) Type Strain | Halorhodospira halophila | |
|---|---|---|---|---|---|---|
| Arsenite (2 mM) | + | + | + | - | - | - |
| Sulfide | + | + | + | + | + | + b |
| Thiosulfate | + | + | + | + | + a | + b |
| Sulfur | + | + | + | + | + a | + b |
| Hydrogen | - | - | ND | - | + a | ND |
| Acetate | + | + | + | + | + | + b |
| Lactate | + | + | + | + | + | ND |
| Propionate | + | + | + | + | + a | ND |
| Fructose | + | - | ND | + | + a | ND |
| Glucose | + | - | ND | - | ND | ND |
| Succinate | + | + | + | + | + a | + b |
| Malate | + | + | + | + | + a | ND |
| Pyruvate | + | + | + | + | + a | ND |
| Butyrate | - | - | ND | + | - | ND |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoeft McCann, S.; Boren, A.; Hernandez-Maldonado, J.; Stoneburner, B.; Saltikov, C.W.; Stolz, J.F.; Oremland, R.S. Arsenite as an Electron Donor for Anoxygenic Photosynthesis: Description of Three Strains of Ectothiorhodospira from Mono Lake, California and Big Soda Lake, Nevada. Life 2017, 7, 1. https://doi.org/10.3390/life7010001
Hoeft McCann S, Boren A, Hernandez-Maldonado J, Stoneburner B, Saltikov CW, Stolz JF, Oremland RS. Arsenite as an Electron Donor for Anoxygenic Photosynthesis: Description of Three Strains of Ectothiorhodospira from Mono Lake, California and Big Soda Lake, Nevada. Life. 2017; 7(1):1. https://doi.org/10.3390/life7010001
Chicago/Turabian StyleHoeft McCann, Shelley, Alison Boren, Jaime Hernandez-Maldonado, Brendon Stoneburner, Chad W. Saltikov, John F. Stolz, and Ronald S. Oremland. 2017. "Arsenite as an Electron Donor for Anoxygenic Photosynthesis: Description of Three Strains of Ectothiorhodospira from Mono Lake, California and Big Soda Lake, Nevada" Life 7, no. 1: 1. https://doi.org/10.3390/life7010001
APA StyleHoeft McCann, S., Boren, A., Hernandez-Maldonado, J., Stoneburner, B., Saltikov, C. W., Stolz, J. F., & Oremland, R. S. (2017). Arsenite as an Electron Donor for Anoxygenic Photosynthesis: Description of Three Strains of Ectothiorhodospira from Mono Lake, California and Big Soda Lake, Nevada. Life, 7(1), 1. https://doi.org/10.3390/life7010001