
SARS-CoV-2-Laden Respiratory Aerosol Deposition in the Lung Alveolar-Interstitial Region Is a Potential Risk Factor for Severe Disease: A Modeling Study


Abstract
1. Introduction
2. Materials and Methods
2.1. Respiratory Aerosol Particle Modeling
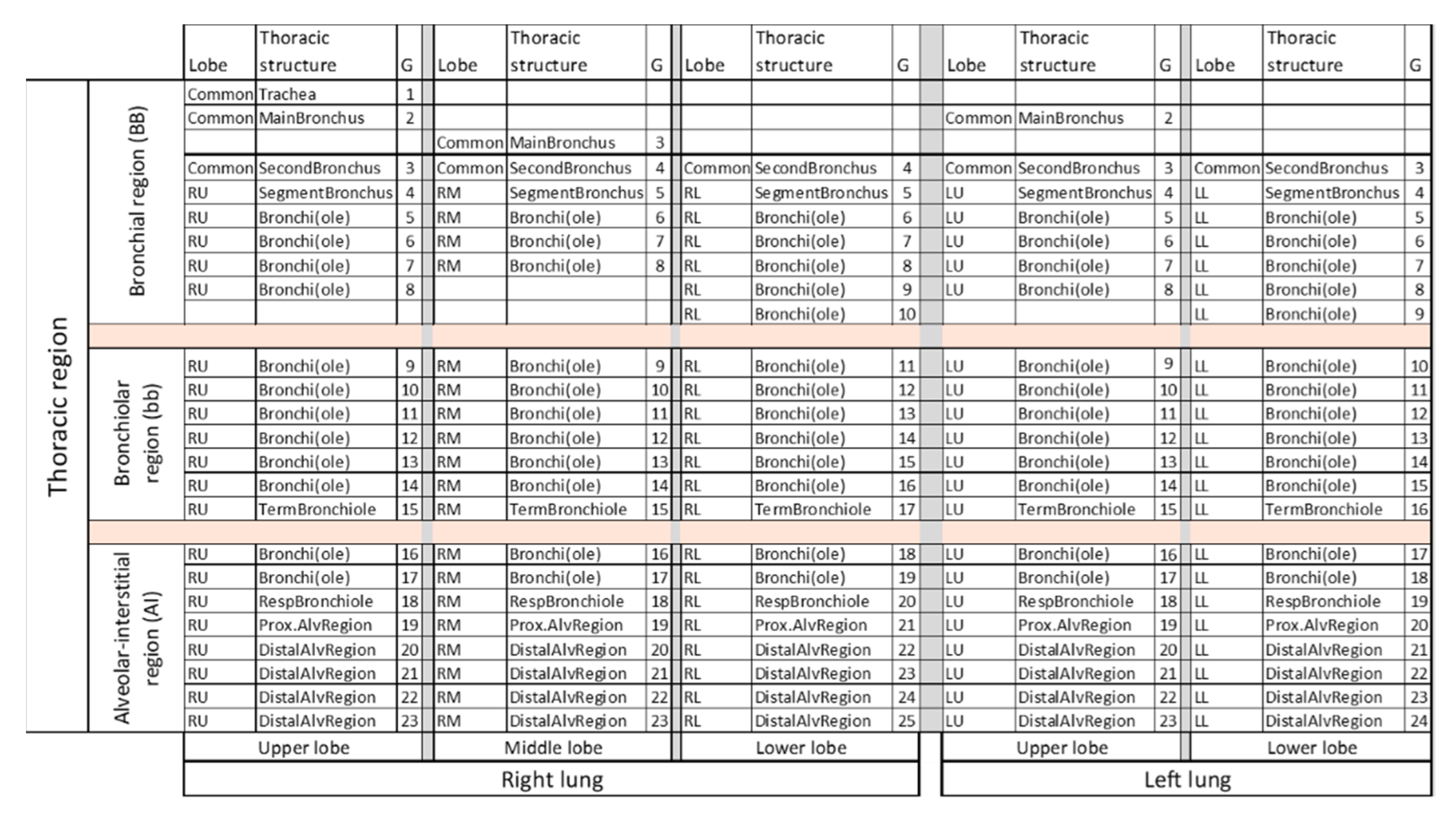
2.2. Lung Modeling
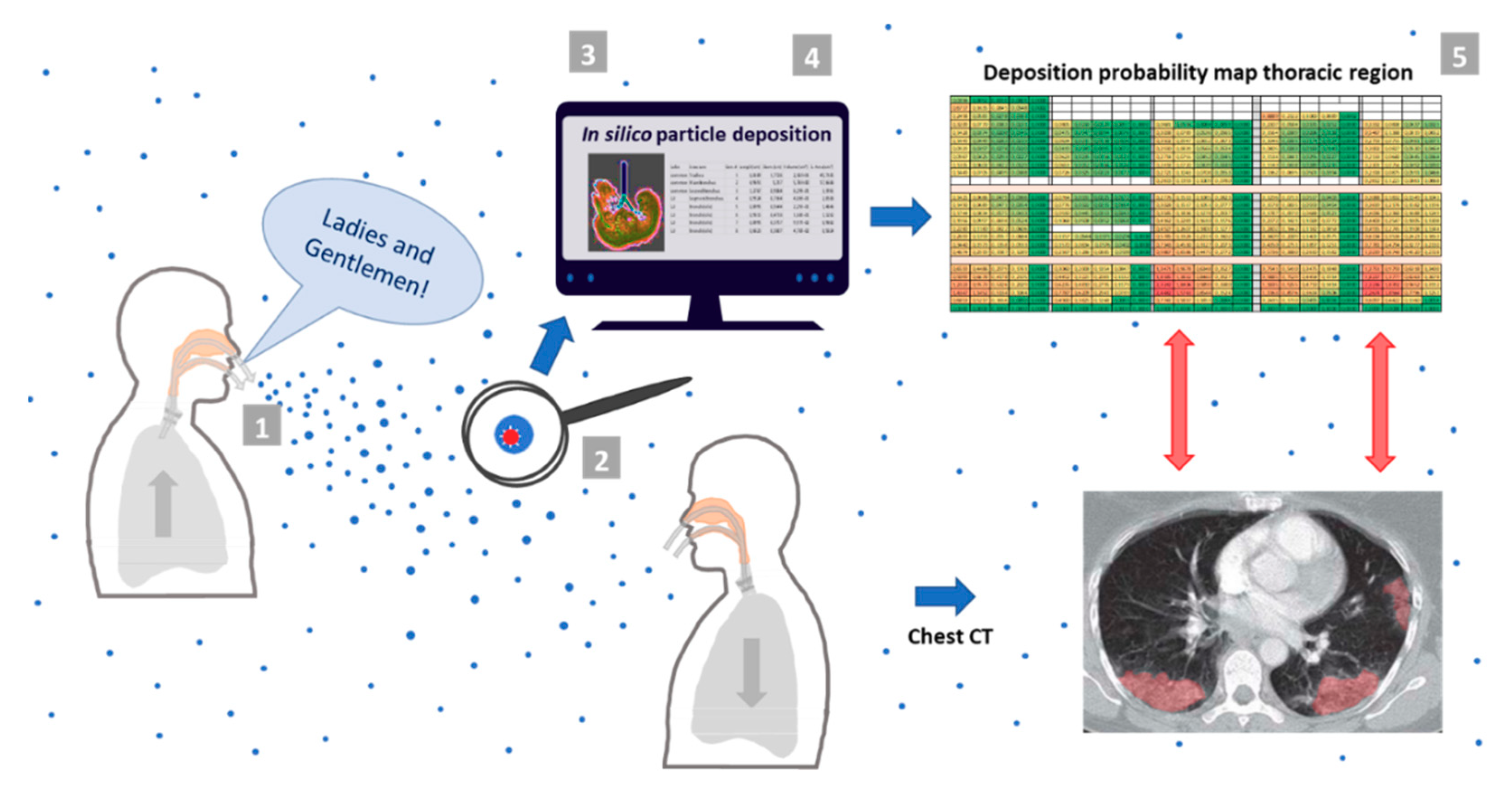
2.3. Deposition Simulation and Analysis
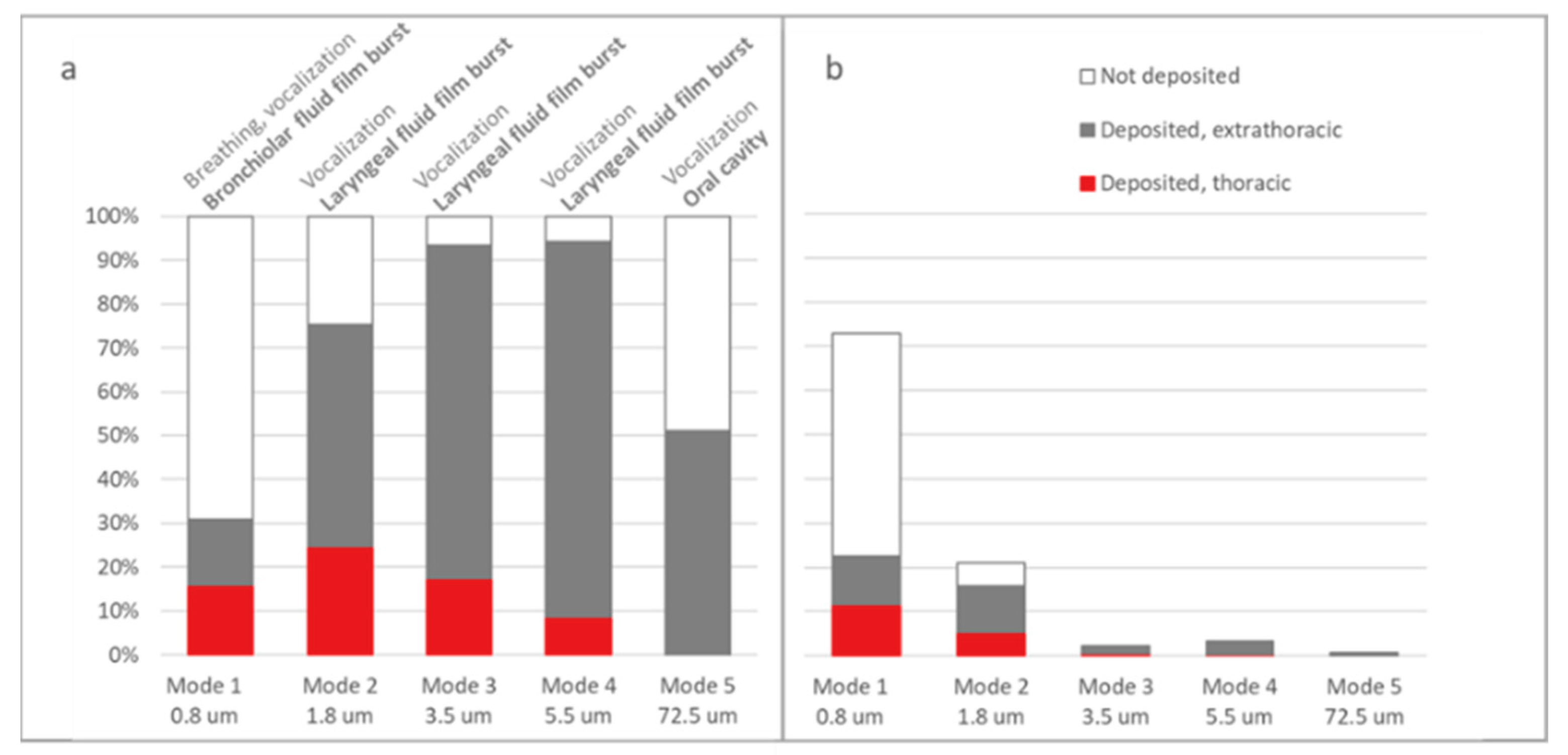
2.4. Simulation of Thoracic ARAP Deposition via in Silico Model MPPD
2.5. Calculation of Extrathoracic Deposition via the ICRP Model
2.6. Calculation of Deposition Probability for Thoracic Deposition Based on Total Deposited Particles
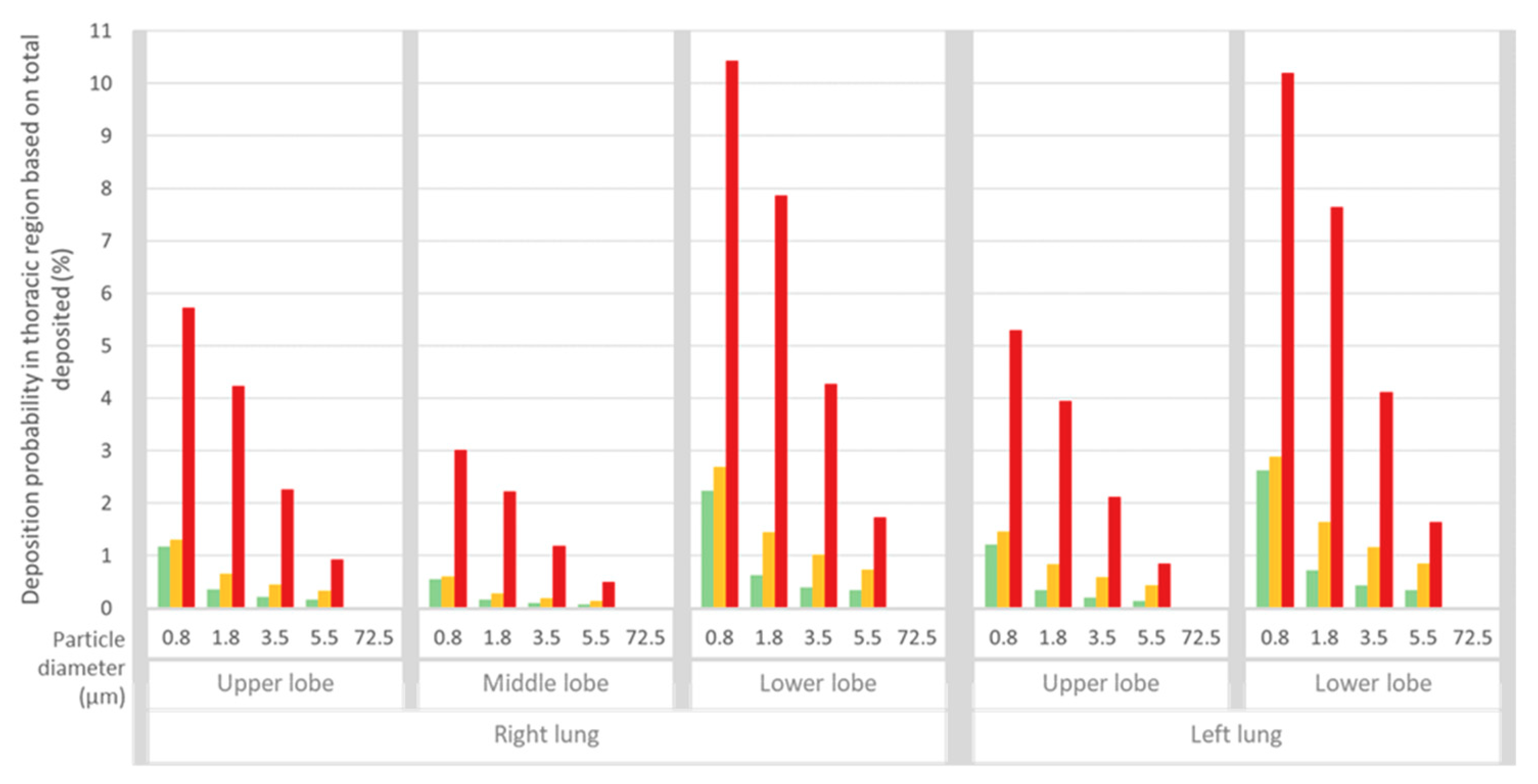
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leung, N.H.; Chu, D.K.; Shiu, E.Y.; Chan, K.-H.; McDevitt, J.J.; Hau, B.J.; Yen, H.-L.; Li, Y.; Ip, D.K.; Peiris, J.M. Respiratory virus shedding in exhaled breath and efficacy of face masks. Nat. Med. 2020, 26, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Milton, D.K.; Fabian, M.P.; Cowling, B.J.; Grantham, M.L.; McDevitt, J.J. Influenza Virus Aerosols in Human Exhaled Breath: Particle Size, Culturability, and Effect of Surgical Masks. PLoS Pathog. 2013, 9, e1003205. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, M.; Hugentobler, W.J.; Iwasaki, A. Seasonality of respiratory viral infections. Annu. Rev. Virol. 2020, 7, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Lin, H.; Wang, J.; Xu, C.; Tatem, A.J.; Meng, B.; Zhang, X.; Liu, Y.; Wang, P.; Wu, G.; et al. Risk of Coronavirus Disease 2019 Transmission in Train Passengers: An Epidemiological and Modeling Study. Clin. Infect. Dis. 2021, 72, 604–610. [Google Scholar] [CrossRef]
- ECDC. COVID-19 Clusters and Outbreaks in Occupational Settings in the EU/EEA and the UK; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2020. [Google Scholar]
- Correa-Martínez, C.L.; Kampmeier, S.; Kümpers, P.; Schwierzeck, V.; Hennies, M.; Hafezi, W.; Kühn, J.; Pavenstädt, H.; Ludwig, S.; Mellmann, A. A pandemic in times of global tourism: Superspreading and exportation of COVID-19 cases from a ski area in Austria. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef]
- Hamner, L. High SARS-CoV-2 attack rate following exposure at a choir practice—Skagit County, Washington, March 2020. Mmwr. Morb. Mortal. Wkly. Rep. 2020, 69, 606–610. [Google Scholar] [CrossRef]
- Anderson, E.L.; Turnham, P.; Griffin, J.R.; Clarke, C.C. Consideration of the Aerosol Transmission for COVID-19 and Public Health. Risk Anal. 2020, 40, 902–907. [Google Scholar] [CrossRef]
- Alwan, N.A.; Burgess, R.A.; Ashworth, S.; Beale, R.; Bhadelia, N.; Bogaert, D.; Dowd, J.; Eckerle, I.; Goldman, L.R.; Greenhalgh, T.; et al. Scientific consensus on the COVID-19 pandemic: We need to act now. Lancet 2020, 396, e71–e72. [Google Scholar] [CrossRef]
- Meyerowitz, E.A.; Richterman, A.; Gandhi, R.T.; Sax, P.E. Transmission of SARS-CoV-2: A review of viral, host, and environmental factors. Ann. Intern. Med. 2020, M20-5008. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Johnson, G.R.; Morawska, L.; Ristovski, Z.D.; Hargreaves, M.; Mengersen, K.; Chao, C.Y.H.; Wan, M.P.; Li, Y.; Xie, X.; Katoshevski, D.; et al. Modality of human expired aerosol size distributions. J. Aerosol Sci. 2011, 42, 839–851. [Google Scholar] [CrossRef]
- Wei, J.; Li, Y. Airborne spread of infectious agents in the indoor environment. Am. J. Infect. Control 2016, 44, S102–S108. [Google Scholar] [CrossRef] [PubMed]
- Morawska, L.; Johnson, G.R.; Ristovski, Z.D.; Hargreaves, M.; Mengersen, K.; Corbett, S.; Chao, C.Y.H.; Li, Y.; Katoshevski, D. Size distribution and sites of origin of droplets expelled from the human respiratory tract during expiratory activities. J. Aerosol Sci. 2009, 40, 256–269. [Google Scholar] [CrossRef]
- Xie, X.; Li, Y.; Chwang, A.; Ho, P.; Seto, W. How far droplets can move in indoor environments-revisiting the Wells evaporation-falling curve. Indoor Air 2007, 17, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Balachandar, S.; Zaleski, S.; Soldati, A.; Ahmadi, G.; Bourouiba, L. Host-to-host airborne transmission as a multiphase flow problem for science-based social distance guidelines. Int. J. Multiph. Flow 2020, 132, 103439. [Google Scholar] [CrossRef]
- Aguiar, J.A.; Tremblay, B.J.M.; Mansfield, M.J.; Woody, O.; Lobb, B.; Banerjee, A.; Chandiramohan, A.; Tiessen, N.; Cao, Q.; Dvorkin-Gheva, A.; et al. Gene expression and in situ protein profiling of candidate SARS-CoV-2 receptors in human airway epithelial cells and lung tissue. Eur. Respir. J. 2020, 56, 2001123. [Google Scholar] [CrossRef]
- Freni, F.; Meduri, A.; Gazia, F.; Nicastro, V.; Galletti, C.; Aragona, P.; Galletti, C.; Galletti, B.; Galletti, F. Symptomatology in head and neck district in coronavirus disease (COVID-19): A possible neuroinvasive action of SARS-CoV-2. Am. J. Otolaryngol. 2020, 41, 102612. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.J. Thoughts on the alveolar phase of COVID-19. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L115–L120. [Google Scholar] [CrossRef]
- Mason, R.J. Pathogenesis of COVID-19 from a cell biology perspective. Eur. Respir. J. 2020, 55, 2000607. [Google Scholar] [CrossRef]
- Donaldson, K.; Borm, P.; Oberdorster, G.; Pinkerton, K.E.; Stone, V.; Tran, C. Concordance between in vitro and in vivo dosimetry in the proinflammatory effects of low-toxicity, low-solubility particles: The key role of the proximal alveolar region. Inhal. Toxicol. 2008, 20, 53–62. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: Summary of a report of 72 314 cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Bai, H.X.; Wang, R.; Xiong, Z.; Hsieh, B.; Chang, K.; Halsey, K.; Tran, T.M.L.; Choi, J.W.; Wang, D.-C.; Shi, L.-B. Artificial Intelligence augmentation of radiologist performance in distinguishing COVID-19 from pneumonia of other origin on chest CT. Radiology 2020, 201491. [Google Scholar] [CrossRef] [PubMed]
- Lieveld, A.; Kok, B.; Schuit, F.; Azijli, K.; Heijmans, J.; van Laarhoven, A.; Assman, N.; Kootte, R.; Olgers, T.; Nanayakkara, P. Diagnosing COVID-19 pneumonia in a pandemic setting: Lung Ultrasound versus CT (LUVCT) A multi-centre, prospective, observational study. ERJ Open Res. 2020, 6, 539. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Liu, H.; Gu, Y.; Guo, X.; Li, H.; Li, L.; Wu, Z.; Liu, M.; Tao, L. Fast automated detection of COVID-19 from medical images using convolutional neural networks. Commun. Biol. 2021, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Han, X.; Jiang, N.; Cao, Y.; Alwalid, O.; Gu, J.; Fan, Y.; Zheng, C. Radiological findings from 81 patients with COVID-19 pneumonia in Wuhan, China: A descriptive study. Lancet Infect. Dis. 2020, 20, 425–434. [Google Scholar] [CrossRef]
- Zu, Z.Y.; Jiang, M.D.; Xu, P.P.; Chen, W.; Ni, Q.Q.; Lu, G.M.; Zhang, L.J. Coronavirus disease 2019 (COVID-19): A perspective from China. Radiology 2020, 296, 200490. [Google Scholar] [CrossRef]
- Sharma, A.; Eisen, J.E.; Shepard, J.-A.O.; Bernheim, A.; Little, B.P. Case 25-2020: A 47-Year-Old Woman with a Lung Mass. N. Engl. J. Med. 2020, 383, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.-Y.; Lee, E.Y.; Yang, J.; Yang, F.; Li, X.; Wang, H.; Lui, M.M.-s.; Lo, C.S.-Y.; Leung, B.; Khong, P.-L. Imaging profile of the COVID-19 infection: Radiologic findings and literature review. Radiol. Cardiothorac. Imaging 2020, 2, e200034. [Google Scholar] [CrossRef]
- Bai, H.X.; Hsieh, B.; Xiong, Z.; Halsey, K.; Choi, J.W.; Tran, T.M.L.; Pan, I.; Shi, L.B.; Wang, D.C.; Mei, J.; et al. Performance of Radiologists in Differentiating COVID-19 from Non-COVID-19 Viral Pneumonia at Chest CT. Radiology 2020, 296, E46–E54. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dong, C.; Hu, Y.; Li, C.; Ren, Q.; Zhang, X.; Shi, H.; Zhou, M. Temporal changes of CT findings in 90 patients with COVID-19 pneumonia: A longitudinal study. Radiology 2020, 296, 200843. [Google Scholar] [CrossRef]
- Bernheim, A.; Mei, X.; Huang, M.; Yang, Y.; Fayad, Z.A.; Zhang, N.; Diao, K.; Lin, B.; Zhu, X.; Li, K. Chest CT findings in coronavirus disease-19 (COVID-19): Relationship to duration of infection. Radiology 2020, 295, 200463. [Google Scholar] [CrossRef]
- Yeh, H.-C.; Schum, G. Models of human lung airways and their application to inhaled particle deposition. Bull. Math. Biol. 1980, 42, 461–480. [Google Scholar] [CrossRef]
- Anjilvel, S.; Asgharian, B. A multiple-path model of particle deposition in the rat lung. Fundam. Appl. Toxicol. 1995, 28, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Miller, F.J.; Asgharian, B.; Schroeter, J.D.; Price, O. Improvements and additions to the multiple path particle dosimetry model. J. Aerosol Sci. 2016, 99, 14–26. [Google Scholar] [CrossRef]
- ICRP. ICRP Publication 66: Human Respiratory Tract Model for Radiological Protection; Ann: ICRP 24(1-3); ICRP: Stockholm, Sweden, 1994; Volume 66. [Google Scholar]
- Asadi, S.; Wexler, A.S.; Cappa, C.D.; Barreda, S.; Bouvier, N.M.; Ristenpart, W.D. Aerosol emission and superemission during human speech increase with voice loudness. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Thomas, R.J. Particle size and pathogenicity in the respiratory tract. Virulence 2013, 4, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Nicas, M.; Nazaroff, W.W.; Hubbard, A. Toward Understanding the Risk of Secondary Airborne Infection: Emission of Respirable Pathogens. J. Occup. Environ. Hyg. 2005, 2, 143–154. [Google Scholar] [CrossRef]
- Holmgren, H.; Bake, B.; Olin, A.-C.; Ljungström, E. Relation Between Humidity and Size of Exhaled Particles. J. Aerosol Med. Pulm. Drug Deliv. 2011, 24, 253–260. [Google Scholar] [CrossRef]
- Thurber, R.V.; Haynes, M.; Breitbart, M.; Wegley, L.; Rohwer, F. Laboratory procedures to generate viral metagenomes. Nat. Protoc. 2009, 4, 470. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Flamholz, A.; Phillips, R.; Milo, R. SARS-CoV-2 (COVID-19) by the numbers. eLife 2020, 9, e57309. [Google Scholar] [CrossRef]
- Couch, R.B.; Knight, V.; Douglas, R.G., Jr.; Black, S.H.; Hamory, B.H. The minimal infectious dose of adenovirus type 4; the case for natural transmission by viral aerosol. Trans. Am. Clin. Clim. Assoc. 1969, 80, 205–211. [Google Scholar]
- Poon, W.C.K.; Brown, A.T.; Direito, S.O.L.; Hodgson, D.J.M.; Le Nagard, L.; Lips, A.; MacPhee, C.E.; Marenduzzo, D.; Royer, J.R.; Silva, A.F.; et al. Soft matter science and the COVID-19 pandemic. Soft Matter. 2020, 16, 8310–8324. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.K.; Wang, Y.-Y.; Wirtz, D.; Hanes, J. Micro- and macrorheology of mucus. Adv. Drug Deliv. Rev. 2009, 61, 86–100. [Google Scholar] [CrossRef]
- App, E.; Zayas, J.; King, M. Rheology of mucus and transepithelial potential difference: Small airways versus trachea. Eur. Respir. J. 1993, 6, 67–75. [Google Scholar]
- Hussain, M.; Madl, P.; Khan, A. Lung deposition predictions of airborne particles and the emergence of contemporary diseases, Part-I. Health 2011, 2, 51–59. [Google Scholar]
- Islam, M.S.; Paul, G.; Ong, H.X.; Young, P.M.; Gu, Y.T.; Saha, S.C. A Review of Respiratory Anatomical Development, Air Flow Characterization and Particle Deposition. Int. J. Envrion. Res. Public Health 2020, 17, 380. [Google Scholar] [CrossRef] [PubMed]
- Mutuku, J.K.; Hou, W.-C.; Chen, W.-H. An Overview of Experiments and Numerical Simulations on Airflow and Aerosols Deposition in Human Airways and the Role of Bioaerosol Motion in COVID-19 Transmission. Aerosol Air Qual. Res. 2020, 20, 1172–1196. [Google Scholar] [CrossRef]
- Koullapis, P.; Stylianou, F.; Sznitman, J.; Olsson, B.; Kassinos, S. Towards whole-lung simulations of aerosol deposition: A model of the deep lung. J. Aerosol Sci. 2020, 105541. [Google Scholar] [CrossRef]
- Knudsen, L.; Ochs, M. The micromechanics of lung alveoli: Structure and function of surfactant and tissue components. Histochem. Cell Biol. 2018, 150, 661–676. [Google Scholar] [CrossRef]
- Atanasova, K.R.; Reznikov, L.R. Strategies for measuring airway mucus and mucins. Respir. Res. 2019, 20, 261. [Google Scholar] [CrossRef]
- Basu, S. Computational characterization of inhaled droplet transport to the nasopharynx. Sci. Rep. 2021, 11, 6652. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Bartrand, T.A.; Weir, M.H.; Omura, T.; Haas, C.N. Development of a Dose-Response Model for SARS Coronavirus. Risk Anal. 2010, 30, 1129–1138. [Google Scholar] [CrossRef]
- Miller, S.L.; Nazaroff, W.W.; Jimenez, J.L.; Boerstra, A.; Buonanno, G.; Dancer, S.J.; Kurnitski, J.; Marr, L.C.; Morawska, L.; Noakes, C. Transmission of SARS-CoV-2 by inhalation of respiratory aerosol in the Skagit Valley Chorale superspreading event. Indoor Air 2021, 31, 314–323. [Google Scholar] [CrossRef]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 2020, 181, 1016–1035.e1019. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front. Med. 2020, 14, 1–8. [Google Scholar] [CrossRef]
- Qi, F.; Qian, S.; Zhang, S.; Zhang, Z. Single cell RNA sequencing of 13 human tissues identify cell types and receptors of human coronaviruses. Biochem. Biophys. Res. Commun. 2020, 526, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Hamming, I.; Timens, W.; Bulthuis, M.; Lely, A.; Navis, G.V.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. A J. Pathol. Soc. Great Br. Irel. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Wruck, W.; Adjaye, J. SARS-CoV-2 receptor ACE2 is co-expressed with genes related to transmembrane serine proteases, viral entry, immunity and cellular stress. Sci. Rep. 2020, 10, 21415. [Google Scholar] [CrossRef]
- Unterholzner, L.; Keating, S.E.; Baran, M.; Horan, K.A.; Jensen, S.B.; Sharma, S.; Sirois, C.M.; Jin, T.; Latz, E.; Xiao, T.S.; et al. IFI16 is an innate immune sensor for intracellular DNA. Nat. Immunol. 2010, 11, 997–1004. [Google Scholar] [CrossRef]
- Kerur, N.; Veettil, M.V.; Sharma-Walia, N.; Bottero, V.; Sadagopan, S.; Otageri, P.; Chandran, B. IFI16 acts as a nuclear pathogen sensor to induce the inflammasome in response to Kaposi Sarcoma-associated herpesvirus infection. Cell Host Microbe 2011, 9, 363–375. [Google Scholar] [CrossRef]
- Yazicioglu, T.; Mühlfeld, C.; Autilio, C.; Huang, C.-K.; Bär, C.; Dittrich-Breiholz, O.; Thum, T.; Pérez-Gil, J.; Schmiedl, A.; Brandenberger, C. Aging impairs alveolar epithelial type II cell function in acute lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 319, L755–L769. [Google Scholar] [CrossRef]
- Thompson, R.B.; Finlay, W.H. Using MRI to measure aerosol deposition. J. Aerosol Med. Pulm. Drug Deliv. 2012, 25, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Biddiscombe, M.F.; Meah, S.N.; Underwood, S.R.; Usmani, O.S. Comparing lung regions of interest in gamma scintigraphy for assessing inhaled therapeutic aerosol deposition. J. Aerosol Med. Pulm. Drug Deliv. 2011, 24, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Buonanno, G.; Stabile, L.; Morawska, L. Estimation of airborne viral emission: Quanta emission rate of SARS-CoV-2 for infection risk assessment. Environ. Int. 2020, 141, 105794. [Google Scholar] [CrossRef] [PubMed]
- Kidd, M.; Richter, A.; Best, A.; Mirza, J.; Percival, B.; Mayhew, M.; Megram, O.; Ashford, F.; White, T.; Moles-Garcia, E. S-variant SARS-CoV-2 is associated with significantly higher viral loads in samples tested by ThermoFisher TaqPath RT-QPCR. medRxiv 2020. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Fraction Per Weight (%) | Mean Fraction Per Weight (%) (a) | Density (g/cm3) |
|---|---|---|---|
| Protein (mucin) | 2–5 | 3.5 | 1.35 |
| Carbohydrate (glycan) | 7.5–9 | 8.3 | 1.5 |
| Lipid | 1–2 | 1.5 | 0.985 |
| Ions | 1 | 1 | 1.409 |
| H2O | 80–90 (b) | 85.7 | 1 |
| Mucus composition | 1.041 |
| Mode | Diameter (µm) | Anatomical Origin [13] | Mass (g) | Volume (µm3) | Density (g/cm3) |
|---|---|---|---|---|---|
| Mode 1—breathing, vocalization | 0.8 | Bronchiolar fluid film burst | 3.57 × 10−13 | 0.269 | 1.328 |
| Mode 2—vocalization | 1.8 | Laryngeal fluid film burst | 4.057 × 10−12 | 3.054 | 1.328 |
| Mode 3—vocalization | 3.5 | Laryngeal fluid film burst | 2.982 × 10−11 | 22.45 | 1.328 |
| Mode 4—vocalization | 5.5 | Laryngeal fluid film burst | 1.157 × 10−10 | 87.115 | 1.328 |
| Mode 5—vocalization | 72.5 | Oral cavity | 2.6507 × 10−7 | 199,532.040 | 1.328 |
| Input Section | Scenario | Parameter | Value Setting |
|---|---|---|---|
| Airway Morphometry | Aerosol | Model | Yeh/Schum 5-Lobe |
| Inhalant Properties | Constant Exposure | FRC | 3300 mL |
| Exposure Condition | URT | 50 mL | |
| Density | 1.328 g/cm3 | ||
| Aspect Ratio | 1.0 (=spherical) | ||
| Diameter | 0.8, 1.8, 3.5, 5.5, 72.5 µm (a) | ||
| Body Orientation | Upright | ||
| Aerosol Concentration | 0.5 mg/m3 (b) | ||
| Breathing Frequency | 15 per minute | ||
| Tidal Volume | 750 mL | ||
| Inspiratory Fraction | 0.5 | ||
| Pause Fraction | 0 | ||
| Breathing Scenario | Oronasal-Normal Augmenter | ||
| Deposition/Clearance | Deposition Only |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hofer, S.; Hofstätter, N.; Duschl, A.; Himly, M. SARS-CoV-2-Laden Respiratory Aerosol Deposition in the Lung Alveolar-Interstitial Region Is a Potential Risk Factor for Severe Disease: A Modeling Study. J. Pers. Med. 2021, 11, 431. https://doi.org/10.3390/jpm11050431
Hofer S, Hofstätter N, Duschl A, Himly M. SARS-CoV-2-Laden Respiratory Aerosol Deposition in the Lung Alveolar-Interstitial Region Is a Potential Risk Factor for Severe Disease: A Modeling Study. Journal of Personalized Medicine. 2021; 11(5):431. https://doi.org/10.3390/jpm11050431
Chicago/Turabian StyleHofer, Sabine, Norbert Hofstätter, Albert Duschl, and Martin Himly. 2021. "SARS-CoV-2-Laden Respiratory Aerosol Deposition in the Lung Alveolar-Interstitial Region Is a Potential Risk Factor for Severe Disease: A Modeling Study" Journal of Personalized Medicine 11, no. 5: 431. https://doi.org/10.3390/jpm11050431
APA StyleHofer, S., Hofstätter, N., Duschl, A., & Himly, M. (2021). SARS-CoV-2-Laden Respiratory Aerosol Deposition in the Lung Alveolar-Interstitial Region Is a Potential Risk Factor for Severe Disease: A Modeling Study. Journal of Personalized Medicine, 11(5), 431. https://doi.org/10.3390/jpm11050431