
CDK1 and HSP90AA1 Appear as the Novel Regulatory Genes in Non-Small Cell Lung Cancer: A Bioinformatics Approach

 ,
,  ,
,
Abstract
:1. Introduction
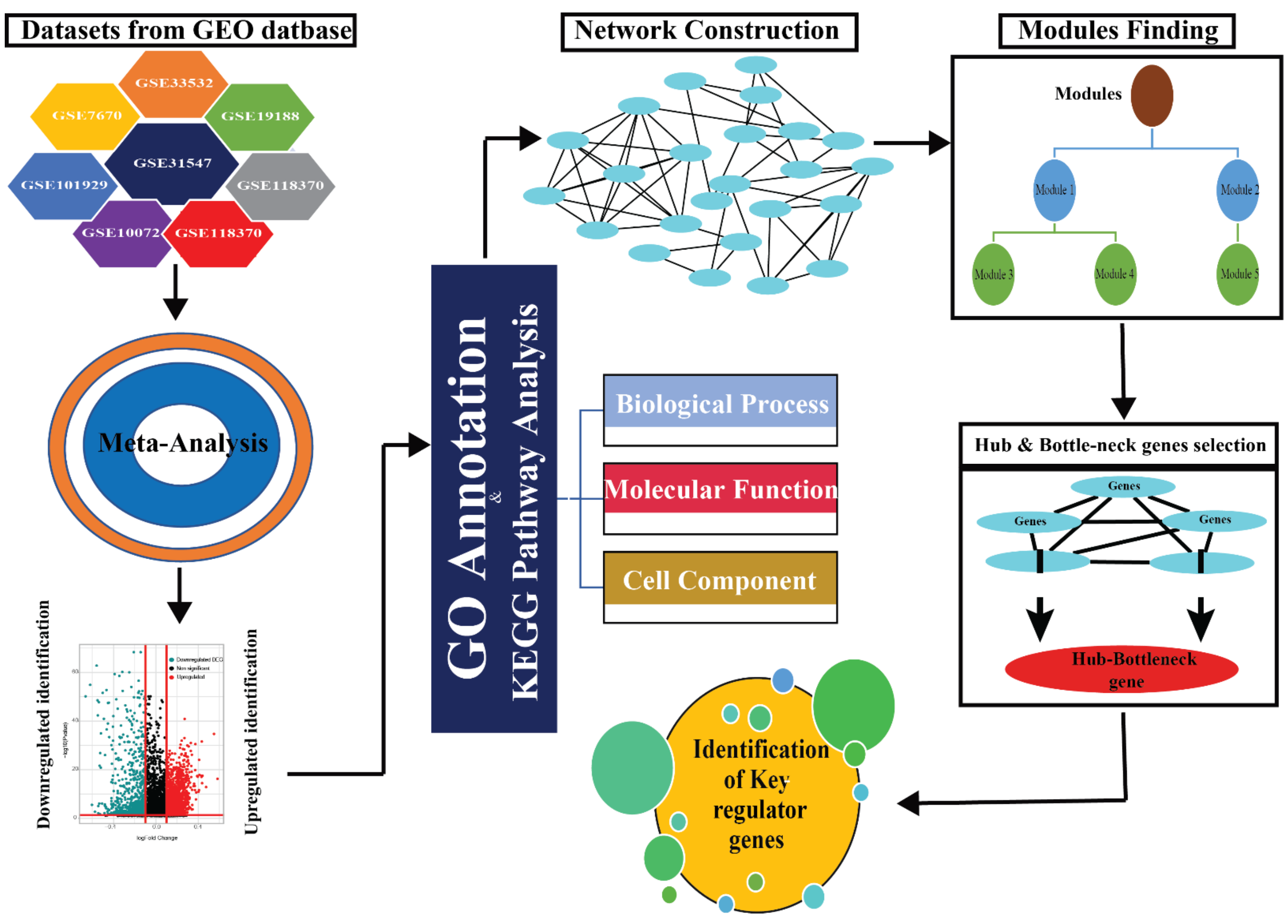
2. Materials and Methods
2.1. Data Collection
2.2. Meta-Analysis of the Datasets
2.3. Gene Ontology Analysis
2.4. PPI Network Construction
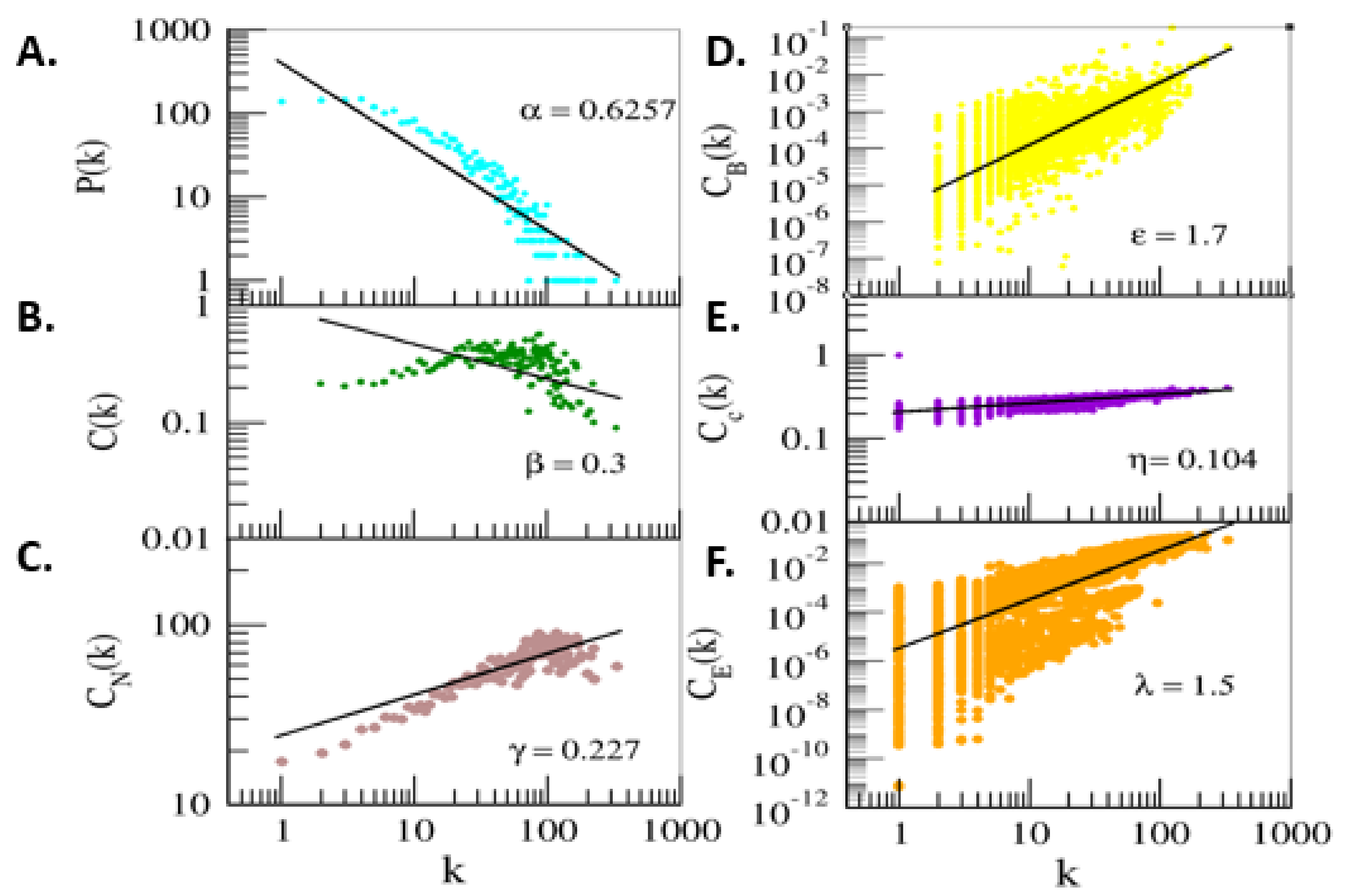
2.5. Network Analysis
2.6. Key Regulator Gene Identification
2.7. Survival Plot Analysis of the Driver Gene
3. Results
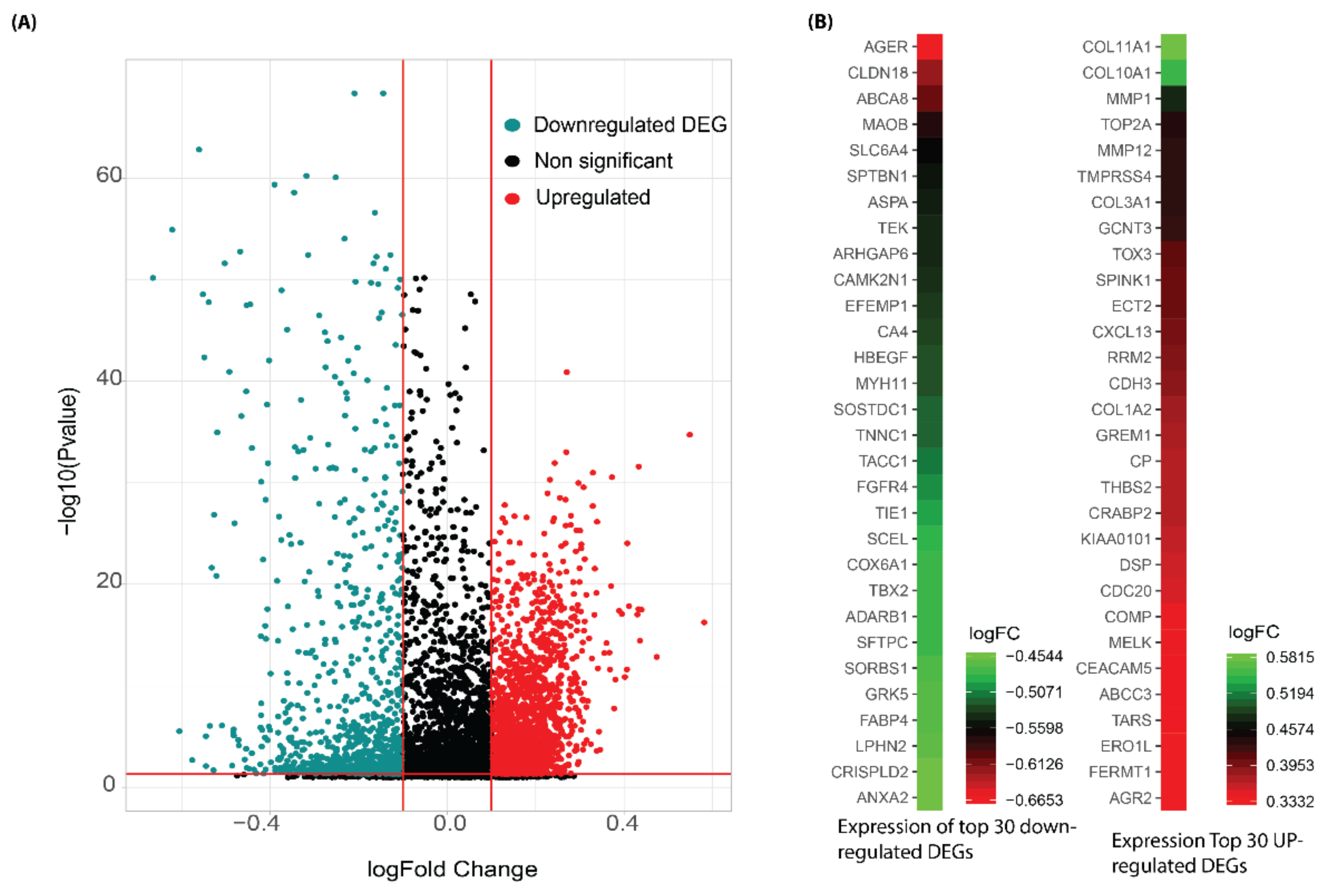
3.1. Identification of Differentially Expressed Genes and Meta-Analysis
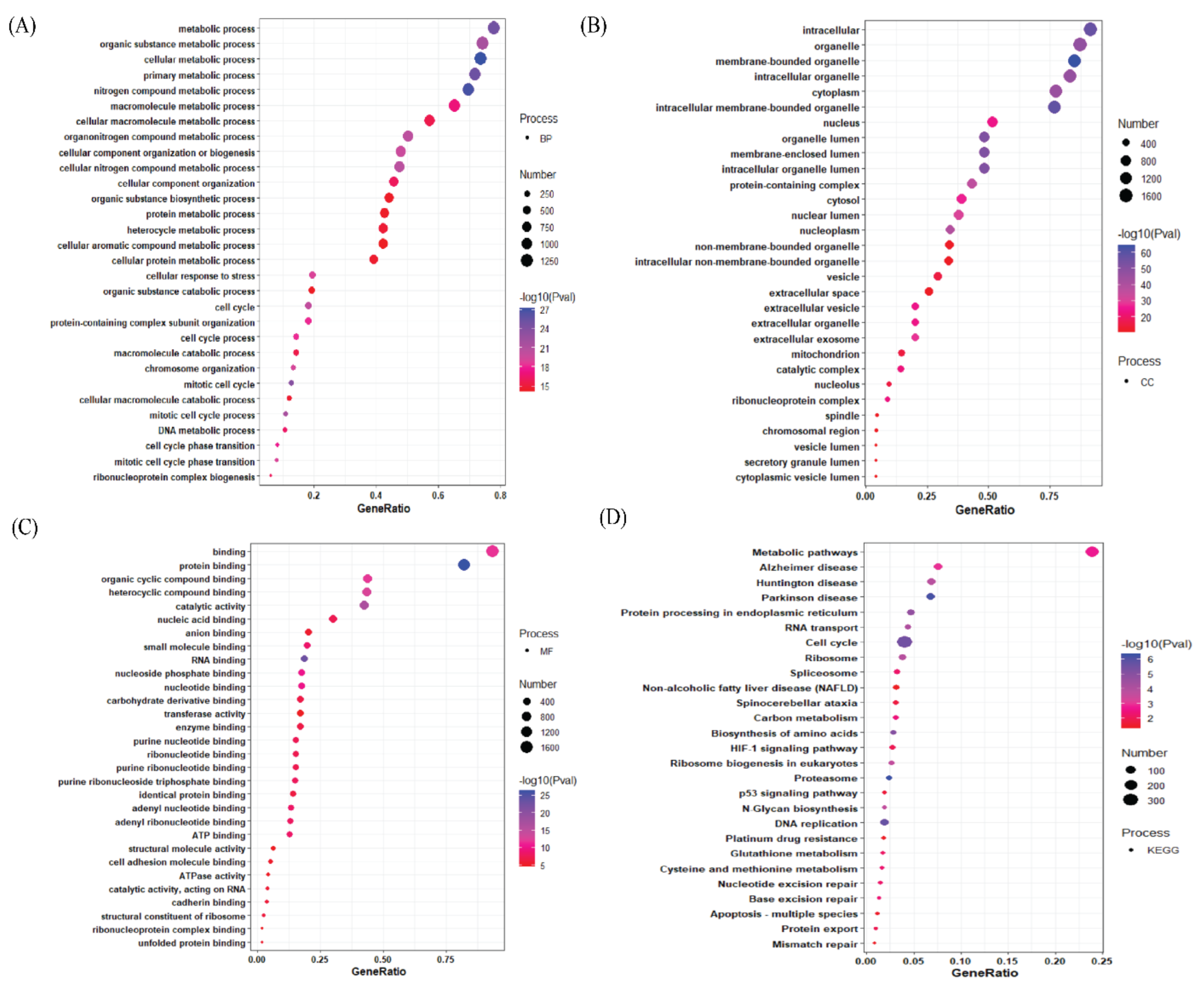
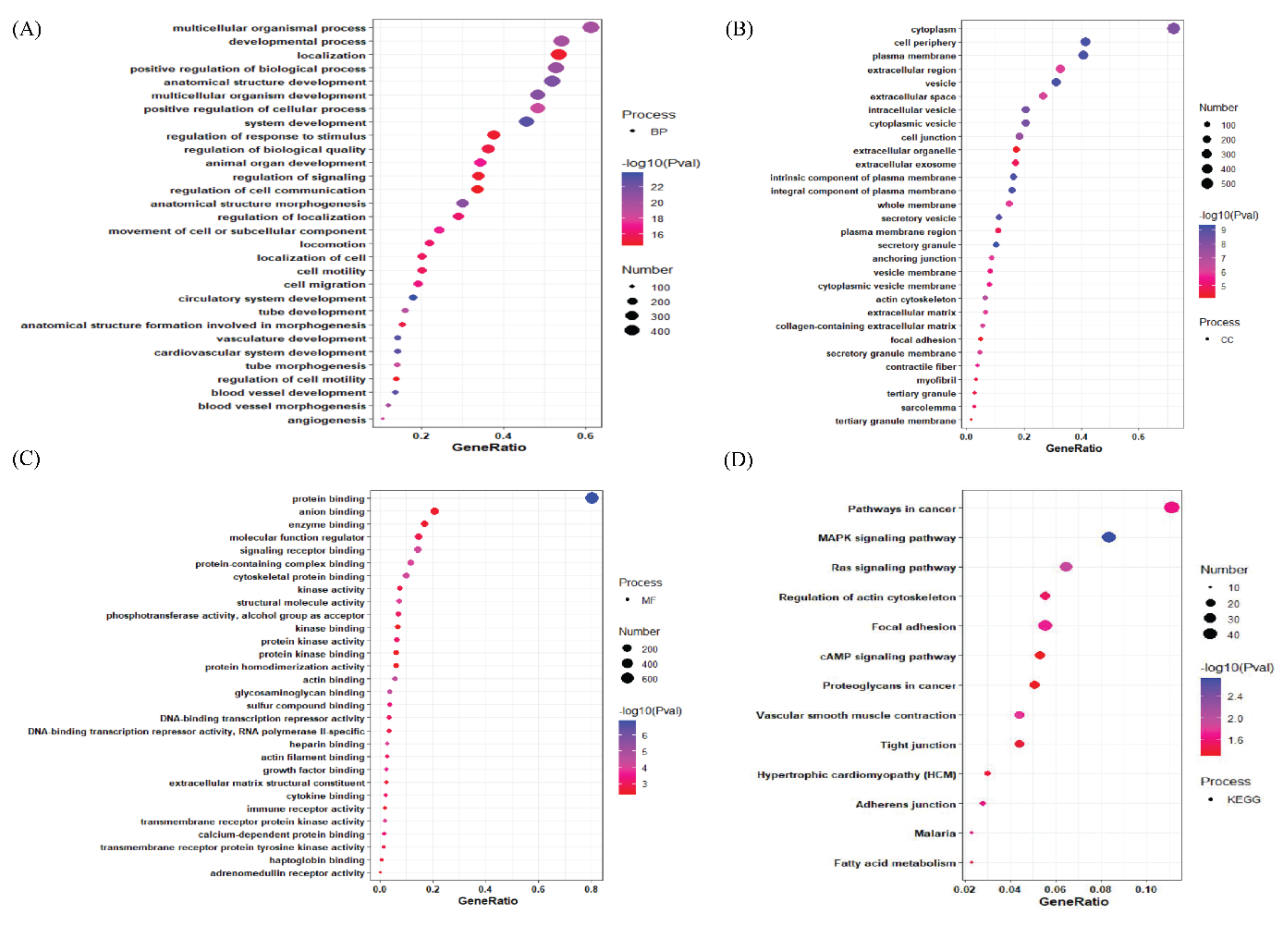
3.2. Gene Ontology Analysis of Differentially Expressed Genes
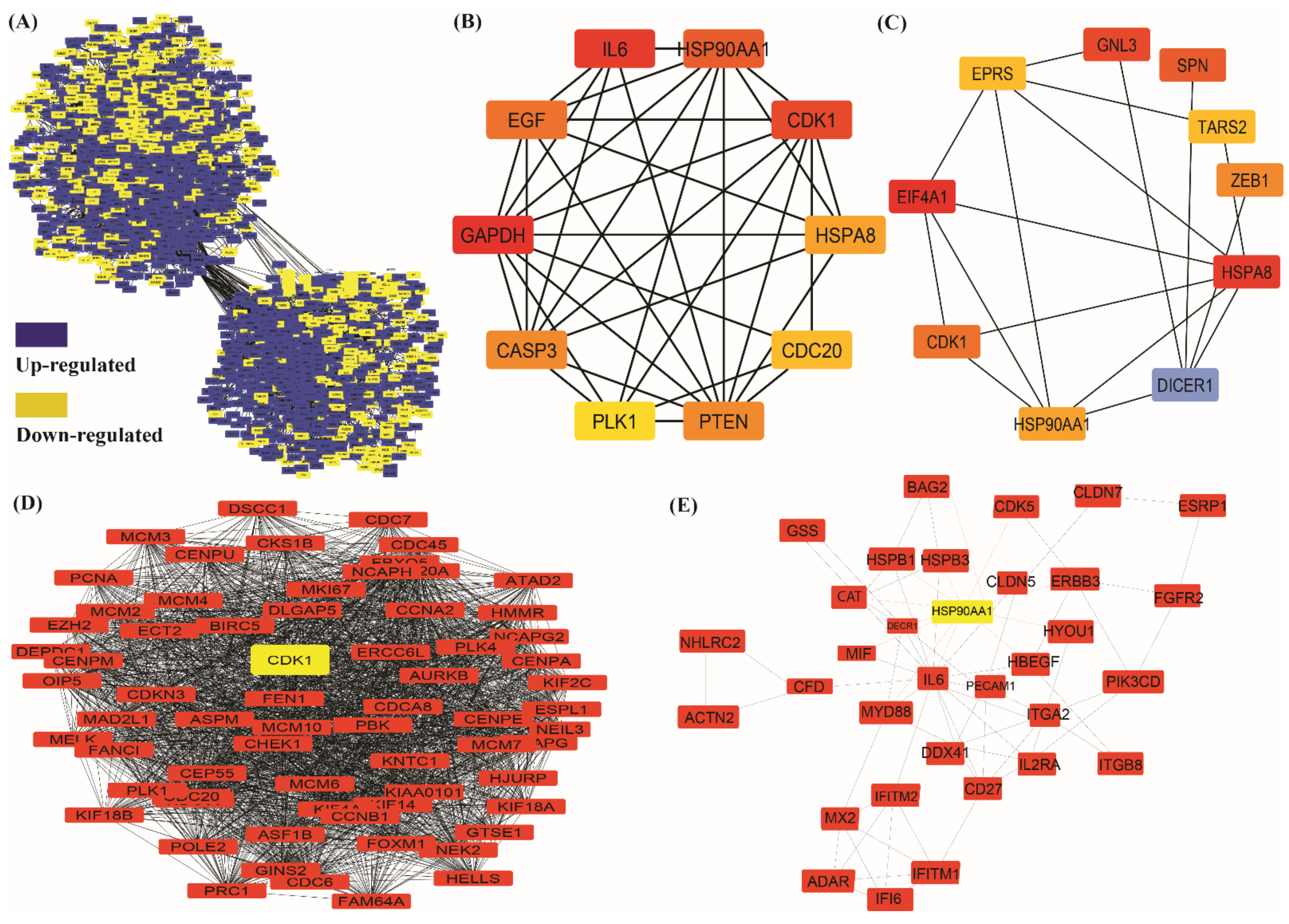
3.3. Protein–Protein Interaction Network Construction and Downstream Analysis
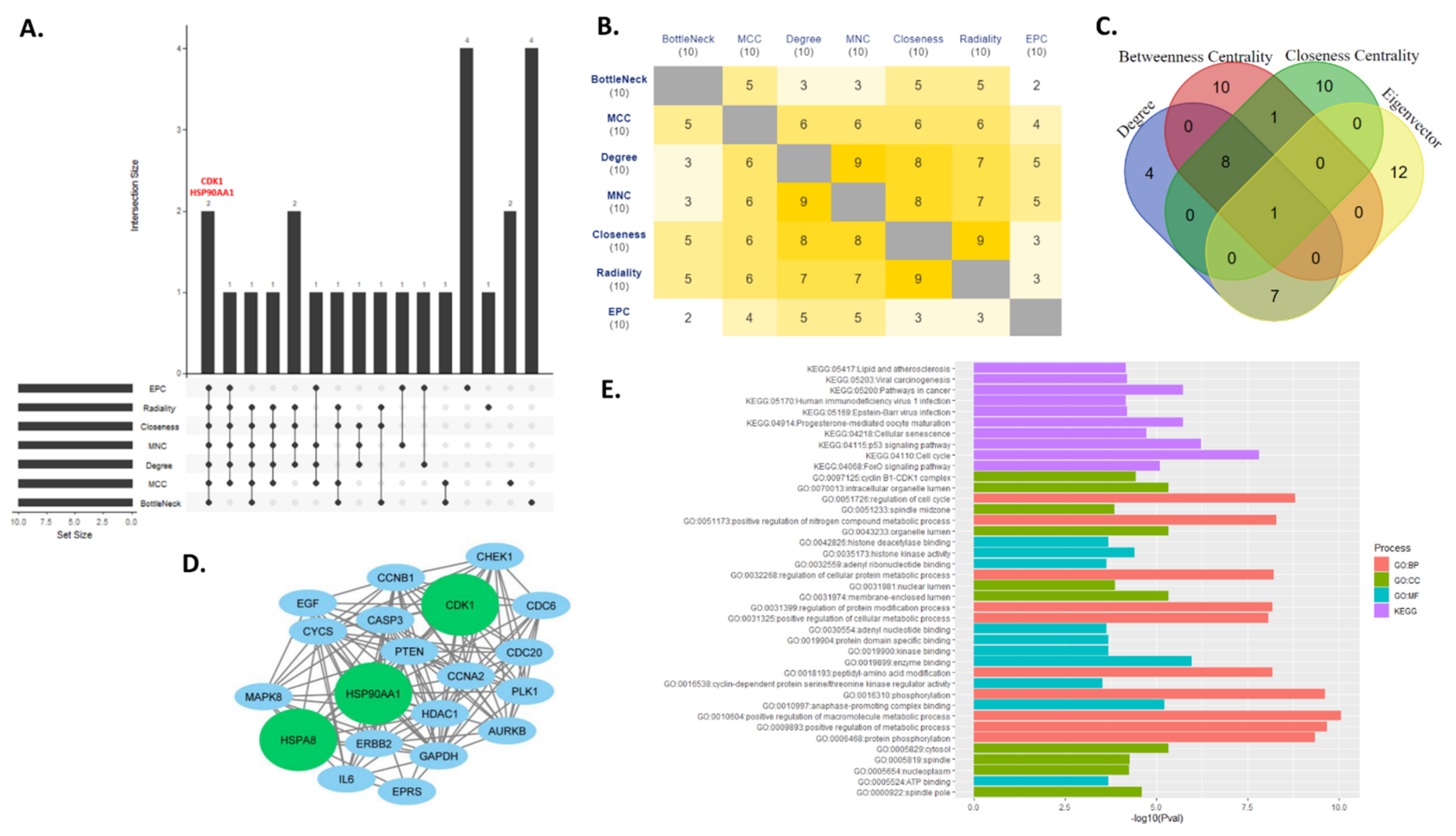
3.4. Key Regulator Gene Identification
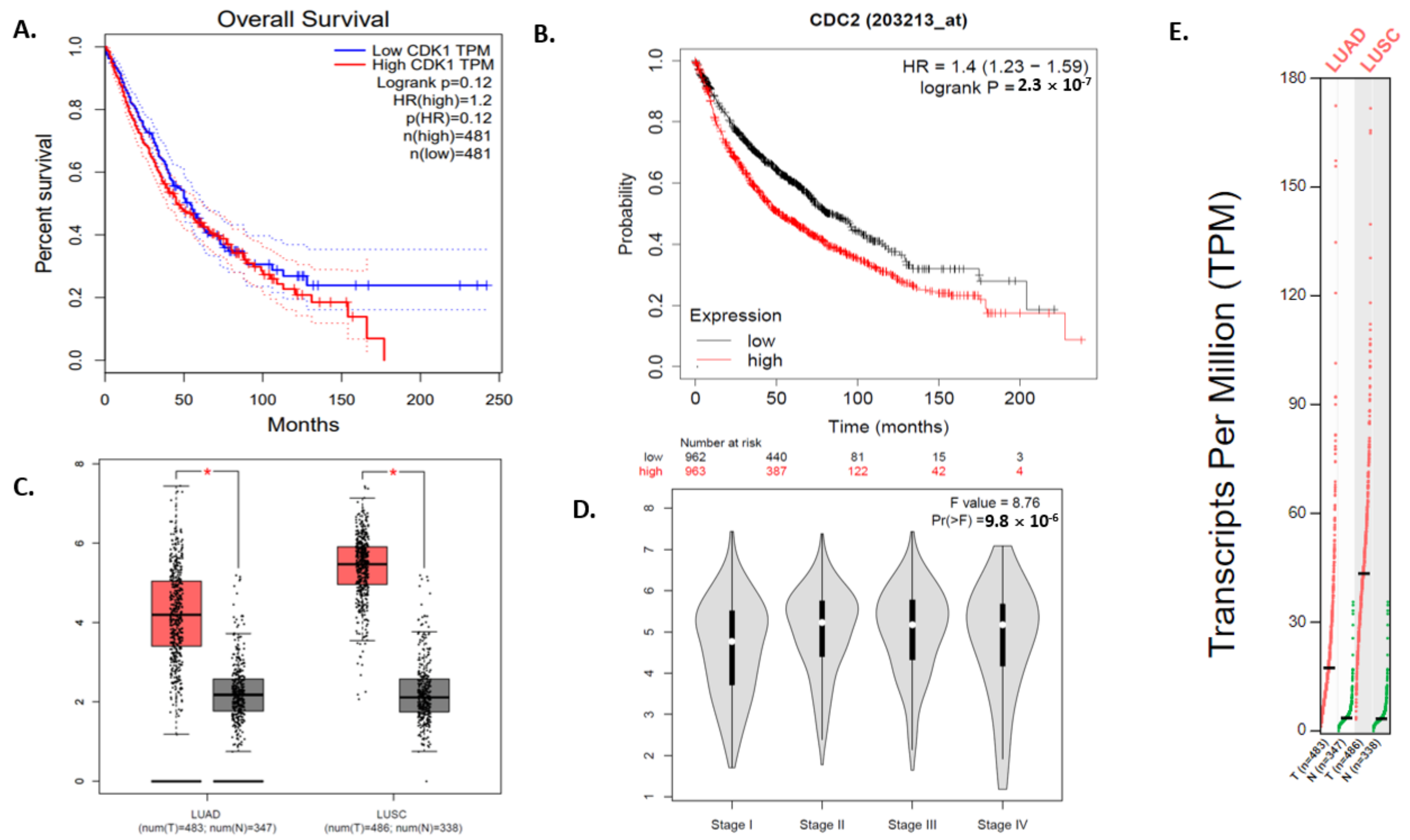
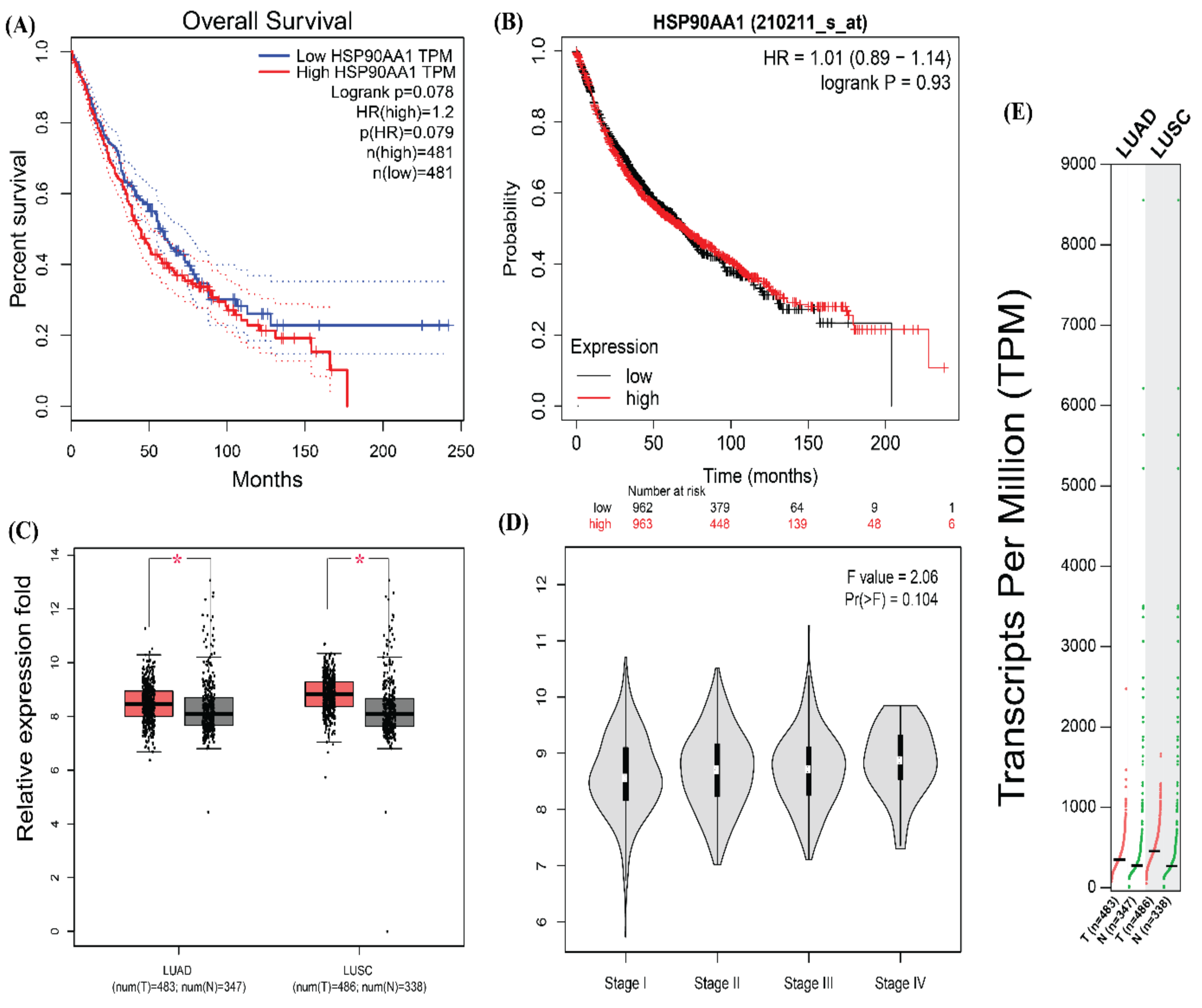
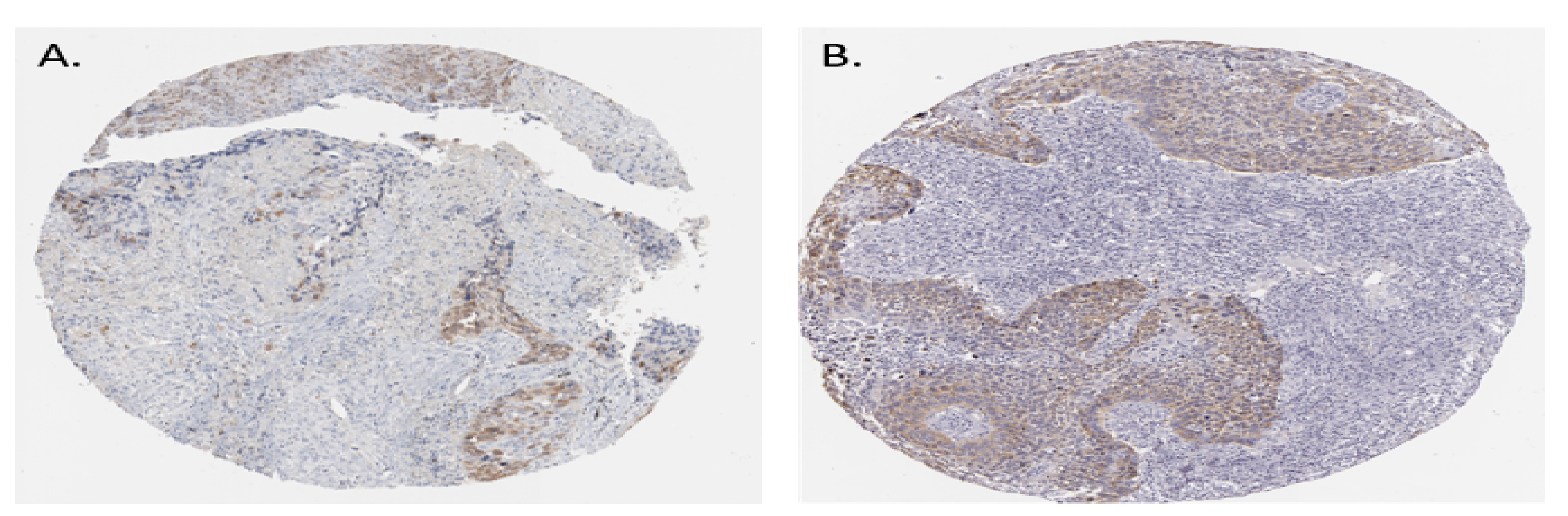
3.5. Survival Plot Analysis of the Driver Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Murray, T.; Thun, M.J. Cancer Statistics, 2008. CA Cancer J. Clin. 2008, 58, 71–96. [Google Scholar] [CrossRef]
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-Small Cell Lung Cancer: Epidemiology, Risk Factors, Treatment, and Survivorship. Mayo. Clin. Proc. 2008, 83, 584–594. [Google Scholar] [CrossRef]
- Chansky, K.; Detterbeck, F.C.; Nicholson, A.G.; Rusch, V.W.; Vallières, E.; Groome, P.; Kennedy, C.; Krasnik, M.; Peake, M.; Shemanski, L.; et al. The IASLC Lung Cancer Staging Project: External Validation of the Revision of the TNM Stage Groupings in the Eighth Edition of the TNM Classification of Lung Cancer. J. Thorac. Oncol. 2017, 12, 1109–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, J.; Govindan, R. Lung Cancer in Never Smokers: A Review. J Clin. Oncol. 2007, 25, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.D.; Pickwell-Smith, B.; Macleod, U. Risk Factors for Emergency Presentation with Lung and Colorectal Cancers: A Systematic Review. BMJ Open 2015, 5, e006965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popper, H.H. Progression and Metastasis of Lung Cancer. Cancer Metastasis Rev. 2016, 35, 75–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenfield, S.A.; Stampfer, M.J.; Rosner, B.A.; Colditz, G.A. Smoking and Smoking Cessation in Relation to Mortality in Women. JAMA 2008, 299, 2037–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Conner, H.; Kobayashi, T.; Kim, H.; Wen, F.; Abe, S.; Fang, Q.; Wang, X.; Hashimoto, M.; Bitterman, P.; et al. Cigarette Smoke Extract Induces DNA Damage but Not Apoptosis in Human Bronchial Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2005, 33, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Coultas, D.B.; Samet, J.M. Occupational Lung Cancer. Clin. Chest Med. 1992, 13, 341–354. [Google Scholar] [CrossRef]
- Edling, C.; Kling, H.; Axelson, O. Radon in Homes--A Possible Cause of Lung Cancer. Scand. J. Work. Environ. Health 1984, 10, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Kastan, M.B. DNA Damage Responses: Mechanisms and Roles in Human Disease: 2007 G.H.A. Clowes Memorial Award Lecture. Mol. Cancer Res. 2008, 6, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hagan, H.M.; Mohammad, H.P.; Baylin, S.B. Double Strand Breaks Can Initiate Gene Silencing and SIRT1-Dependent Onset of DNA Methylation in an Exogenous Promoter CpG Island. PLoS Genet. 2008, 4, e1000155. [Google Scholar] [CrossRef] [PubMed]
- Cuozzo, C.; Porcellini, A.; Angrisano, T.; Morano, A.; Lee, B.; Pardo, A.D.; Messina, S.; Iuliano, R.; Fusco, A.; Santillo, M.R.; et al. DNA Damage, Homology-Directed Repair, and DNA Methylation. PLoS Genet. 2007, 3, e110. [Google Scholar] [CrossRef] [PubMed]
- Do, H.; Wong, N.C.; Murone, C.; John, T.; Solomon, B.; Mitchell, P.L.; Dobrovic, A. A Critical Re-Assessment of DNA Repair Gene Promoter Methylation in Non-Small Cell Lung Carcinoma. Sci. Rep. 2014, 4, 4186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrelo, R.; Cheng, W.H.; Setien, F.; Ropero, S.; Espada, J.; Fraga, M.F.; Herranz, M.; Paz, M.F.; Sanchez-Cespedes, M.; Artiga, M.J.; et al. Epigenetic Inactivation of the Premature Aging Werner Syndrome Gene in Human Cancer. Proc Natl. Acad. Sci. USA 2006, 103, 8822–8827. [Google Scholar] [CrossRef] [Green Version]
- Wolf, P.; Hu, Y.C.; Doffek, K.; Sidransky, D.; Ahrendt, S.A. O(6)-Methylguanine-DNA Methyltransferase Promoter Hypermethylation Shifts the p53 Mutational Spectrum in Non-Small Cell Lung Cancer. Cancer Res. 2001, 61, 8113–8117. [Google Scholar] [PubMed]
- Ekim, M.; Caner, V.; Büyükpınarbaşılı, N.; Tepeli, E.; Elmas, L.; Bağcı, G. Determination of O⁶-Methylguanine DNA Methyltransferase Promoter Methylation in Non-Small Cell Lung Cancer. Genet. Test Mol. Biomark. 2011, 15, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Safar, A.M.; Spencer, H.; Su, X.; Coffey, M.; Cooney, C.; Ratnasinghe, L.D.; Hutchins, L.F.; Fan, C.-Y. Methylation Profiling of Archived Non–Small Cell Lung Cancer: A Promising Prognostic System. Clin. Cancer Res. 2005, 11, 4400–4405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.; Reis-Silva, M.; Alarcão, A.; Couceiro, P.; Sousa, V.; Carvalho, L. Promoter Hypermethylation of DNA Repair Genes MLH1 and MSH2 in Adenocarcinomas and Squamous Cell Carcinomas of the Lung. Rev. Port. Pneumol. 2014, 20, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.-N.; Tseng, R.-C.; Hsu, H.-S.; Chen, J.-Y.; Tzao, C.; Ho, W.L.; Wang, Y.-C. Epigenetic Inactivation of the Chromosomal Stability Control Genes BRCA1, BRCA2, and XRCC5 in Non–Small Cell Lung Cancer. Clin. Cancer Res. 2007, 13, 832–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolova, T.; Christmann, M.; Kaina, B. FEN1 is Overexpressed in Testis, Lung, and Brain Tumors. Anticancer Res. 2009, 29, 2453–2459. [Google Scholar]
- Zarogoulidis, K.; Zarogoulidis, P.; Darwiche, K.; Boutsikou, E.; Machairiotis, N.; Tsakiridis, K.; Katsikogiannis, N.; Kougioumtzi, I.; Karapantzos, I.; Huang, H.; et al. Treatment of Non-Small Cell Lung Cancer (NSCLC). J. Thorac. Dis. 2013, 5 (Suppl. 4), S389–S396. [Google Scholar] [PubMed]
- Hou, J.; Aerts, J.; den Hamer, B.; van Ijcken, W.; den Bakker, M.; Riegman, P.; Van Der Leest, C.; Van Der Spek, P.; Foekens, J.A.; Hoogsteden, H.C.; et al. Gene Expression-Based Classification of Non-Small Cell Lung Carcinomas and Survival Prediction. PLoS ONE 2010, 5, e10312. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lu, C.; Huang, Y.; Zhou, J.; Wang, X.; Liu, C.; Chen, J.; Le, A.H. SPINK1 promotes cell growth and metastasis of lung adenocarcinoma and acts as a novel prognostic biomarker. BMB Rep. 2018, 51, 648–653. [Google Scholar] [CrossRef] [Green Version]
- Landi, M.T.; Dracheva, T.; Rotunno, M.; Figueroa, J.D.; Liu, H.; Dasgupta, A.; Mann, F.E.; Fukuoka, J.; Hames, M.; Bergen, A.; et al. Gene Expression Signature of Cigarette Smoking and Its Role in Lung Adenocarcinoma Development and Survival. PLoS ONE 2008, 3, e1651. [Google Scholar] [CrossRef]
- Mitchell, K.A.; Zingone, A.; Toulabi, L.; Boeckelman, J.; Ryan, B.M. Comparative Transcriptome Profiling Reveals Coding and Noncoding RNA Differences in NSCLC from African Americans and European Americans. Clin. Cancer Res. 2017, 23, 7412–7425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.-J.; Chang, C.-W.; Wu, Y.-C.; Chen, K.-C.; Lin, C.-J.; Liang, S.-C.; Lin, C.-H.; Whang-Peng, J.; Hsu, S.-L.; Chen, C.-H.; et al. Selection of DDX5 as a novel internal control for Q-RT-PCR from microarray data using a block bootstrap re-sampling scheme. BMC Genom. 2007, 8, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.H.; Lai, J.M.; Chou, T.Y.; Chen, C.Y.; Su, L.-J.; Lee, Y.-C.; Cheng, T.-S.; Hong, Y.-R.; Chou, C.-K.; Jacqueline, W.-P.; et al. VEGFA upregulates FLJ10540 and modulates migration and invasion of lung cancer via PI3K/AKT pathway. PLoS ONE 2009, 4, e5052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, M.; Anton, B.; Xu, E.C.; Schnabel, P.; Warth, A.; Hoffmann, H.; Dienemann, H.; Riedlinger, J.; Bodenmueller, H.; Zolg, W.; et al. Intra-tumor Heterogeneity of Gene Expression Profiles in Early Stage Non-Small Cell Lung Cancer. J. Bioinform. Res. Stud. 2014, 1, 1. [Google Scholar]
- Okayama, H.; Kohno, T.; Ishii, Y.; Shimada, Y.; Shiraishi, K.; Iwakawa, R.; Furuta, K.; Tsuta, K.; Shibata, T.; Yamamoto, S.; et al. Identification of Genes Upregulated in ALK-Positive and EGFR/KRAS/ALK-Negative Lung Adenocarcinomas. Cancer Res. 2012, 72, 100–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, M.; Yamaguchi, R.; Nakata, A.; Kohno, T.; Nagasaki, M.; Shimamura, T.; Imoto, S.; Saito, A.; Ueno, K.; Hatanaka, Y.; et al. Epidermal Growth Factor Receptor Tyrosine Kinase Defines Critical Prognostic Genes of Stage I Lung Adenocarcinoma. PLoS ONE 2012, 7, e43923. [Google Scholar] [CrossRef] [PubMed]
- Toro-Domínguez, D.; García, J.A.V.; Martorell-Marugán, J.; Román-Montoya, Y.; Alarcón-Riquelme, M.E.; Carmona-Sáez, P. A Survey of Gene Expression Meta-Analysis: Methods and Applications. Brief. Bioinform. 2020, 22, 1694–1705. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Stelzl, U.; Worm, U.; Lalowski, M.; Haenig, C.; Brembeck, F.H.; Goehler, H.; Stroedicke, M.; Zenkner, M.; Schoenherr, A.; Koeppen, S.; et al. A Human Protein-Protein Interaction Network: A Resource for Annotating the Proteome. Cell 2005, 122, 957–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Mering, C.; Huynen, M.; Jaeggi, D.; Schmidt, S.; Bork, P.; Snel, B. STRING: A Database of Predicted Functional Associations between Proteins. Nucleic Acids Res. 2003, 31, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.E.J. Modularity and Community Structure in Networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [Green Version]
- Bandettini, W.P.; Kellman, P.; Mancini, C.; Booker, O.J.; Vasu, S.; Leung, S.W.; Wilson, J.R.; Shanbhag, S.M.; Chen, M.Y.; Arai, A.E. MultiContrast Delayed Enhancement (MCODE) Improves Detection of Subendocardial Myocardial Infarction by Late Gadolinium Enhancement Cardiovascular Magnetic Resonance: A Clinical Validation Study. J. Cardiovasc. Magn. Reson. 2012, 14, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heard, N.A.; Rubin-Delanchy, P. Choosing between Methods of Combining p-Values. Biometrika 2018, 105, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Tseng, G.C. An Adaptively Weighted Statistic for Detecting Differential Gene Expression when Combining Multiple Transcriptomic Studies. Ann. Appl. Stat. 2011, 5, 994–1019. [Google Scholar] [CrossRef]
- Minna, J.D.; Roth, J.A.; Gazdar, A.F. Focus on Lung Cancer. Cancer Cell 2002, 1, 49–52. [Google Scholar] [CrossRef] [Green Version]
- Lukas, R.V.; Gondi, V.; Kamson, D.O.; Kumthekar, P.; Salgia, R. State-of-the-Art Considerations in Small Cell Lung Cancer Brain Metastases. Oncotarget 2017, 8, 71223–71233. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Ye, X.; Chen, R.; Gao, Q.; Zhao, D.; Ling, C.; Qian, Y.; Xu, C.; Tao, M.; Xie, Y. Sirtuin 7 Promotes Non-Small Cell Lung Cancer Progression by Facilitating G1/S Phase and Epithelial-Mesenchymal Transition and Activating AKT and ERK1/2 Signaling. Oncol. Rep. 2020, 44, 959–972. [Google Scholar] [CrossRef] [PubMed]
- Mangangcha, I.R.; Malik, M.Z.; Küçük, Ö.; Ali, S.; Singh, R.K.B. Kinless hubs are potential target genes in prostate cancer network. Genomics 2020, 112, 6. [Google Scholar]
- Zubbair, M.; Keilash, C.; Shahnawaz, A.; Romana, I.; Brojen, S. Methodology of predicting key regulators in ovarian cancer network: A network theoretical approach. BMC Cancer 2019, 19, 1129. [Google Scholar]
- Irengbam, R.M.; Zubbair, M.; Ömer, K.; Shakir, A.; Singh, R.K.B. Identification of key regulators in Prostate cancer from gene expression datasets of patients. Sci. Rep. 2019, 9, 16420. [Google Scholar]
- Zubbair, M.; Shahnawaz, A.; Singh, S.S.; Ishrat, R.; Singh, R.K.B. Dynamical states, possibilities and propagation of stress signal. Sci. Rep. 2017, 7, 40596. [Google Scholar]
- Malik, Z.; Alam, J.; Ishrat, R.; Agarwal, S.M.; Singh, R.K.B. Control of apoptosis by SMAR1. Mol. BioSyst. 2017, 13, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Shahnawaz, A.; Malik, Z.; Shyamchand, S.S.; Chirom, K.; Ishrat, R.; Singh, R.K.B. Exploring novel key regulators in breast cancer network. PLoS ONE 2018, 13, e0198525. [Google Scholar]
- Anam, F.; Safia, T.; Ahmed, M.M.; Alam, A.; Ali, S.; Malik, Z.; Ali, S.; Ishrat, R. Assessment of the key regulatory genes and their Enterology for Turner Syndrome employing network approach. Sci. Rep. 2018, 8, 10091. [Google Scholar]
- Aftab, A.; Nikhat, I.; Ahmed, M.M.; Tazyeen, S.; Tamkeen, N.; Farooqui, A.; Malik, Z. Romana Ishrat. Identification and Classification of Differentially Expressed Genes and Network Meta-Analysis Reveals Potential Molecular Signatures Associated with Tuberculosis. Front. Genet. 2019, 10, 932. [Google Scholar]
- Malumbres, M.; Barbacid, M. Cell Cycle, CDKs and Cancer: A Changing Paradigm. Nat. Rev. Cancer. 2009, 9, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Matera, R.; Saif, M.W. New Therapeutic Directions for Advanced Pancreatic Cancer: Cell Cycle Inhibitors, Stromal Modifiers and Conjugated Therapies. Expert Opin. Emerg. Drugs 2017, 22, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Brauer, C.; Thu, K.L.; Mason, J.M.; Blaser, H.; Bray, M.R.; Mak, T.W. Targeting Mitosis in Cancer: Emerging Strategies. Mol. Cell 2015, 60, 524–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.-C.; He, L.; Khoshaba, R.; Lu, F.-G.; Cai, C.; Zhou, F.-L.; Liao, D.-F.; Cao, D. Curcumin Nicotinate Selectively Induces Cancer Cell Apoptosis and Cycle Arrest through a P53-Mediated Mechanism. Molecules 2019, 24, 4179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Peng, R.; Sun, Y.; Wang, J.; Chong, X.; Zhang, Z. Identification of Key Genes in Non-Small Cell Lung Cancer by Bioinformatics Analysis. PeerJ 2019, 7, e8215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Kawakami, H.; Liu, W.; Zeng, X.; Strebhardt, K.; Tao, K.; Huang, S.; Sinicrope, F.A. Targeting CDK1 and MEK/ERK Overcomes Apoptotic Resistance in BRAF-Mutant Human Colorectal Cancer. Mol. Cancer Res. 2017, 16, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Gao, Y.; Ye, H.; Gerrin, S.; Ma, F.; Wu, Y.; Zhang, T.; Russo, J.; Cai, C.; Yuan, X.; et al. Positive Feedback Loop Mediated by Protein Phosphatase 1α Mobilization of P-TEFb and Basal CDK1 Drives Androgen Receptor in Prostate Cancer. Nucleic Acids Res. 2017, 45, 3738–3751. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Cho, H.; Shin, H.-Y.; Chung, J.-Y.; Kang, E.S.; Lee, E.-J.; Kim, J.-H. Accumulation of Cytoplasmic Cdk1 is Associated with Cancer Growth and Survival Rate in Epithelial Ovarian Cancer. Oncotarget 2016, 7, 49481–49497. [Google Scholar] [CrossRef] [Green Version]
- Kubo, H.; Suzuki, T.; Matsushima, T.; Ishihara, H.; Uchino, K.; Suzuki, S.; Tada, S.; Yoshimura, M.; Kondo, T. Cyclin-Dependent Kinase-Specific Activity Predicts the Prognosis of Stage I and Stage II Non-Small Cell Lung Cancer. BMC Cancer 2014, 14, 755. [Google Scholar] [CrossRef] [Green Version]
- Kuang, Y.; Guo, W.; Ling, J.; Xu, D.; Liao, Y.; Zhao, H.; Du, X.; Wang, H.; Xu, M.; Song, H.; et al. Iron-Dependent CDK1 Activity Promotes Lung Carcinogenesis Via Activation of the GP130/STAT3 Signaling Pathway. Cell Death Dis. 2019, 10, 297. [Google Scholar] [CrossRef]
- Shi, Y.-X.; Zhu, T.; Zou, T.; Zhuo, W.; Chen, Y.-X.; Huang, M.-S.; Zheng, W.; Wang, C.-J.; Li, X.; Mao, X.-Y.; et al. Prognostic and Predictive Values of CDK1 and MAD2L1 in Lung Adenocarcinoma. Oncotarget 2016, 7, 85235–85243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Liao, Y.; Long, D.; Yu, T.; Shen, F.; Lin, X. The Cdc2/Cdk1 Inhibitor, Purvalanol A, Enhances the Cytotoxic Effects of Taxol through Op18/Sathmin in Non-Small Cell Lung Cancer Cells in Vitro. Int. J. Mol. Med. 2017, 40, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Danilov, A.V.; Hu, S.; Orr, B.; Godek, K.; Mustachio, L.M.; Sekula, D.; Liu, X.; Kawakami, M.; Johnson, F.M.; Compton, D.A.; et al. Dinaciclib Induces Anaphase Catastrophe in Lung Cancer Cells via Inhibition of Cyclin-Dependent Kinases 1 and 2. Mol. Cancer Ther. 2016, 15, 2758–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquot, C.; Rousseau, B.; Carbonnelle, D.; Chinou, I.; Malleter, M.; Tomasoni, C.; Roussakis, C. Cucurbitacin-D-induced CDK1 mRNA Up-Regulation Causes Proliferation Arrest of a Non-Small Cell Lung Carcinoma Cell Line (NSCLC-N6). Anticancer Res. 2014, 34, 4797–4806. [Google Scholar] [PubMed]
- Li, M.; He, F.; Zhang, Z.; Xiang, Z.; Hu, D. CDK1 Serves as a Potential Prognostic Biomarker and Target for Lung Cancer. J. Int. Med. Res. 2020, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, M.I.G.; Floor, K.; Roepman, P.; Rodriguez, J.A.; Meijer, G.A.; Mooi, W.J.; Jassem, E.; Niklinski, J.; Muley, T.; van Zandwijk, N.; et al. Integration of Gene Dosage and Gene Expression in Non-Small Cell Lung Cancer, Identification of HSP90 as Potential Target. PLoS ONE 2008, 3, e0001722. [Google Scholar] [CrossRef]
- Workman, P.; Burrows, F.; Neckers, L.; Rosen, N. Drugging the Cancer Chaperone HSP90: Combinatorial Therapeutic Exploitation of Oncogene Addiction and Tumor Stress. Ann. N. Y. Acad. Sci. 2007, 1113, 202–216. [Google Scholar] [CrossRef]
- Sequist, L.V.; Gettinger, S.; Senzer, N.N.; Martins, R.G.; Jänne, P.A.; Lilenbaum, R.; Gray, J.E.; Iafrate, A.J.; Katayama, R.; Hafeez, N.; et al. Activity of IPI-504, a Novel Heat-Shock Protein 90 Inhibitor, in Patients with Molecularly Defined Non-Small-Cell Lung Cancer. J. Clin. Oncol. 2010, 28, 4953–4960. [Google Scholar] [CrossRef]
- Burrows, F.; Zhang, H.; Kamal, A. Hsp90 Activation and Cell Cycle Regulation. Cell Cycle 2004, 3, 1530–1536. [Google Scholar] [CrossRef] [Green Version]
- Kolosenko, I.; Grander, D.; Tamm, K. IL-6 Activated JAK/STAT3 Pathway and Sensitivity to Hsp90 Inhibitors in Multiple Myeloma. Curr. Med. Chem. 2014, 21, 3042–3047. [Google Scholar] [CrossRef]
- Patel, K.; Wen, J.; Magliocca, K.; Muller, S.; Liu, Y.; Chen, Z.G.; Saba, N.; Diaz, R. Heat Shock Protein 90 (HSP90) is Overexpressed in p16-Negative Oropharyngeal Squamous Cell Carcinoma, and Its inhibition in Vitro Potentiates the Effects of Chemoradiation. Cancer Chemother. Pharmacol. 2014, 74, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Liu, X.; Lou, J.; Han, X.; Zhang, L.; Wang, Q.; Li, B.; Dong, M.; Zhang, Y. Plasma Levels of Heat Shock Protein 90 Alpha Associated with Lung Cancer Development and Treatment Responses. Clin. Cancer Res. 2014, 20, 6016–6022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Chen, S.; Han, H.; Li, H.; Huang, Z.; Zhang, J.; Yin, Q.; Wang, X.; Ma, X.; Dai, P.; et al. Expression of Hsp90α and Cyclin B1 Were Related to Prognosis of Esophageal Squamous Cell Carcinoma and Keratin Pearl Formation. Int. J. Clin. Exp. Pathol. 2014, 7, 1544–1552. [Google Scholar] [PubMed]
- Tian, W.-L.; He, F.; Fu, X.; Lin, J.-T.; Tang, P.; Huang, Y.-M.; Guo, R.; Sun, L. High Expression of Heat Shock Protein 90 Alpha and its Significance in Human Acute Leukemia Cells. Gene 2014, 542, 122–128. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Pick, E.; Kluger, Y.; Gould-Rothberg, B.; Lazova, R.; Camp, R.L.; Rimm, D.L.; Kluger, H.M. HSP90 as a Marker of Progression in Melanoma. Ann. Oncol. 2007, 19, 590–594. [Google Scholar] [CrossRef]
- Záčková, M.; Moučková, D.; Lopotová, T.; Ondračková, Z.; Klamová, H.; Moravcová, J. Hsp90-a Potential Prognostic Marker in CML. Blood Cells Mol. Dis. 2013, 50, 184–189. [Google Scholar] [CrossRef]
- Calderwood, S.K.; Neckers, L. Hsp90 in Cancer: Transcriptional Roles in the Nucleus. Adv. Cancer Res. 2016, 129, 89–106. [Google Scholar]
- Zhang, B.; Wang, J.; Huang, Z.; Wei, P.; Liu, Y.; Hao, J.; Zhao, L.; Zhang, F.; Tu, Y.; Wei, T. Aberrantly Upregulated TRAP1 is Required for Tumorigenesis of Breast Cancer. Oncotarget 2015, 6, 44495–44508. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, C.; Mancini, M.; Benazzi, C.; Della Salda, L. Heat Shock Protein 90 is Associated with Hyperplasia and Neoplastic Transformation of Canine Prostatic Epithelial Cells. J. Comp. Pathol. 2014, 150, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Badowska-Kozakiewicz, A.M.; Malicka, E. Immunohistochemical Evaluation of Expression of Heat Shock Proteins HSP70 and HSP90 in Mammary Gland Neoplasms in Bitches. Pol. J. Veter.-Sci. 2012, 15, 209–214. [Google Scholar] [CrossRef]
- Lee, J.H.; Kang, K.W.; Kim, J.-E.; Hwang, S.W.; Park, J.H.; Kim, S.-H.; Ji, J.H.; Kim, T.G.; Nam, H.-Y.; Roh, M.S.; et al. Differential Expression of Heat Shock Protein 90 Isoforms in Small Cell Lung Cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 9487–9493. [Google Scholar] [PubMed]
- Wong, D.S.; Jay, D.G. Emerging Roles of Extracellular Hsp90 in Cancer. Adv. Cancer Res. 2016, 129, 141–163. [Google Scholar] [PubMed]
- Sims, J.D.; McCready, J.; Jay, D.G. Extracellular Heat Shock Protein (Hsp)70 and Hsp90α Assist in Matrix Metalloproteinase-2 Activation and Breast Cancer Cell Migration and Invasion. PLoS ONE 2011, 6, e18848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegelin, M.D. Inhibition of the mitochondrial Hsp90 chaperone network: A Novel, Efficient Treatment Strategy for Cancer? Cancer Lett. 2013, 333, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, A.; Sand, J.M.; Bauer, S.J.; Hafeez, B.B.; Meske, L.; Verma, A.K. Topically Applied Hsp90 Inhibitor 17AAG Inhibits UVR-Induced Cutaneous Squamous Cell Carcinomas. J. Investig. Dermatol. 2015, 135, 1098–1107. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.-F.; Xu, J.-H.; Gu, Y.-Y.; Pan, F.-M.; Tao, J.-H.; Wang, D.-G.; Xu, S.-Q.; Xiao, H.; Chen, P.-L.; Liu, S.; et al. Single Nucleotide Polymorphisms of HSP90AA1 Gene Influence Response of SLE Patients to Glucocorticoids Treatment. SpringerPlus 2016, 5, 222. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Q.; Chang, J.T.; Geradts, J.; Neckers, L.M.; Haystead, T.; Spector, N.L.; Lyerly, H.K. Amplification and High-Level Expression of Heat Shock Protein 90 Marks Aggressive Phenotypes of Human Epidermal Growth Factor Receptor 2 Negative Breast Cancer. Breast Cancer Res. 2012, 14, R62. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattacharyya, N.; Gupta, S.; Sharma, S.; Soni, A.; Bagabir, S.A.; Bhattacharyya, M.; Mukherjee, A.; Almalki, A.H.; Alkhanani, M.F.; Haque, S.; et al. CDK1 and HSP90AA1 Appear as the Novel Regulatory Genes in Non-Small Cell Lung Cancer: A Bioinformatics Approach. J. Pers. Med. 2022, 12, 393. https://doi.org/10.3390/jpm12030393
Bhattacharyya N, Gupta S, Sharma S, Soni A, Bagabir SA, Bhattacharyya M, Mukherjee A, Almalki AH, Alkhanani MF, Haque S, et al. CDK1 and HSP90AA1 Appear as the Novel Regulatory Genes in Non-Small Cell Lung Cancer: A Bioinformatics Approach. Journal of Personalized Medicine. 2022; 12(3):393. https://doi.org/10.3390/jpm12030393
Chicago/Turabian StyleBhattacharyya, Nirjhar, Samriddhi Gupta, Shubham Sharma, Aman Soni, Sali Abubaker Bagabir, Malini Bhattacharyya, Atreyee Mukherjee, Atiah H. Almalki, Mustfa F. Alkhanani, Shafiul Haque, and et al. 2022. "CDK1 and HSP90AA1 Appear as the Novel Regulatory Genes in Non-Small Cell Lung Cancer: A Bioinformatics Approach" Journal of Personalized Medicine 12, no. 3: 393. https://doi.org/10.3390/jpm12030393
APA StyleBhattacharyya, N., Gupta, S., Sharma, S., Soni, A., Bagabir, S. A., Bhattacharyya, M., Mukherjee, A., Almalki, A. H., Alkhanani, M. F., Haque, S., Ray, A. K., & Malik, M. Z. (2022). CDK1 and HSP90AA1 Appear as the Novel Regulatory Genes in Non-Small Cell Lung Cancer: A Bioinformatics Approach. Journal of Personalized Medicine, 12(3), 393. https://doi.org/10.3390/jpm12030393