1. Introduction
The selection of optimal oviposition sites is a key option to increase fitness, especially in animals who do not provide parental care to their offspring. Thus, this applies to odonates, in which oviposition site selection can significantly affect egg mortality [
1,
2]. The insertion of eggs into living or decaying plant tissues, referred to as endophytic oviposition, reduces water loss, minimizes winter mortality, and provides some protection from natural enemies [
3]. Most damselflies and some dragonflies (family Aeshnidae) use this endophytic oviposition strategy, whereas most other dragonfly species employ exophytic oviposition [
4,
5].
Endophytic oviposition is an ancient strategy for egg laying in Odonatoptera; it is documented from the Triassic, but its origin may be even older [
6,
7]. The endophytic ovipositor of damselflies consists of three pairs of valvulae to facilitate oviposition into substrates, such as plant tissues [
8,
9]. Most damselfly species use a wide range of host plants, but some species prefer particular plant species, or particular parts of plants, to lay their eggs in. Sensilla on the valves and styli may be involved in oviposition plant recognition [
10,
11,
12]. Female damselflies in the family Lestidae (e.g.,
Lestes spp.), in most cases, utilize plants that are submerged and often have relatively soft tissues; however, some species (e.g.,
Chalcolestes spp.) prefer to oviposit into stiff tissues, such as reeds or even tree bark [
9,
13]. Additionally, some other lestids (e.g.,
Sympecma spp.) oviposit into decaying plant material [
12,
14].
Based on current knowledge, Lestidae is the most diverse group of European damselflies that exhibit clutch patterning [
13]. Matushkina et al. [
9] showed that a high diversity in clutch patterning was related to the mechanical properties of the host plant and their availability. However, several species, such as
Chalcolestes viridis, prefer stiff substrates, despite the availability of softer plants (personal observation). In general, lestid females need more time to deposit an egg compared to other damselflies [
8,
13], which may result in higher mortality risk during oviposition [
15,
16]. However, they have a solid ovipositor and are able to exert the necessary force for oviposition into hard plant tissues [
8]. Thus, there is a trade-off between the cost of searching for oviposition sites, including mortality risk during oviposition, and the ability to hide/protect eggs. This is also an example of the traditional trade-off between quantity and quality (i.e., between offspring number and parental investment in their protection). In general, odonates with endophytic oviposition deposit less than ∼300 eggs at one oviposition event, while exophytic odonates often lay 1500 or more eggs in each clutch [
4,
5]. Egg mortality is widespread in aquatic insects; therefore, females developed various strategies to defend their eggs against predators. The efficiency of such defense is related to the search capabilities of the predator; therefore, oviposition in hard-to-access or cryptic locations may be an effective strategy. Eggs may be defended physically and chemically [
17].
It is known that egg parasitoids are able to use the mixtures of plant volatiles (semiochemicals induced during host oviposition) and host odour for host localization [
18,
19]. Significantly more eggs are parasitized when they are clumped [
20,
21]. Additionally, they are significantly less parasitized when they are oviposited on habitat edges than the eggs oviposited in the center of the habitat [
21]. This indicates that investment in oviposition site selection may greatly affect offspring fitness. Freshwater insects have a wide range of oviposition strategies to reduce egg detection by egg parasitoids [
22,
23,
24]. As already mentioned, the distribution of lestid eggs is not random: females show a clear preference for certain host plants and even for specific microhabitats, and eggs are laid in multi-egg clumps [
12,
25]. However, very little is known about how females’ preference for specific oviposition substrates influences the likelihood of egg mortality. In this study, we compared the proportion of parasitized eggs of different lestid species with different oviposition strategies, focusing on their preference for specific oviposition substrate (i.e., plant species).
We evaluated two main hypotheses:
There are differences in protection against egg parasitoids between different study species. Preference for stiff substrate in C. viridis can provide significant protection from egg parasitoids.
There are differences in overall egg mortality rates between individual damselfly species.
3. Results
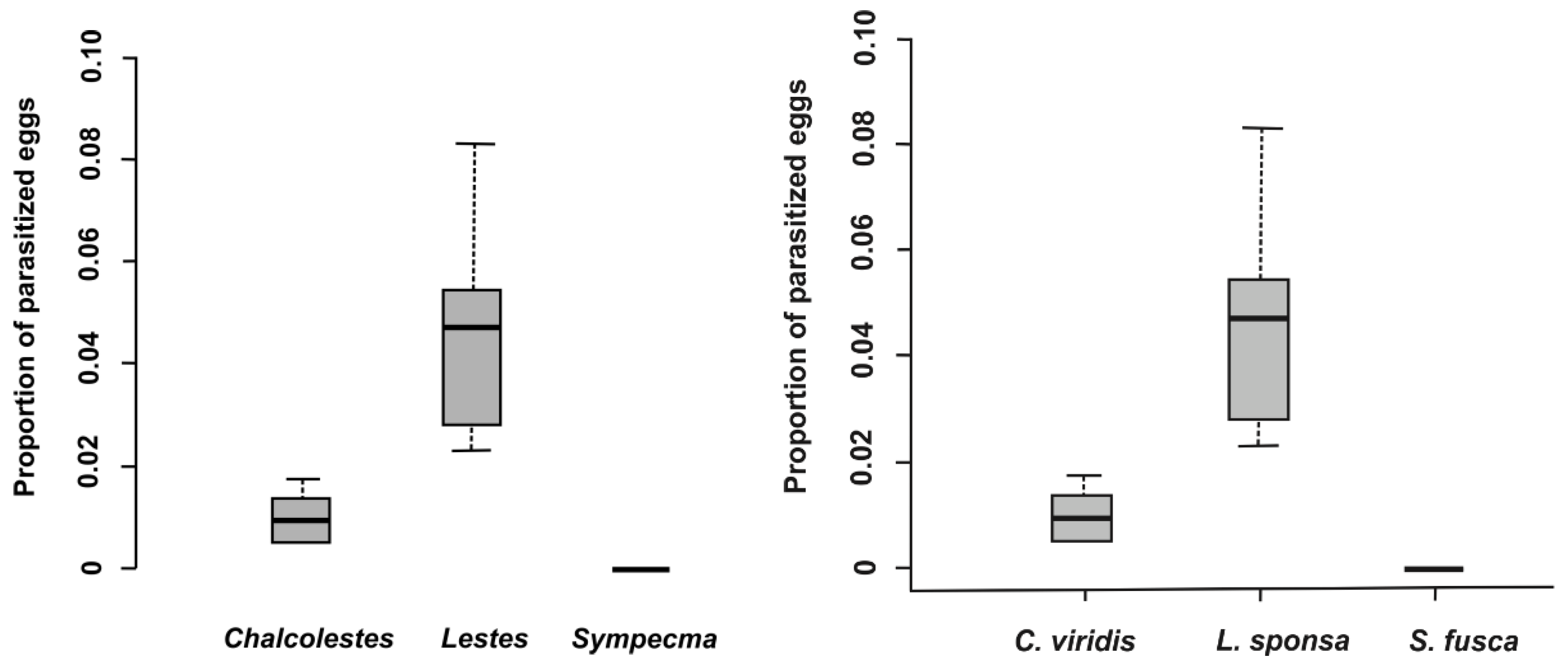
In total, 28,098 eggs of all lestid species were extracted from plant tissues (
Table 3). There was a significantly higher proportion of parasitized eggs for
L. sponsa (4.7%) than for
C. viridis (1.0%), whereas we did not find any parasitoids in the eggs of
S. fusca (
Table 3 and
Table 4,
Figure 1) while the proportion of parasitized eggs was not affected by oviposition plant (
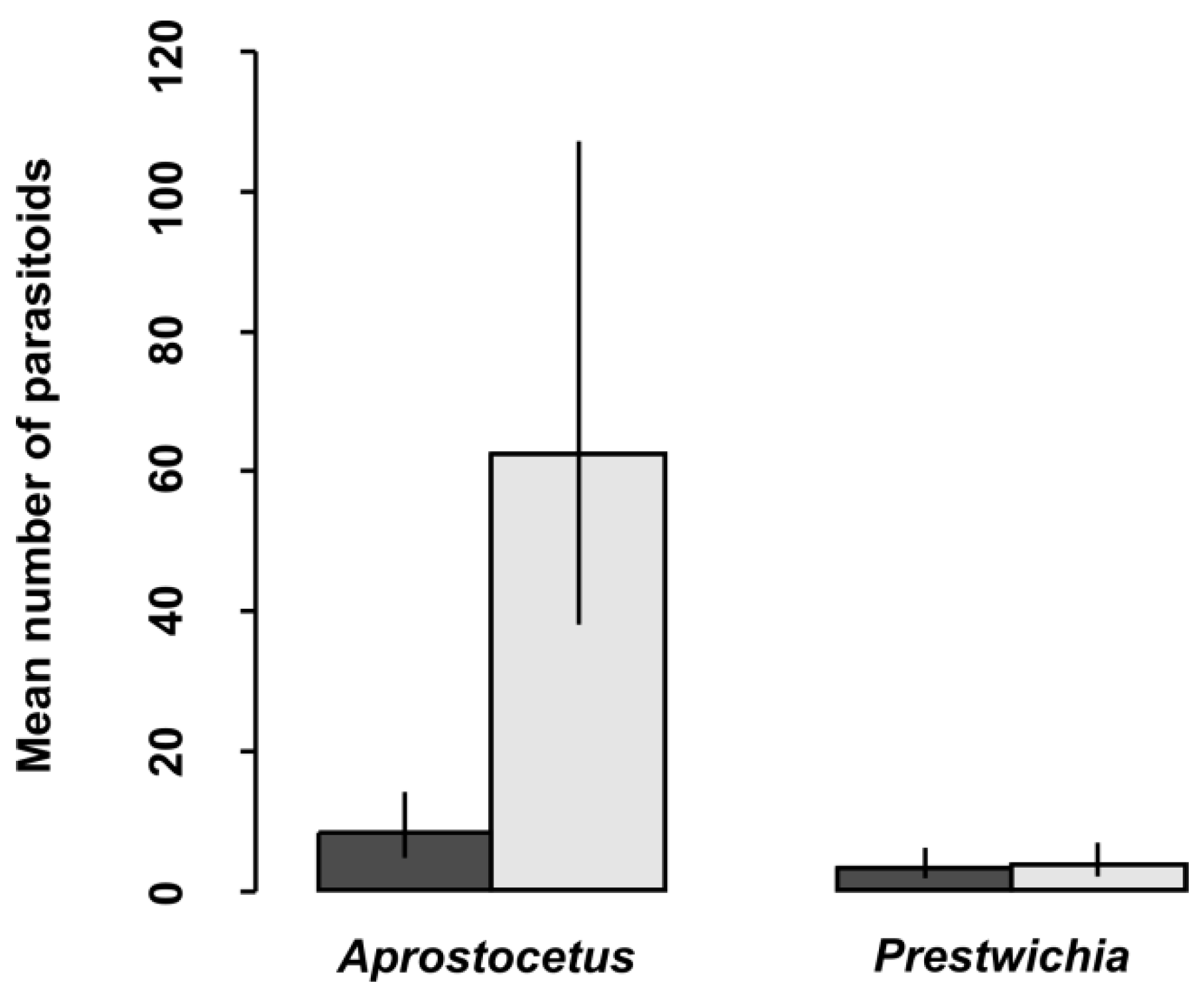
Table 4). We also looked at the proportion and abundance of individual species of parasitoids. Interestingly, parasitoid species differed significantly between eggs of damselfly species (
Table 5,
Figure 2). According to our results, the prevalence of parasitoids from the genus
Aprostocetus was significantly higher in
L. sponsa than in
C. viridis, whereas no such trend was found for
Prestwichia aquatica (
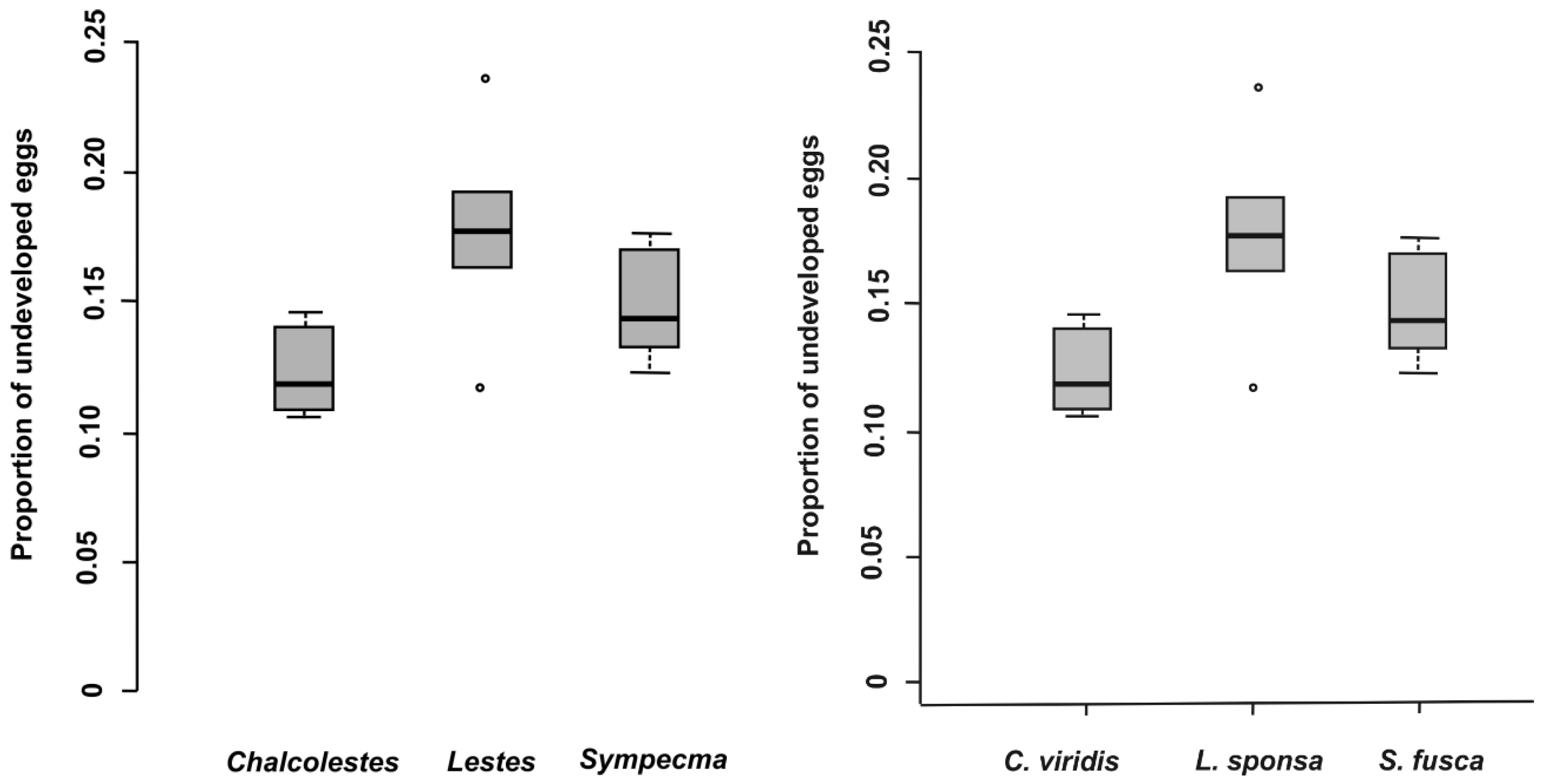
Figure 2). The proportion of undeveloped eggs significantly varied across species and was not affected by oviposition plant (
Figure 3,
Table 6). The capability of
C. viridis to oviposit into stiff tissues, outside the aquatic habitat, provided other additional benefits, such as a lower proportion of undeveloped eggs compared to the other species. A significantly higher proportion of undeveloped eggs was found in
L. sponsa (
Figure 3).
4. Discussion
Species from the family Lestidae represent damselflies with very diverse egg-laying strategies, differing in clutch size, host plant, and microclimate [
13]. Based on our findings, it is evident that eggs of species with different oviposition preferences have a significantly different survival outcome. It seems that the higher costs associated with some oviposition strategies could be considered as a trade-off between investment into costly oviposition strategy and parasitoid avoidance.
The level of parasitoid protection in eggs laid into soft plant tissues in
L. sponsa was lower than that in
C. viridis eggs oviposited into stiff plants tissues. This indicates that the ability of
C. viridis to oviposit into substrates such as tree bark provides a defense for the eggs and increases the chances of offspring survival, whereas the
L. sponsa eggs deposited into the soft-stemmed herbs are more exposed to enemies. Species that invest heavily in egg defense are unable to lay as many eggs as species that do not protect their eggs [
34] like our study species.
A significantly lower proportion of undeveloped eggs in
C. viridis may be due to the fact that their overall oviposition rate is very low compared to other damselflies [
13]. Females produce larger clusters of eggs, which is a “logical choice” due to the stiffness of their preferred oviposition substrates. However, prolonged oviposition means that
C. viridis females are more susceptible to predation [
5]. In this respect, oviposition of
S. fusca into soft plant tissues may be the most advantageous strategy. Oviposition rates of
S. fusca are significantly higher than in other lestids (except
L. barbarus) [
13] and have a relatively low proportion of undeveloped eggs. Another potential advantage may be the fact that eggs placed in floating dead plant tissues are very difficult for non-aquatic predators to reach. Furthermore, dead plants do not produce semiochemicals, which can be used for egg detection. This may be an explanation why
S. fusca eggs were completely free of parasitoids. However, dead plant tissues have only a short-term durability. Therefore, eggs placed in this substrate must hatch promptly after oviposition and may not survive a long overwintering period. Thus, dead plant material, as an oviposition substrate, would be suitable for a limited number of species, notably those who will not overwinter in plant tissues. Specific life history of damselflies in the genus
Sympecma includes several unique adaptations, including adult overwintering and rapid larval development [
14]. The whole complex of adaptations yields benefits by reducing competition for both larvae and adults [
35]. According to our results, phenological shift, obtained through adult overwintering, may be another reason why parasitoids were absent from
S. fusca eggs. During coevolution, parasitoids with short-lived adult life stages tend to focus their development to a period when sufficient hosts (eggs) are available [
36]. Accordingly, there could be an enemy-free space for species with fundamentally different phenology, such as
S. fusca.
Unique adaptations of the genus
Sympecma, including phenological shift, evolved in extreme arid conditions [
14,
37]. Other lestid species must protect their eggs against parasitoids. Based on our findings, the most common parasitoid species
Aprostocetus (
Ootetrastichus)
pseudopodiellus was significantly less abundant in
C. viridis eggs than in those of
L. sponsa. This indicates that at least several parasitoid species are somehow limited in their ability to detect and/or attack
C. viridis eggs. We assume this is because
C. viridis eggs, which are generally oviposited into stiff tissues, are better protected against predation. Another possible explanation is that
C. viridis tends to oviposit into the bark of trees, which are usually several meters away from the aquatic habitat, and can thus be at the edge of interest for parasitoids looking for aquatic hosts. This concurs with Cronin et al. [
21], who have found significantly fewer parasitoids at the habitat edges.
Conversely, while oviposition away from the aquatic habitat may be beneficial for eggs, it could pose an insurmountable problem for larvae. Immediately after hatching, larvae must find water. Therefore, in only several odonate species, such as
C. viridis, the first instar is a “true prolarva” that can reach the water by jumping or springing [
5]. In most odonate species, the prolarval stadium is extensively reduced and lasts less than one minute [
4,
38]. The mortality of prolarvae that must cross land to reach water is probably high, and therefore should not be disregarded in estimates of the overall survival of individual species [
5].
5. Conclusions
In conclusion, our findings show that egg mortality varies significantly between species. According to our results, it is obvious that the ability to lay eggs into plant substrates that are somewhat unsuitable for oviposition (e.g., stiff or decaying plant tissues), and thus are not used as often, may serve as an efficient strategy against some parasitoid groups. Although the mechanisms of the egg search in odonate parasitoids are poorly known, our results suggest that oviposition into different substrates may elicit different chemical reaction, which may consequently affect the ability to detect host eggs. Moreover, a suitable oviposition substrate can significantly enhance survival during overwintering, by mitigating the effect of some lethal environmental processes (e.g., the formation of ice crystals or desiccation). Broadly, however, the utilization of a particular substrate is not only related to the mechanical properties of the plant tissues and female oviposition capabilities, but also to other adaptations, such as prolonged prolarva in C. viridis or adult overwintering in S. fusca.






{kind=link}
{kind=link}
{kind=link}