Laboratory and Greenhouse Evaluation of a Granular Formulation of Beauveria bassiana for Control of Western Flower Thrips, Frankliniella occidentalis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rearing Protocols for WFT
2.2. Fungal Strains and Culture
2.3. Production of Beauveria bassiana Granules
2.4. Pathogenicity Assay of Beauveria bassiana Granules
2.5. Micromorphological Observations of Fungal Infection
2.6. Colonization of Soil by Beauveria bassiana under Different Soil Moisture Conditions
2.7. Greenhouse Experiments
2.8. Data Analysis
3. Results
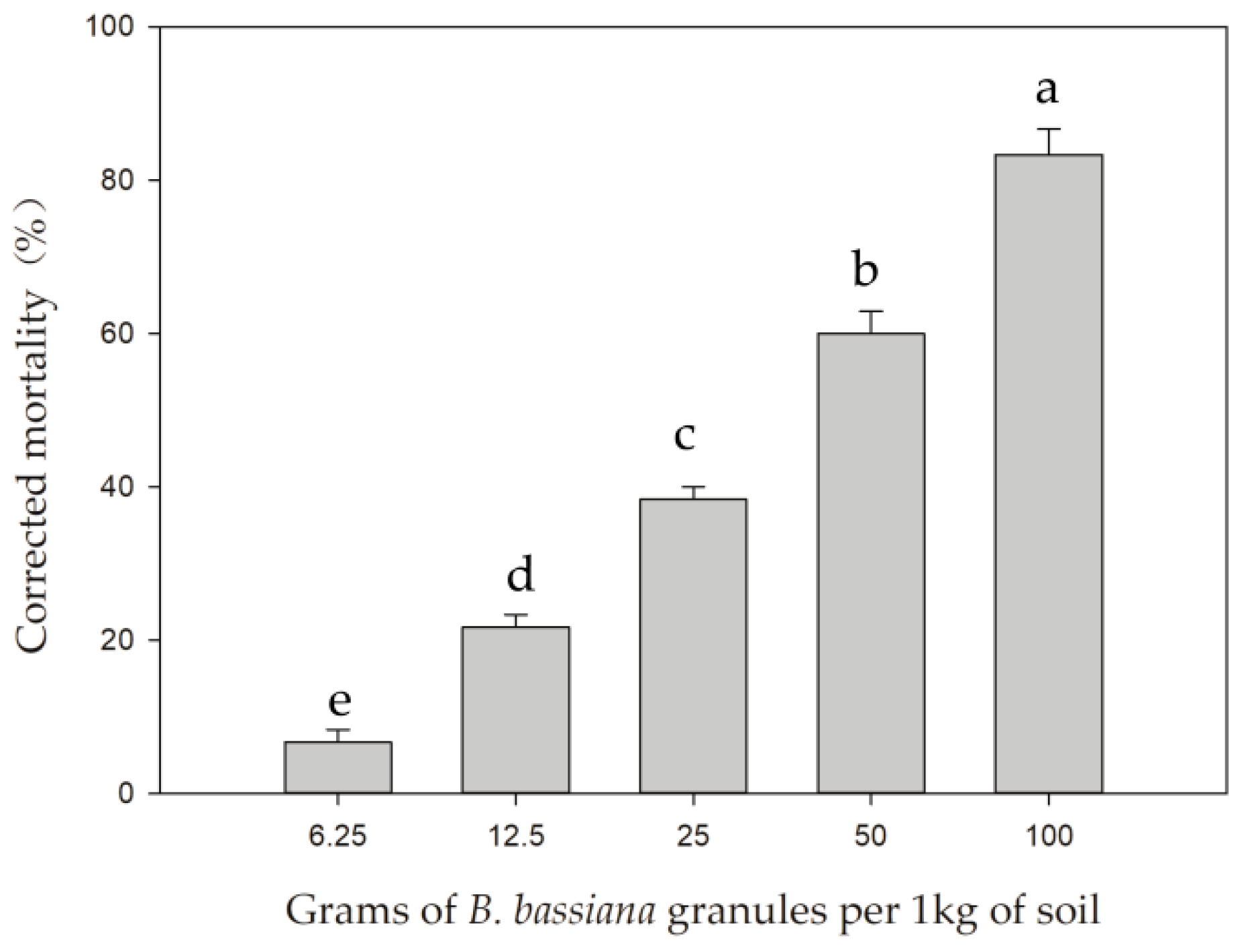
3.1. Pathogenicity of Beauveria bassiana Granules to Frankliniella occidentalis
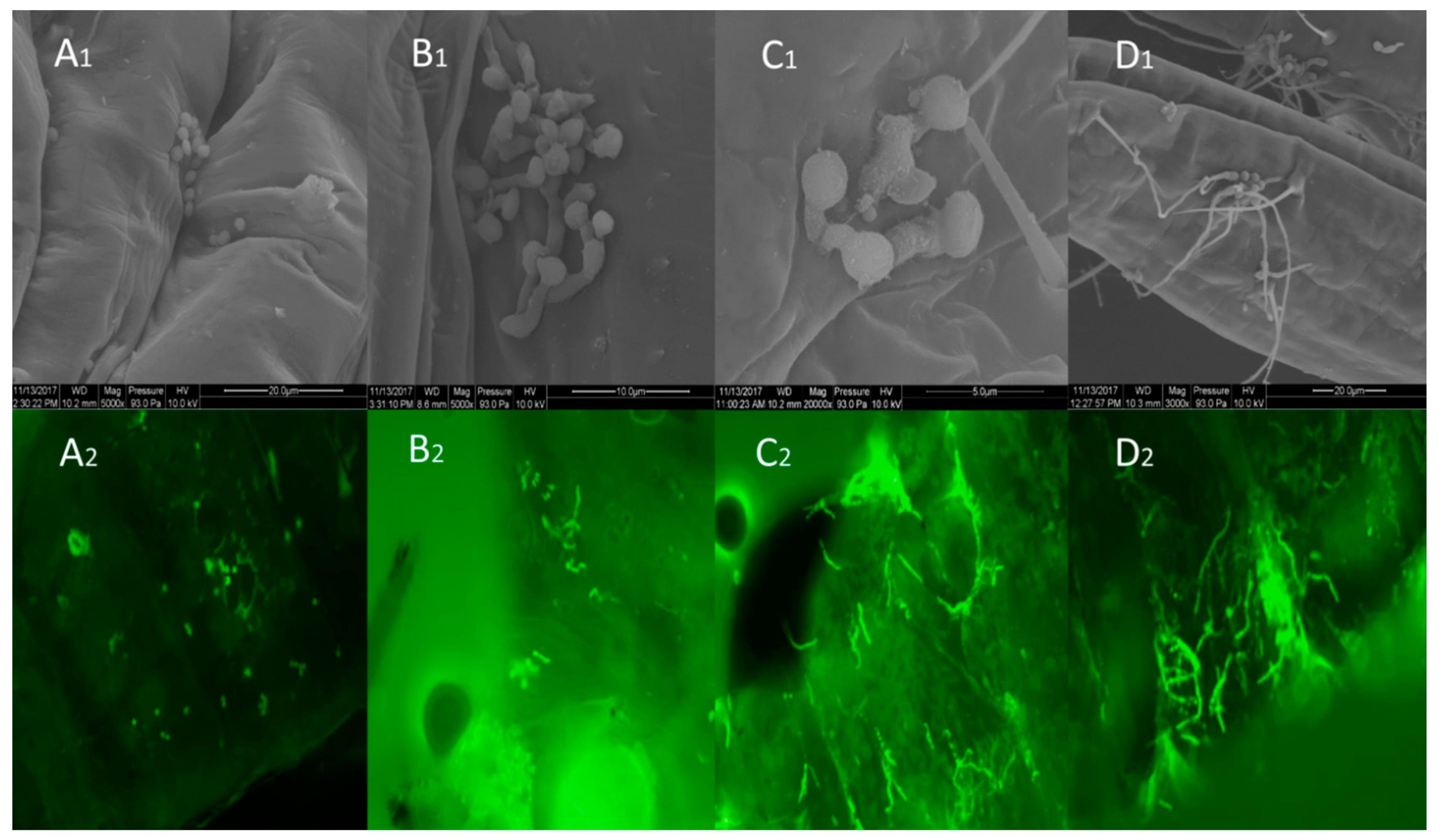
3.2. Micromorphological Observations of Fungal Infection
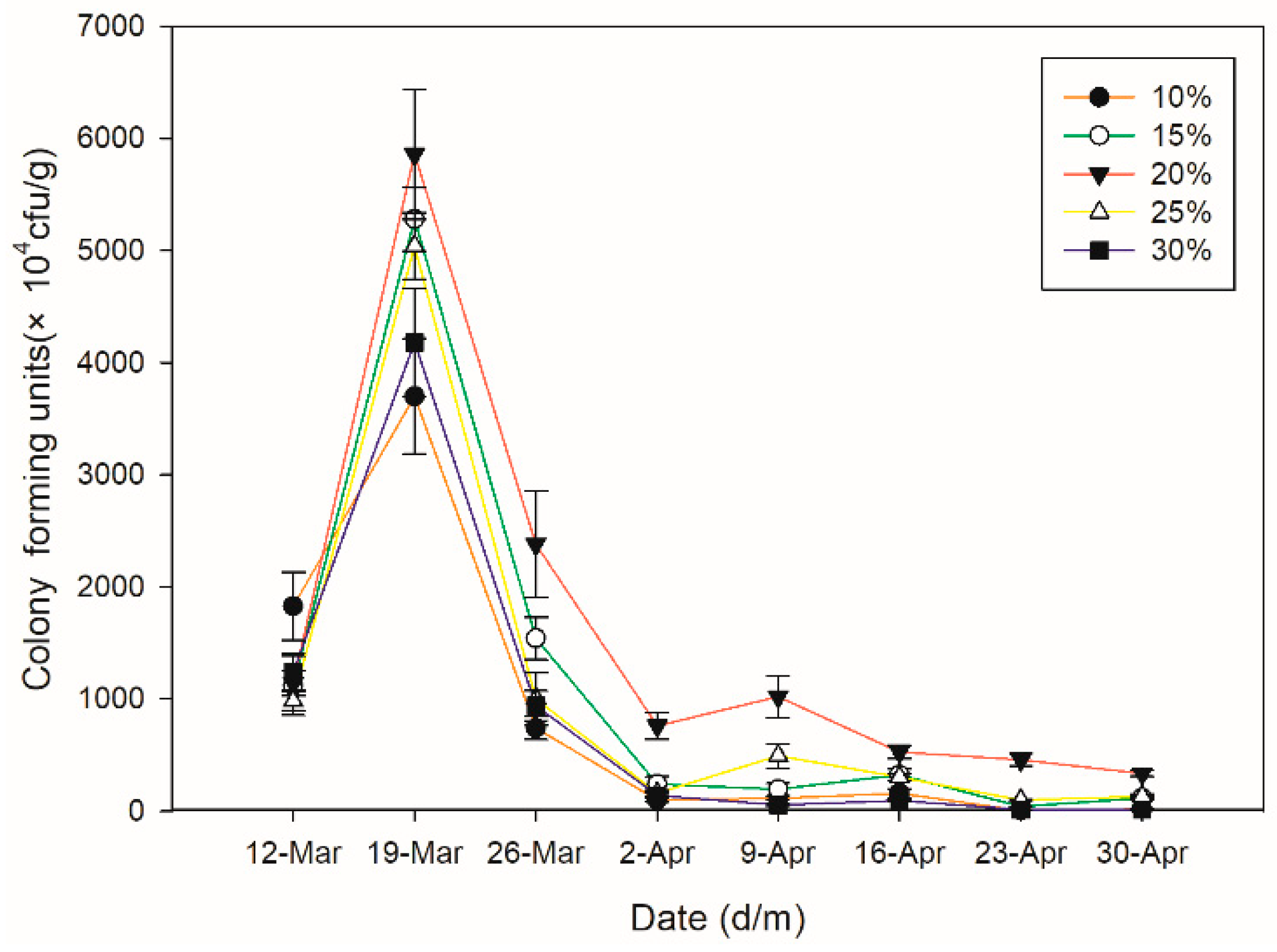
3.3. Colonization of Beauveria bassiana under Different Soil Water Concentrations
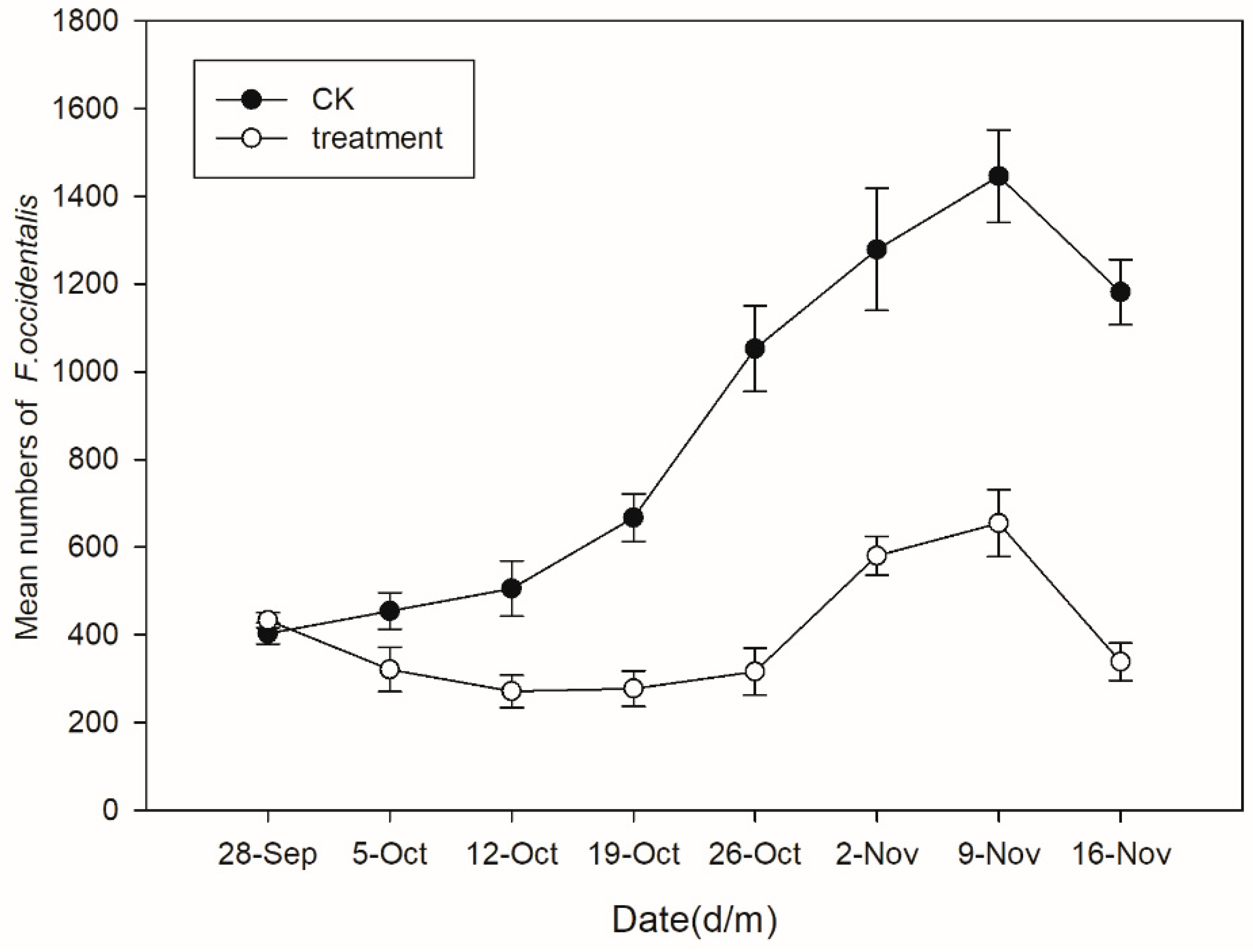
3.4. Efficacy of Soil-Applied Beauveria bassiana Granules against Frankliniella occidentalis in a Greenhouse
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reitz, S.R. Biology and ecology of the western flower thrips (Thysanoptera: Thripidae): The making of a pest. Fla. Entomol. 2009, 92, 7–13. [Google Scholar] [CrossRef]
- Mouden, S.; Sarmiento, K.F.; Klinkhamer, P.G.; Leiss, K.A. Integrated pest management in western flower thrips: Past, present and future. Pest Manag. Sci. 2017, 73, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.L.; Lei, Z.R.; Reitz, S.R. Western flower thrips resistance to insecticides: Detection, mechanisms and management strategies. Pest Manag. Sci. 2012, 68, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Cloyd, R.A. Western flower thrips (Thysanoptera: Thripidae) and insecticide resistance: An overview and strategies to mitigate insecticide resistance development. J. Entomol. Sci. 2016, 51, 257–273. [Google Scholar] [CrossRef]
- Moritz, G.; Kumm, S.; Mound, L. Tospovirus transmission depends on thrips ontogeny. Virus Res. 2004, 99, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Robb, K.L. Analysis of Frankliniella occidentalis (Pergande) as a Pest of Floricultural Crops in California Greenhouse. Ph.D. Dissertation, University of California, San Diego, CA, USA, 1989; 135p. [Google Scholar]
- Pappu, H.R.; Jones, R.A.C.; Jain, R.K. Global status of Tospovirus epidemics in diverse cropping systems: Successes achieved and challenges ahead. Virus Res. 2009, 141, 219–236. [Google Scholar] [CrossRef]
- Webster, C.G.; Reitz, S.R.; Perry, K.L.; Adkins, S. A natural M RNA reassortant arising from two species of plant-and insect-infecting Bunyaviruses and comparison of its sequence and biological properties to parental species. Virology 2011, 413, 216–225. [Google Scholar] [CrossRef]
- Steiner, M.Y.; Spohr, L.J.; Goodwin, S. Relative humidity controls pupation success and dropping behaviour of western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Aust. J. Entomol. 2011, 50, 179–186. [Google Scholar] [CrossRef]
- Holmes, N.; Bennison, J.; Maulden, K.; Kirk, W. The pupation behaviour of the western flower thrips, Frankliniella occidentalis (Pergande). Acta Phytopathol. Entomol. Hung. 2012, 47, 87–96. [Google Scholar] [CrossRef]
- Skinner, M.; Gouli, S.; Frank, C.E.; Parker, B.L.; Kim, J.S. Management of Frankliniella occidentalis (Thysanoptera: Thripidae) with granular formulations of entomopathogenic fungi. Biol. Control. 2012, 63, 246–252. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ugine, T.A.; Ramos, M.E.; Sanderson, J.P. Efficacy of spray applications of entomopathogenic fungi against western flower thrips infesting greenhouse impatiens under variable moisture conditions. Biol. Control 2016, 97, 31–47. [Google Scholar] [CrossRef] [Green Version]
- Nault, B.A.; Shelton, A.M. Impact of insecticide efficacy on developing action thresholds for pest management: A case study of onion thrips (Thysanoptera: Thripidae) on onion. J. Econ. Entomol. 2010, 103, 1315–1326. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Reitz, S.R.; Yuan, H.G.; Lei, Z.R.; Paini, D.R.; Gao, Y.L. Pesticide-mediated interspecific competition between local and invasive thrips pests. Sci. Rep. 2017, 7, 40512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Reitz, S.R.; Wang, H.H.; Lei, Z.R. Sublethal effects of Beauveria bassiana (Ascomycota: Hypocreales) on life table parameters of Frankliniella occidentalis (Thysanoptera: Thripidae). J. Econ. Entomol. 2015, 108, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Reitz, S.R.; Yearby, E.L.; Funderburk, J.E.; Stavisky, J.; Momol, M.T.; Olson, S.M. Integrated management tactics for Frankliniella thrips (Thysanoptera: Thripidae) in field-grown pepper. J. Econ. Entomol. 2003, 96, 1201–1214. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Zhang, Z.K.; Gao, Y.L.; Xu, X.N.; Lei, Z.R. Interactions between foliage- and soil-dwelling predatory mites and consequences for biological control of Frankliniella occidentalis. Biocontrol 2016, 61, 717–727. [Google Scholar] [CrossRef]
- Shah, P.A.; Pell, J.K. Entomopathogenic fungi as biological control agents. Appl. Microbiol. Biotechnol. 2003, 61, 413–423. [Google Scholar] [CrossRef] [PubMed]
- de Faria, M.R.; Wright, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Ludwig, S.W.; Oetting, R.D. Efficacy of Beauveria bassiana plus insect attractants for enhanced control of Frankliniella occidentalis (Thysanoptera: Thripidae). Fla. Entomol. 2002, 85, 270–272. [Google Scholar] [CrossRef]
- Jacobson, R.; Chandler, D.; Fenlon, J.; Russell, K. Compatibility of Beauveria bassiana (Balsamo) Vuillemin with Amblyseius cucumeris Oudemans (Acarina: Phytoseiidae) to control Frankliniella occidentalis Pergande (Thysanoptera: Thripidae) on cucumber plants. Biocontrol Sci. Technol. 2001, 11, 391–400. [Google Scholar] [CrossRef]
- Ansari, M.; Brownbridge, M.; Shah, F.; Butt, T. Efficacy of entomopathogenic fungi against soil-dwelling life stages of western flower thrips, Frankliniella occidentalis, in plant-growing media. Entomol. Exp. Appl. 2008, 127, 80–87. [Google Scholar] [CrossRef]
- Wu, S.Y.; Zhen, H.; Wang, E.D.; Xu, X.N.; Lei, Z.R. Application of Beauveria bassiana and Neoseiulus barkeri for improved control of Frankliniella occidentalis in greenhouse cucumber. Crop Prot. 2017, 96, 83–87. [Google Scholar] [CrossRef]
- Wu, S.Y.; Gao, Y.L.; Xu, X.N.; Zhang, Y.P.; Wang, J.; Lei, Z.R.; Smagghe, G. Laboratory and greenhouse evaluation of a new entomopathogenic strain of Beauveria bassianafor control of the onion thrips Thrips tabaci. Biocontrol Sci. Technol. 2013, 23, 794–802. [Google Scholar] [CrossRef]
- Ugine, T.A.; Wraight, S.P.; Sanderson, J.P. Influences of impatiens pollen and exposure to Beauveria bassiana on bionomics of western flower thrips Frankliniella occidentalis. Biol. Control 2006, 37, 186–195. [Google Scholar] [CrossRef]
- Jackson, M.A.; Jaronski, S.T. Production of microsclerotia of the fungal entomopathogen Metarhizium anisopliae and their potential for use as a biocontrol agent for soil-inhabiting insects. Mycol. Res. 2009, 113, 842–850. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, S.J.; Skinner, M.; Parker, B.L. A novel approach: Beauveria bassiana granules applied to nursery soil for management of rice water weevils in paddy fields. Pest Manag. Sci. 2014, 70, 1186–1191. [Google Scholar] [CrossRef]
- Goettel, M.S.; Inglis, G.D. Fungi: Hyphomycetes. In Manual of Techniques in Insect Pathology; Elsevier: Amsterdam, The Netherlands, 1997; pp. 213–249. [Google Scholar]
- Li, Y.P.; Lei, Z.R.; Wang, H.H. Selection of Beauveria bassiana strains against Frankliniella occidentalis and their conidial production characteristics. Chin. J. Biol. Control 2013, 29, 219–226. [Google Scholar]
- Wu, S.Y.; Gao, Y.L.; Smagghe, G.; Xu, X.N.; Lei, Z.R. Interactions between the entomopathogenic fungus Beauveria bassiana and the predatory mite Neoseiulus barkeri and biological control of their shared prey/host Frankliniella occidentalis. Biol. Control 2016, 98, 43–51. [Google Scholar] [CrossRef]
- Li, Y.L.; Wang, Y.Y.; Dong, J.Z.; Zhou, T.; Li, J. The effects on survival ability of Beauveria bassiana under soil microecological environment. Chin. Agric. Sci. Bull. 2012, 36, 4–8. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Wu, S.Y.; Gao, Y.L.; Zhang, Y.P.; Wang, E.D.; Xu, X.N.; Lei, Z.R. An entomopathogenic strain of Beauveria bassiana against Frankliniella occidentalis with no detrimental effect on the predatory mite Neoseiulus barkeri: Evidence from laboratory bioassay and scanning electron microscopic observation. PLoS ONE 2014, 9, e84732. [Google Scholar] [CrossRef] [PubMed]
- Holder, D.J.; Keyhani, N.O. Adhesion of the entomopathogenic fungus Beauveria (Cordyceps) bassiana to substrata. Appl. Environ. Microbiol. 2005, 71, 5260–5266. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.J.; Lei, Z.R.; Wang, S.Y.; Wang, H.H. A new fluorescent microscopy method for identifying Beauveria bassiana infected Bemisia tabaci nymphs. Chin. J. Appl. Entomol. 2015, 52, 267–271. [Google Scholar]
- Charnley, A.; Leger, R.S. The role of cuticle-degrading enzymes in fungal pathogenesis in insects. In The Fungal Spore and Disease Initiation in Plants and Animals; Springer: Berlin/Heidelberg, Germany, 1991; pp. 267–286. [Google Scholar]
- Gillespie, J.P.; Bailey, A.M.; Cobb, B.; Vilcinskas, A. Fungi as elicitors of insect immune responses. Arch. Insect Biochem. 2000, 44, 49–68. [Google Scholar] [CrossRef]
- Fargues, J.; Luz, C. Effects of fluctuating moisture and temperature regimes on the infection potential of Beauveria bassiana for Rhodnius prolixus. J. Invertebr. Pathol. 2000, 75, 202–211. [Google Scholar] [CrossRef]
- Rangel, D.E.N.; Anderson, A.J.; Roberts, D.W. Evaluating physical and nutritional stress during mycelial growth as inducers of tolerance to heat and UV-B radiation in Metarhizium anisopliae conidia. Mycoll. Res. 2008, 112, 1362–1372. [Google Scholar] [CrossRef] [PubMed]
- Hallsworth, J.E.; Magan, N. Water and temperature relations of growth of the entomogenous fungi Beauveria bassiana, Metarhizium anisopliae, and Paecilomyces farinosus. J. Invertebr. Pathol. 1999, 74, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Fuxa, J.R.; Richter, A.R. Effects of soil moisture and composition and fungal isolate on prevalence of Beauveria bassiana in laboratory colonies of the red imported fire ant (Hymenoptera: Formicidae). Environ. Entomol. 2004, 33, 975–981. [Google Scholar] [CrossRef]
- Shipp, J.L.; Zhang, Y.; Hunt, D.W.A.; Ferguson, G. Influence of humidity and greenhouse microclimate on the efficacy of Beauveria bassiana (Balsamo) for control of greenhouse arthropod pests. Environ. Entomol. 2003, 32, 1154–1163. [Google Scholar] [CrossRef]
- Saito, T.; Brownbridge, M. Compatibility of soil-dwelling predators and microbial agents and their efficacy in controlling soil-dwelling stages of western flower thrips Frankliniella occidentalis. Biol. Control 2016, 92, 92–100. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Lei, Z.; Reitz, S.R.; Wu, S.; Gao, Y. Laboratory and Greenhouse Evaluation of a Granular Formulation of Beauveria bassiana for Control of Western Flower Thrips, Frankliniella occidentalis. Insects 2019, 10, 58. https://doi.org/10.3390/insects10020058
Zhang X, Lei Z, Reitz SR, Wu S, Gao Y. Laboratory and Greenhouse Evaluation of a Granular Formulation of Beauveria bassiana for Control of Western Flower Thrips, Frankliniella occidentalis. Insects. 2019; 10(2):58. https://doi.org/10.3390/insects10020058
Chicago/Turabian StyleZhang, Xingrui, Zhongren Lei, Stuart R. Reitz, Shengyong Wu, and Yulin Gao. 2019. "Laboratory and Greenhouse Evaluation of a Granular Formulation of Beauveria bassiana for Control of Western Flower Thrips, Frankliniella occidentalis" Insects 10, no. 2: 58. https://doi.org/10.3390/insects10020058
APA StyleZhang, X., Lei, Z., Reitz, S. R., Wu, S., & Gao, Y. (2019). Laboratory and Greenhouse Evaluation of a Granular Formulation of Beauveria bassiana for Control of Western Flower Thrips, Frankliniella occidentalis. Insects, 10(2), 58. https://doi.org/10.3390/insects10020058