Ability of Muscidifurax raptorellus and Other Parasitoids and Predators to Control Drosophila suzukii Populations in Raspberries in the Laboratory

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Sources
2.2. Parasitoid and Predator Experiment with Multiple D. suzukii Life Stages
2.3. Predator Experiment with Single D. suzukii Life Stage
2.4. Data Analysis
3. Results
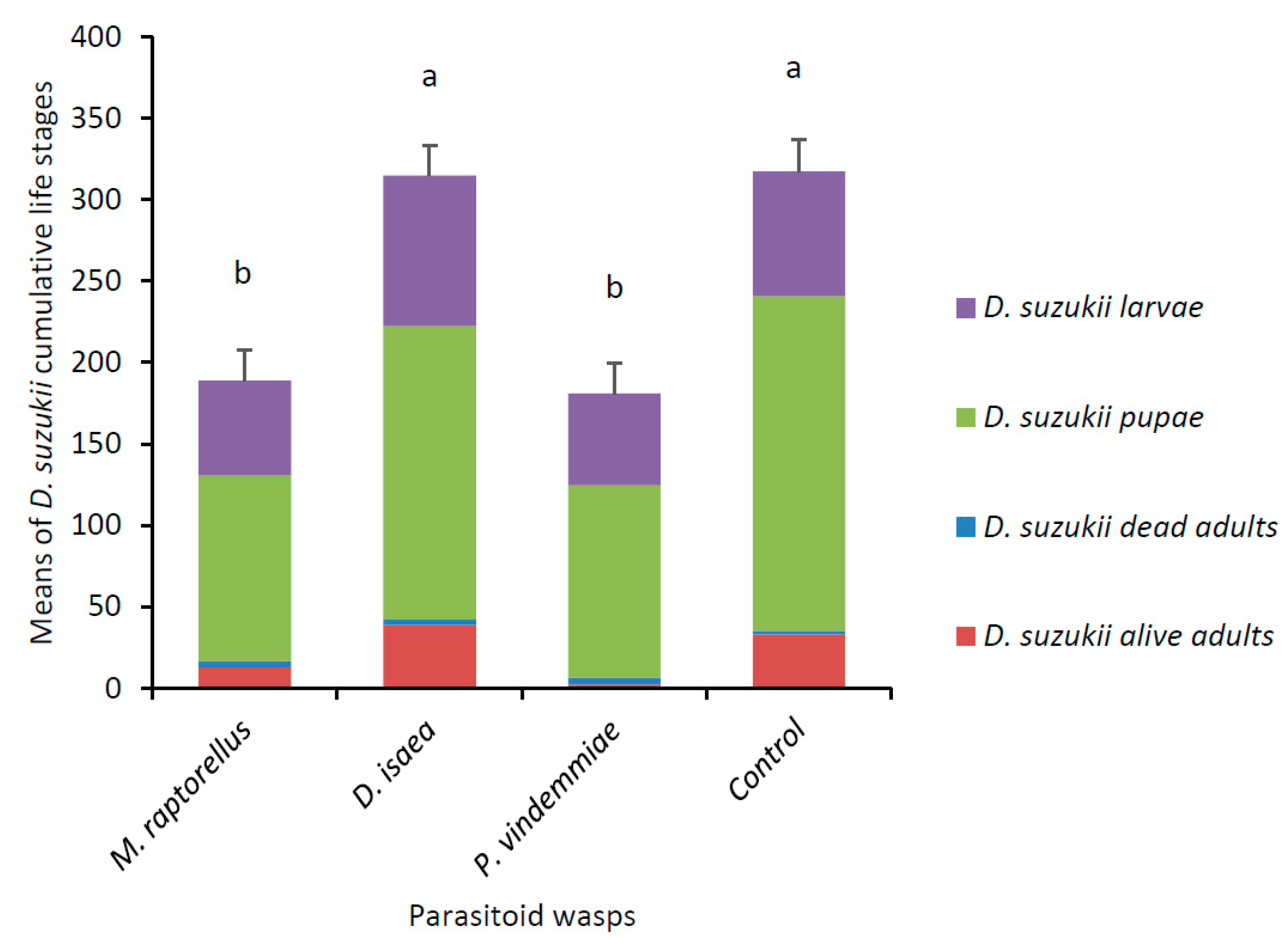
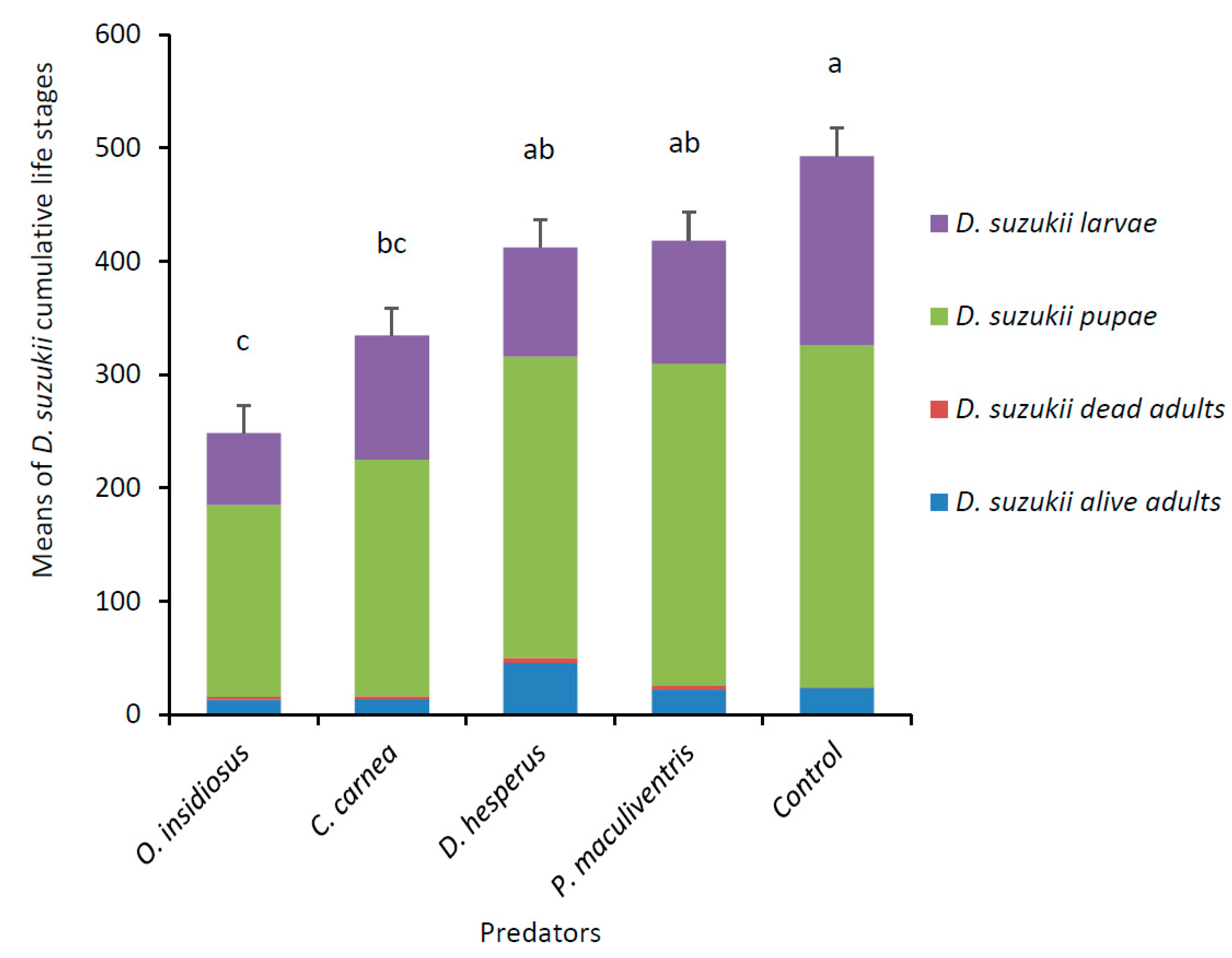
3.1. Parasitoid and Predator Experiment with Multiple D. suzukii Life Stages
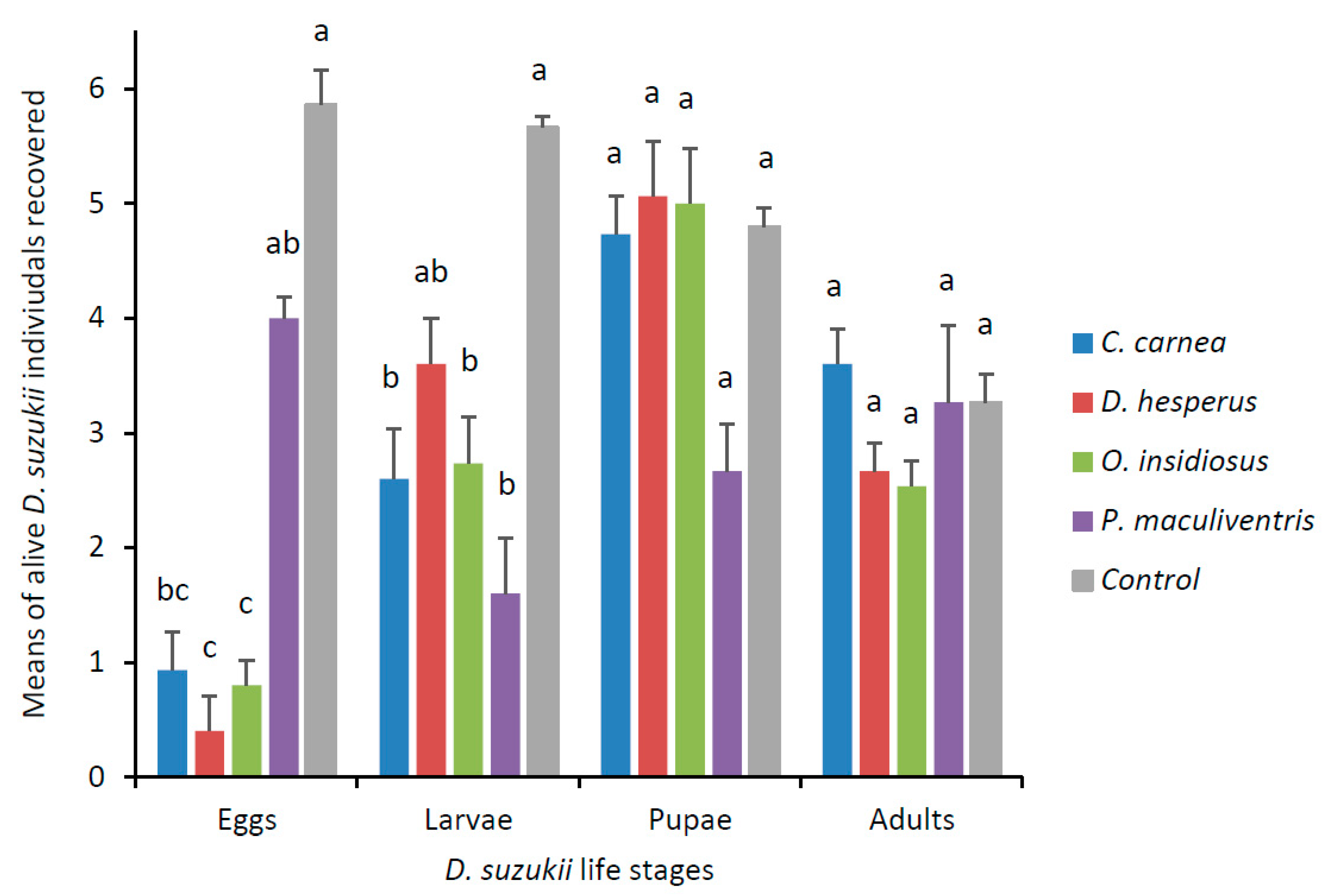
3.2. Predator Experiment with Single D. suzukii Life Stage
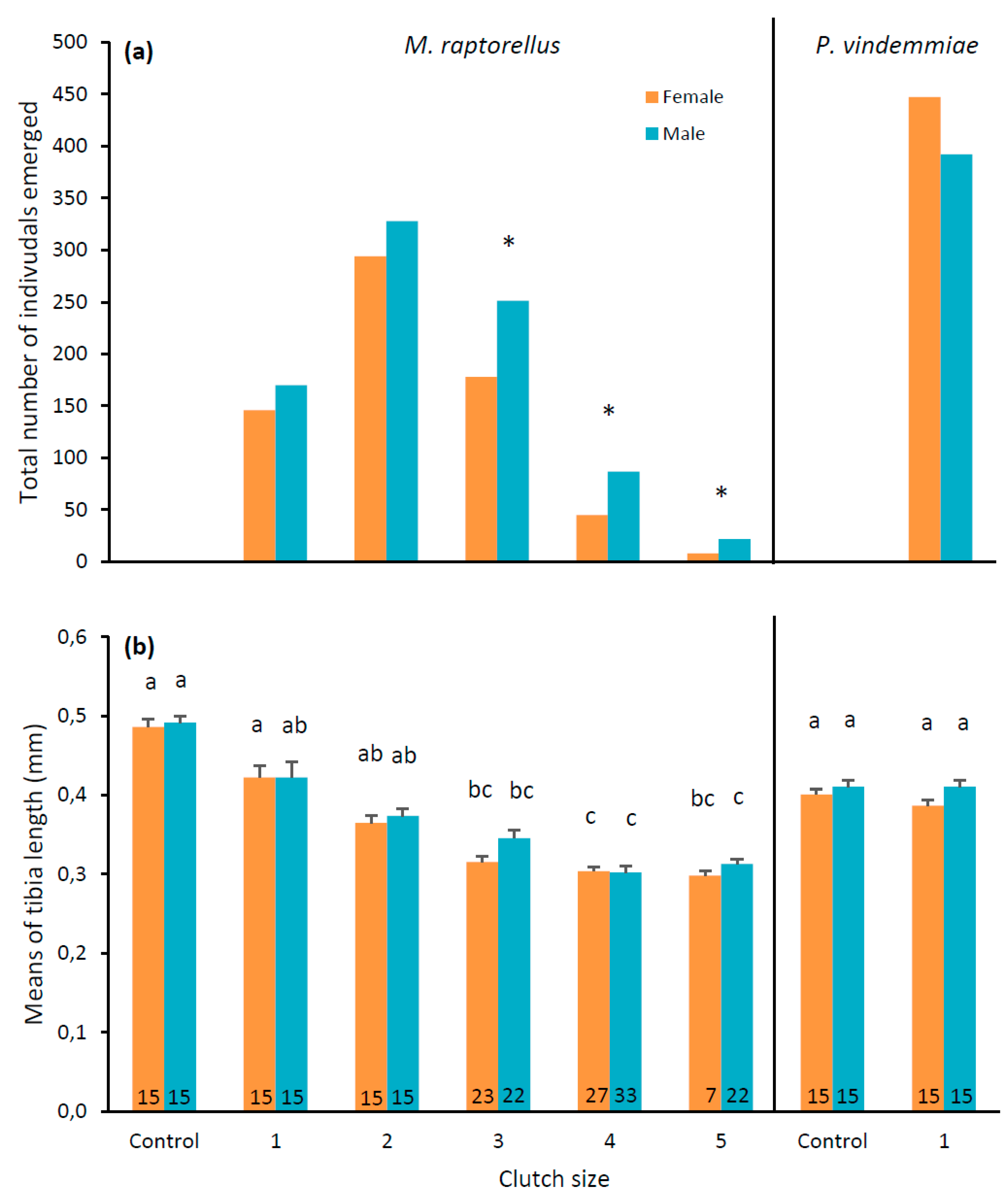
3.3. Pupal Parasitoid Observations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bolda, M.P.; Goodhue, R.E.; Zalom, F.G. Spotted wing drosophila: Potential economic impact of a newly established pest. Agric. Res. Econ. Update 2010, 13, 5–8. [Google Scholar]
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufield, P. In Focus: Spotted wing drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; van Steenwyk, R.A. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Farnsworth, D.; Hamby, K.A.; Bolda, M.; Goodhue, R.E.; Williams, J.C.; Zalom, F.G. Economic analysis of revenue losses and control costs associated with the spotted wing drosophila, Drosophila suzukii (Matsumura), in the California raspberry industry. Pest Manag. Sci. 2017, 73, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Beers, E.H.; van Steenwyk, R.A.; Shearer, P.W.; Coates, W.W.; Grant, J.A. Developing Drosophila suzukii management programs for sweet cherry in the western United States. Pest Manag. Sci. 2011, 67, 1386–1395. [Google Scholar] [CrossRef] [PubMed]
- Emiljanowicz, L.M.; Ryan, G.D.; Langille, A.; Newman, J. Development, reproductive output and population growth of the fruit fly pest Drosophila suzukii (Diptera: Drosophilidae) on artificial diet. J. Econ. Entomol. 2014, 107, 1392–1398. [Google Scholar] [CrossRef] [PubMed]
- SAgE Pesticides. Available online: https://www.sagepesticides.qc.ca/ (accessed on 15 November 2018).
- Cormier, D.; Veilleux, J.; Firlej, A. Exclusion net to control spotted wing drosophila in blueberry fields. IOBC WPRS Bull. 2015, 109, 181–184. [Google Scholar]
- Baroffio, C.; Kuonen, F.; Huber, B.; Kopp, M.C.; Marazzi, C.; Thoss, H.; Vulliemin, D.; Zurfluh, M. Drosophila suzukii: Stratégie 2018 pour les petits fruits. Plantes Agroscope Fiche Tech. 2018, 78, 1–2. [Google Scholar]
- Haye, T.; Girod, P.; Cuthbertson, A.G.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Baroffio, C.; Zhang, J.P.; Desneux, N. Current SWD IPM tactics and their practical implementation in fruit crops across different regions around the world. J. Pest Sci. 2016, 89, 643–651. [Google Scholar] [CrossRef]
- Klick, J.; Yang, W.Q.; Walton, V.M.; Dalton, D.T.; Hagler, J.R.; Dreves, A.J.; Lee, J.C.; Bruck, D.J. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. J. Appl. Entomol. 2015, 140, 37–46. [Google Scholar] [CrossRef]
- Knoll, V.; Ellenbroek, T.; Romeis, J.; Collatz, J. Seasonal and regional presence of hymenopteran parasitoids of Drosophila in Switzerland and their ability to parasitize the invasive Drosophila suzukii. Sci. Rep. 2017, 7, 40697. [Google Scholar] [CrossRef] [PubMed]
- Hajek, A.E.; Eilenberg, J. Natural Enemies: An Introduction to Biological Control, 2nd ed.; Cambridge University Press: Cambridge, UK, 2018; 452p. [Google Scholar]
- Cloutier, C.; Cloutier, C. Les solutions biologiques de lutte pour la répression des insectes et acariens ravageurs des cultures. In La Lutte Biologique; Vincent, C., Coderre, D., Editeur, G., Eds.; Morin: Boucherville, QC, Canada, 1992; Chapitre 2; 672p. [Google Scholar]
- Rossi Stacconi, M.V.; Grassi, A.; Ioriatti, C.; Anfora, G. Augmentative releases of Trichopria drosophilae for the suppression of early season Drosophila suzukii populations. BioControl 2018, 64, 9–19. [Google Scholar] [CrossRef]
- Daane, K.M.; Wang, X.G.; Biondi, A.; Miller, B.; Miller, J.C.; Riedl, H.; Shearer, P.W.; Guerrieri, E.; Giorgini, M.; Buffington, M.; et al. First exploration of parasitoids of Drosophila suzukii. J. Pest Sci. 2016, 89, 823–835. [Google Scholar] [CrossRef]
- Hajek, A.E.; Hurley, B.P.; Kenis, M.; Garnas, J.R.; Bush, S.J.; Wingfield, M.J.; van Lenteren, J.C.; Cock, M.J.W. Exotic biological control agents: A solution or contribution to arthropod invasions? Biol. Invasions 2016, 18, 953–969. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.G.; Kaçar, G.; Biondi, A.; Daane, K.M. Foraging efficiency and outcomes of interactions of two pupal parasitoids attacking the invasive spotted wing drosophila. Biol. Control 2016, 96, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Chabert, S.; Allemand, R.; Poyet, M.; Eslin, P.; Gibert, P. Ability of European parasitoids (Hymenoptera) to control a new invasive Asiatic pest, Drosophila suzukii. Biol. Control 2012, 63, 40–47. [Google Scholar] [CrossRef]
- Thistlewood, H.M.; Gibson, G.A.; Gillespie, D.R.; Fitzpatrick, S.M. Drosophila suzukii (Matsumura), Spotted Wing Drosophila (Diptera: Drosophilidae). In Biological Control Programmes in Canada 2001–2012; Mason, P.G., Gillespie, D.R., Eds.; CAB International: Wallingford, UK, 2013; Chapter 21; pp. 152–155. [Google Scholar]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Audsley, N. Efficacy of commercially available invertebrate predators against Drosophila suzukii. Insects 2014, 5, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Gabarra, R.; Riudavets, J.; Rodriguez, G.A.; Pujade-Villar, J.; Arnó, J. Prospects for the biological control of Drosophila suzukii. BioControl 2015, 60, 331–339. [Google Scholar] [CrossRef]
- Renkema, J.M.; Telfer, Z.; Gariepy, T.; Hallett, R.H. Dalotia coriaria as a predator of Drosophila suzukii: Functional responses, reduced fruit infestation and molecular diagnostics. Biol. Control 2015, 89, 1–10. [Google Scholar] [CrossRef]
- Woltz, J.M.; Donahue, K.M.; Bruck, D.J.; Lee, J.C. Efficacy of commercially available predators, nematodes and fungal entomopathogens for augmentative control of Drosophila suzukii. J. Appl. Entomol. 2015, 139, 759–770. [Google Scholar] [CrossRef]
- Firlej, A.; Vanoosthuyse, F. La Drosophile à Ailes Tachetées, un Ravageur des Petits Fruits au Québec; Synthèse Bibliographique; Institut de Recherche et de Développement en Agroenvironnement (IRDA) et Partenaires: Saint-Bruno-de-Montarville, QC, Canada, 2017; 93p. [Google Scholar]
- Schetelig, M.F.; Lee, K.Z.; Otto, S.; Talmann, L.; Stökl, J.; Degenkolb, T.; Vilcinskas, A.; Halitschke, R. Environmentally sustainable pest control options for Drosophila suzukii. J. Appl. Entomol. 2017, 142, 3–17. [Google Scholar] [CrossRef]
- Legner, E.F. Inheritance of gregarious and solitary oviposition in Muscidifurax raptorellus Kogan and Legner (Hymenoptera: Pteromalidae). Can. Entomol. 1987, 119, 791–808. [Google Scholar] [CrossRef]
- Petersen, J.J.; Currey, D.M. Timing of releases of gregarious Muscidifurax raptorellus (Hymenoptera: Pteromalidae) to control flies associated with confined beef cattle. J. Agric. Entomol. 1996, 13, 55–63. [Google Scholar]
- Minkenberg, O.P.; Van Lenteren, J.C. The Leafminers, Liriomyza bryoniae and L. trifolii (Diptera: Agromyzidae), Their Parasites and Host Plants: A Review; Agricultural University: Wageningen, The Netherlands, 1986; pp. 16–17. [Google Scholar]
- Featured Creatures. Available online: http://entnemdept.ufl.edu/creatures/beneficial/podisus_maculiventris.htm (accessed on 15 November 2018).
- McGregor, R.R.; Gillespie, D.R.; Quiring, D.M.; Foisy, M.R. Potential use of Dicyphus hesperus Knight (Heteroptera: Miridae) for biological control of pests of greenhouse tomatoes. Biol. Control 1999, 16, 104–110. [Google Scholar] [CrossRef]
- Arnó, J.; Riudavets, J.; Gabarra, R. Survey of host plants and natural enemies of Drosophila suzukii in an area of strawberry production in Catalonia (northeast Spain). IOBC Bull. 2012, 80, 29–34. [Google Scholar]
- Albouy, V. La Lutte Biologique au Jardin; Éditions Quae: Paris, France, 2012; 102p. [Google Scholar]
- New, T.R. The biology of Chrysopidae and Hemerobiidae (Neuroptera), with reference to their usage as biocontrol agents: A review. Trans. R. Entomol. Soc. Lond. 1975, 127, 115–140. [Google Scholar] [CrossRef]
- Renkema, J.M.; Cuthbertson, A.G.S. Impact of multiple natural enemies on immature Drosophila suzukii in strawberries and blueberries. BioControl 2018, 63, 719–728. [Google Scholar] [CrossRef]
- Jervis, M.A. Insects as Natural Enemies: A Practical Perspective; Springer Science & Business Media: Dordrecht, The Netherlands, 2007; 748p. [Google Scholar]
- Godfray, H.C.J. Parasitoids: Behavioral and Evolutionary Ecology; Princeton University Press Books: Princeton, NJ, USA, 1994; 473p. [Google Scholar]
- Rossi Stacconi, M.V.; Grassi, A.; Dalton, D.T.; Miller, B.; Ouantar, M.; Loni, A.; Ioriatti, C.; Walton, V.M.; Anfora, G. First field records of Pachycrepoideus vindemmiae as a parasitoid of Drosophila suzukii in European and Oregon small fruit production areas. Entomolgia 2013, 1, 11–16. [Google Scholar]
- Cheah, C.A.; Coaker, T.H. Host finding and discrimination in Diglyphus isaea, a parasitoid of the chrysanthemum leaf miner, Chromatomyia syngenesiae. Biocontrol Sci. Technol. 1992, 2, 109–118. [Google Scholar] [CrossRef]
- Van Driesche, R.; Hoddle, M.; Center, T. Control of Pests and Weeds by Natural Enemies: An Introduction to Biological Control; Blackwell Publishing: Singapore, 2008; 473p. [Google Scholar]
- Harvey, J.; Vet, L.; Jiang, N.; Gols, R. Nutritional ecology of the interaction between larvae of the gregarious ectoparasitoid, Muscidifurax raptorellus (Hymenoptera: Pteromalidae), and their pupal host, Musca domestica (Diptera: Muscidae). Physiol. Entomol. 1998, 23, 113–120. [Google Scholar] [CrossRef]
- Geden, C.J.; Moon, R.D. Host ranges of gregarious muscoid fly parasitoids: Muscidifurax raptorellus (Hymenoptera: Pteromalidae), Tachinaephagus zealandicus (Hymenoptera: Encyrtidae), and Trichopria nigra (Hymenoptera: Diapriidae). Environ. Entomol. 2009, 38, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Mackauer, M.; Sequeira, R. Patterns of Development in Insect Parasites. In Parasites and Pathogens of Insects; Beckage, N., Thompson, S., Federici, B., Eds.; Academic Press, Elsevier: Cambridge, MA, USA, 1993; 364p. [Google Scholar]
- Charnov, E.L. The Theory of Sex Allocation; Princeton University Press Books: Princeton, NJ, USA, 1982; 355p. [Google Scholar]
- Harvey, J.A.; Gols, G.J.Z. The influence of host quality on progeny and sex allocation in the pupal ectoparasitoid, Muscidifurax raptorellus (Hymenoptera: Pteromalidae). Bull. Entomol. Res. 1998, 88, 299–304. [Google Scholar] [CrossRef]
- Abram, P.; Agriculture and Agri-Food Canada, Agassiz, BC, Canada. Personal communication, 2019.
- Ballman, E.S.; Collins, J.A.; Drummond, F.A. Pupation Behavior and Predation on Drosophila suzukii (Diptera: Drosophilidae) Pupae in Maine Wild Blueberry Fields. J. Econ. Entomol. 2017, 110, 2308–2317. [Google Scholar] [CrossRef] [PubMed]
- Woltz, J.M.; Lee, J.C. Pupation behavior and larval and pupal biocontrol of Drosophila suzukii in the field. Biol. Control 2017, 110, 62–69. [Google Scholar] [CrossRef]
- Preisser, E.L.; Bolnick, D.I.; Benard, M.F. Scared to death? The effects of intimidation and consumption in predator-prey interactions. Ecology 2005, 86, 501–509. [Google Scholar] [CrossRef]
- Rossi Stacconi, M.V.; Buffington, M.; Daane, K.M.; Dalton, D.T.; Grassi, A.; Kaçar, G.; Miller, B.; Miller, J.C.; Baser, N.; Ioriatti, C.; et al. Host stage preference, efficacy and fecundity of parasitoids attacking Drosophila suzukii in newly invaded areas. Biol. Control 2015, 84, 28–35. [Google Scholar] [CrossRef]
- Miller, B.; Anfora, G.; Buffington, M.; Daane, K.M.; Dalton, D.T.; Hoelmer, K.M.; Rossi-Stacconi, M.V.; Grassi, A.; Ioriatti, C.; Loni, A.; et al. Seasonal occurrence of resident parasitoids associated with Drosophila suzukii in two small fruit production regions of Italy and the U.S.A. Bull. Insectol. 2015, 68, 255–263. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
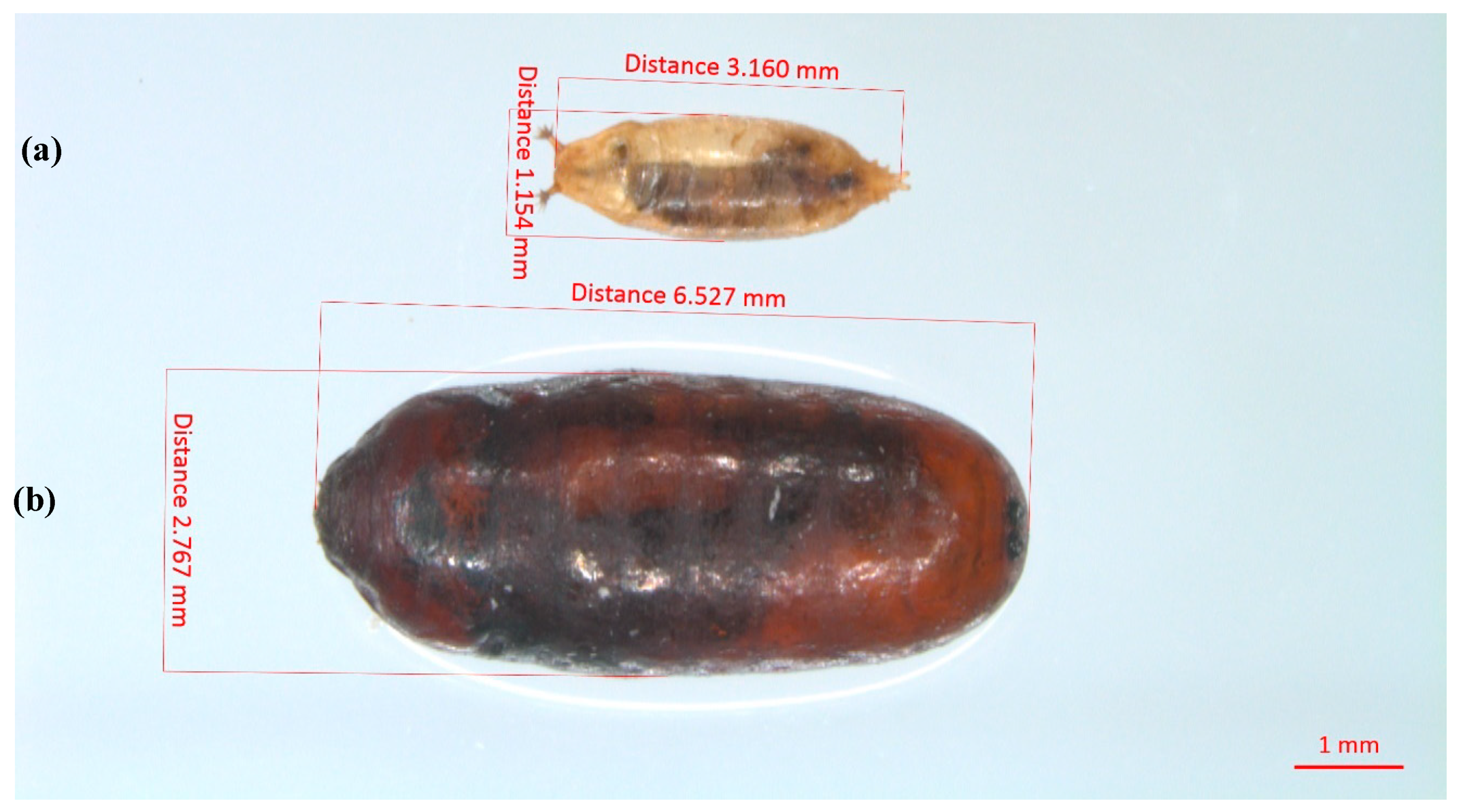{kind=link}
| Parasitoid Species | Mean Number of D. suzukii Pupae per Repetition (± SD) | Mean Number of Parasitized D. suzukii Pupae per Repetition with Parasitoid Emergence (± SD) | Successful Parasitism (%) |
|---|---|---|---|
| M. raptorellus | 232.3 ± 50.9 | 100.9 ± 26.1 a | 43.4 |
| P. vindemmiae | 232.5 ± 45.5 | 104.3 ± 21.2 a | 44.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonneau, P.; Renkema, J.; Fournier, V.; Firlej, A. Ability of Muscidifurax raptorellus and Other Parasitoids and Predators to Control Drosophila suzukii Populations in Raspberries in the Laboratory. Insects 2019, 10, 68. https://doi.org/10.3390/insects10030068
Bonneau P, Renkema J, Fournier V, Firlej A. Ability of Muscidifurax raptorellus and Other Parasitoids and Predators to Control Drosophila suzukii Populations in Raspberries in the Laboratory. Insects. 2019; 10(3):68. https://doi.org/10.3390/insects10030068
Chicago/Turabian StyleBonneau, Phanie, Justin Renkema, Valérie Fournier, and Annabelle Firlej. 2019. "Ability of Muscidifurax raptorellus and Other Parasitoids and Predators to Control Drosophila suzukii Populations in Raspberries in the Laboratory" Insects 10, no. 3: 68. https://doi.org/10.3390/insects10030068