Life History Effects Linked to an Advantage for wAu Wolbachia in Drosophila



Abstract
:1. Introduction
2. Methods
2.1. Lines and Infection Status
2.2. Comparison of Lines on Grapes with Fungal Growth
2.3. Comparisons of Lines on Laboratory Medium
2.4. Toxin Effects
3. Results
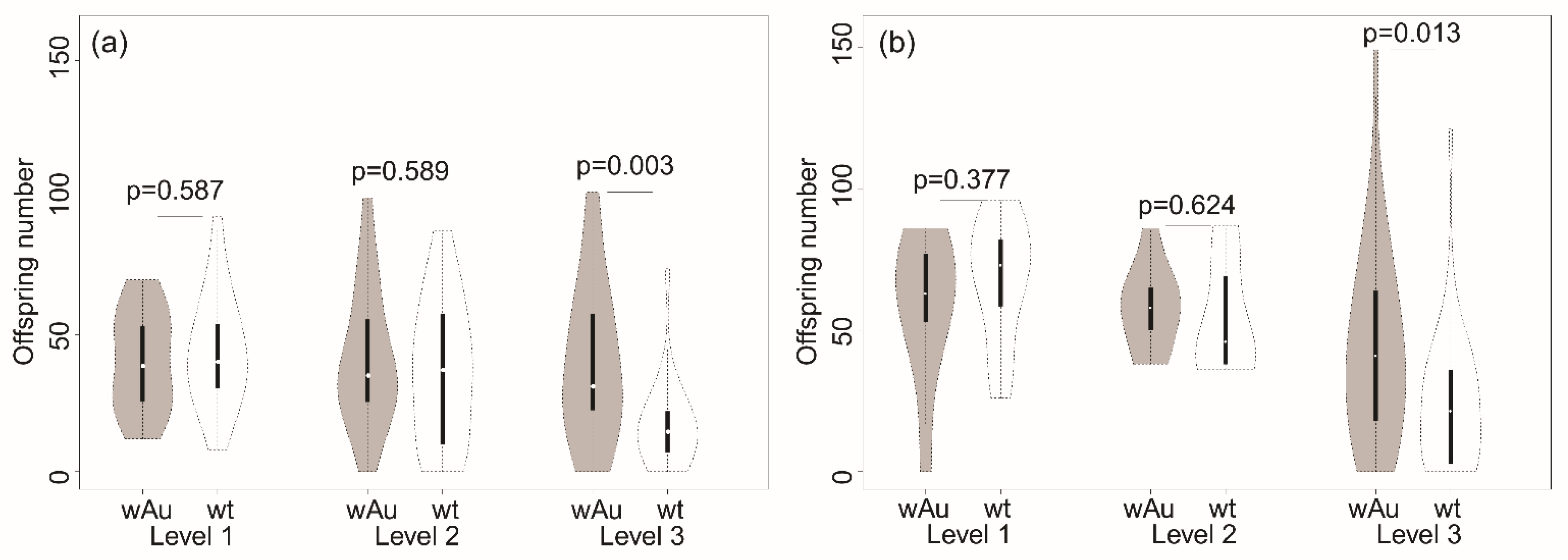
3.1. Comparison of Lines on Grapes with Fungal Growth
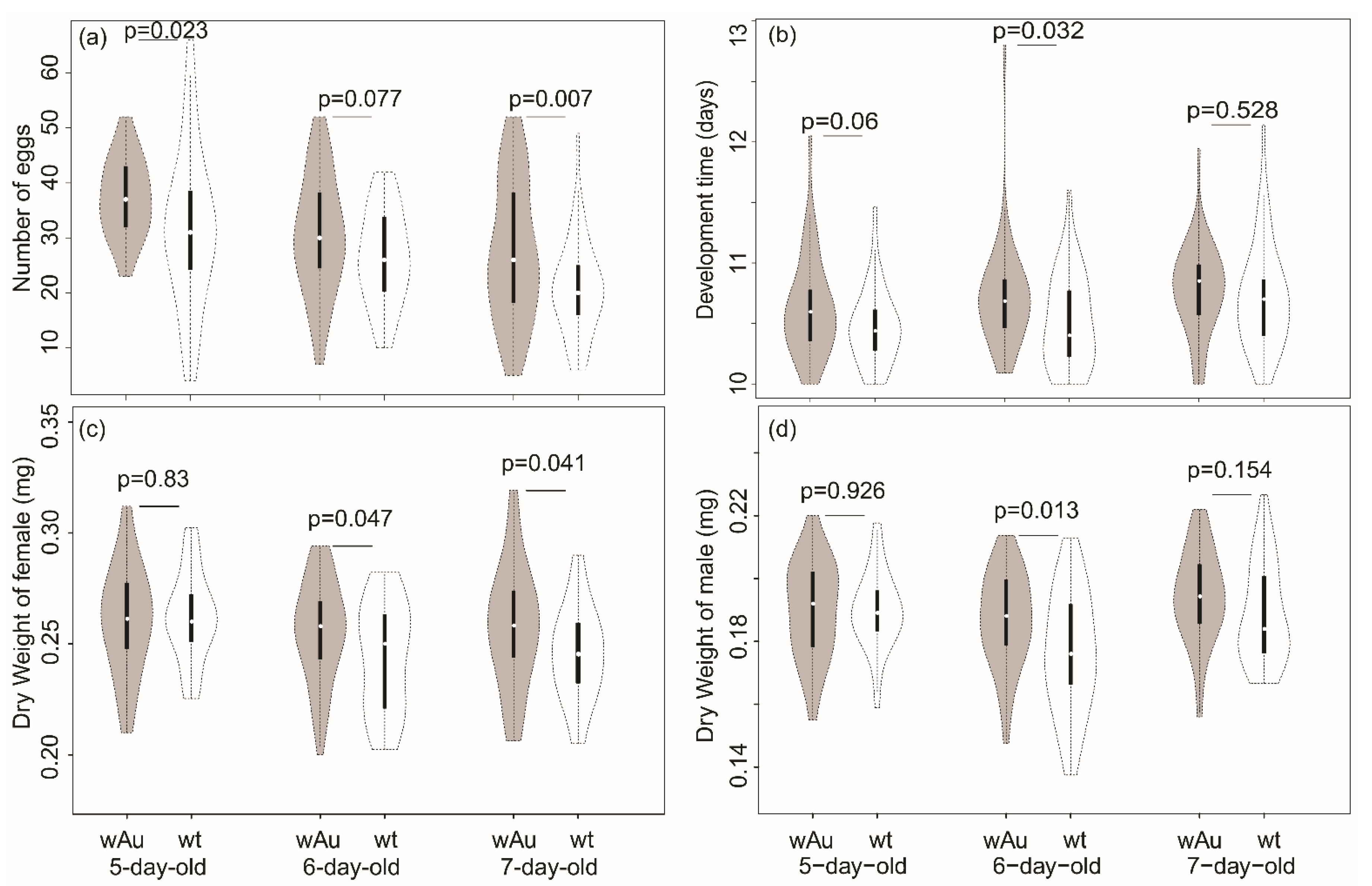
3.2. Comparison of Lines on Laboratory Medium
3.3. Toxin Effects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pais, R.; Lohs, C.; Wu, Y.N.; Wang, J.W.; Aksoy, S. The obligate mutualist Wigglesworthia glossinidia influences reproduction, digestion, and immunity processes of its host, the tsetse fly. Appl. Environ. Microbiol. 2008, 74, 5965–5974. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. Nutritional interactions in insect-microbial symbioses: Aphids and their symbiotic bacteria Buchnera. Annu. Rev. Entomol. 1998, 43, 17–37. [Google Scholar] [CrossRef]
- Duron, O.; Morel, O.; Noel, V.; Buysse, M.; Binetruy, F.; Lancelot, R.; Loire, E.; Menard, C.; Bouchez, O.; Vavre, F.; et al. Tick-bacteria mutualism depends on B vitamin synthesis pthways. Curr. Biol. 2018, 28, 1896–1902. [Google Scholar] [CrossRef] [PubMed]
- Koga, R.; Tsuchida, T.; Fukatsu, T. Changing partners in an obligate symbiosis: A facultative endosymbiont can compensate for loss of the essential endosymbiont Buchnera in an aphid. Proc. R. Soc. B Biol. Sci. 2003, 270, 2543–2550. [Google Scholar] [CrossRef] [PubMed]
- Sazama, E.J.; Bosch, M.J.; Shouldis, C.S.; Ouellette, S.P.; Wesner, J.S. Incidence of Wolbachia in aquatic insects. Ecol. Evol. 2017, 7, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia: Analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PloS ONE 2012, 7, e38544. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Hoffmann, A.A.; Werren, J.H. Influential Passengers: Inherited Microorganisms and Arthropod Reproduction; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Ant, T.H.; Herd, C.S.; Geoghegan, V.; Hoffmann, A.A.; Sinkins, S.P. The Wolbachia strain wAu provides highly efficient virus transmission blocking in Aedes aegypti. PLoS Pathog. 2018, 14, e1006815. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.J.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.W.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z.Y. The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti. Plos Pathogens 2010, 6, e1000833. [Google Scholar] [CrossRef] [PubMed]
- Caspari, E.; Watson, G.S. On the evolutionary importance of cytoplasmic sterility in mosquitoes. Evolution 1959, 13, 568–570. [Google Scholar] [CrossRef]
- Turelli, M.; Hoffmann, A.A. Rapid spread of an inherited incompatibility factor in California Drosophila. Nature 1991, 353, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, M.; Nikoh, N.; Hosokawa, T.; Fukatsu, T. Riboflavin provisioning underlies Wolbachia’s fitness contribution to its insect host. Mbio 2015, 6, e01732-15. [Google Scholar] [CrossRef]
- Taylor, M.J.; Bandi, C.; Hoerauf, A. Wolbachia bacterial endosymbionts of filarial nematodes. In Advances in Parasitology, Volume 60; Baker, J.R., Muller, R., Rollinson, D., Eds.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2005; Volume 60, pp. 245–284. [Google Scholar]
- Brownlie, J.C.; Cass, B.N.; Riegler, M.; Witsenburg, J.J.; Iturbe-Ormaetxe, I.; McGraw, E.A.; O’Neill, S.L. Evidence for metabolic provisioning by a common invertebrate endosymbiont, Wolbachia pipientis, during periods of nutritional stress. Plos Pathog. 2009, 5, e1000368. [Google Scholar] [CrossRef]
- Hamm, C.A.; Begun, D.J.; Vo, A.; Smith, C.C.R.; Saelao, P.; Shaver, A.O.; Jaenike, J.; Turelli, M. Wolbachia do not live by reproductive manipulation alone: Infection polymorphism in Drosophila suzukii and D. subpulchrella. Mol. Ecol. 2014, 23, 4871–4885. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Clancy, D.; Duncan, J. Naturally-occurring Wolbachia infection in Drosophila simulans that does not cause cytoplasmic incompatibility. Heredity 1996, 76, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kriesner, P.; Conner, W.R.; Weeks, A.R.; Turelli, M.; Hoffmann, A.A. Persistence of a Wolbachia infection frequency cline in Drosophila melanogaster and the possible role of reproductive dormancy. Evolution 2016, 70, 979–997. [Google Scholar] [CrossRef] [PubMed]
- Kriesner, P.; Hoffmann, A.A. Rapid spread of a Wolbachia infection that does not affect host reproduction in Drosophila simulans cage populations. Evolution 2018, 72, 1475–1487. [Google Scholar] [CrossRef] [PubMed]
- Dobson, S.L.; Marsland, E.J.; Rattanadechakul, W. Mutualistic Wolbachia infection in Aedes albopictus: Accelerating cytoplasmic drive. Genetics 2002, 160, 1087–1094. [Google Scholar]
- Fenton, A.; Johnson, K.N.; Brownlie, J.C.; Hurst, G.D.D. Solving the Wolbachia paradox: Modeling the tripartite interaction between host, Wolbachia, and a natural enemy. Am. Nat. 2011, 178, 333–342. [Google Scholar] [CrossRef]
- Lee, S.F.; White, V.L.; Weeks, A.R.; Hoffmann, A.A.; Endersby, N.M. High-throughput PCR assays to monitor Wolbachia infection in the dengue mosquito (Aedes aegypti) and Drosophila simulans. Appl. Environ. Microbiol. 2012, 78, 4740–4743. [Google Scholar] [CrossRef]
- Kriesner, P.; Hoffmann, A.A.; Lee, S.F.; Turelli, M.; Weeks, A.R. Rapid sequential spread of two Wolbachia variants in Drosophila simulans. PLoS Pathog. 2013, 9, e1003607. [Google Scholar] [CrossRef] [PubMed]
- Alkhayyat, F.; Yu, J.-H. Upstream regulation of mycotoxin biosynthesis. In Advances in Applied Microbiology; Sariaslani, S., Gadd, G.M., Eds.; Academic Press: New York, NY, USA, 2014; Volume 86, pp. 251–278. [Google Scholar]
- Mutlu, A.G. Increase in mitochondrial DNA copy number in response to Ochratoxin A and methanol-induced mitochondrial DNA damage in Drosophila. Bull. Environ. Contam. Toxicol. 2012, 89, 1129–1132. [Google Scholar] [CrossRef]
- Begg, M.; Robertson, F.W. The nutritional requirements of Drosophila melanogaster. J. Exp. Biol. 1950, 26, 380–387. [Google Scholar] [CrossRef]
- Ballard, J.W.O.; Melvin, R.G. Tetracycline treatment influences mitochondrial metabolism and mtDNA density two generations after treatment in Drosophila. Insect Mol. Biol. 2007, 16, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Parsons, P.A. The analysis of quantitative variation in natural populations with isofemale strains. Genet. Sel. Evol. 1988, 20, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, A.A.; Turelli, M. Cytoplasmic incompatibility in insects. In Influential Passengers: Microorganisms and Invertebrate Reproduction; O’Neill, S., Hoffmann, A., Werren, J., Eds.; Oxford University Press: Oxford, UK, 1997; pp. 42–80. [Google Scholar]
- Weeks, A.R.; Turelli, M.; Harcombe, W.R.; Reynolds, K.T.; Hoffmann, A.A. From parasite to mutualist: Rapid evolution of Wolbachia in natural populations of Drosophila. PloS Biol. 2007, 5, e114. [Google Scholar] [CrossRef] [PubMed]
- Fast, E.M.; Toomey, M.E.; Panaram, K.; Desjardins, D.; Kolaczyk, E.D.; Frydman, H.M. Wolbachia enhance Drosophila stem cell proliferation and target the germline stem cell niche. Science 2011, 334, 990–992. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.R.; Sun, J.T.; Xue, X.F.; Hong, X.Y. Cytoplasmic incompatibility and fitness benefits in the two-spotted spider mite Tetranychus urticae (red form) doubly infected with Wolbachia and Cardinium. Syst. Appl. Acarol. 2016, 21, 1161–1173. [Google Scholar] [CrossRef]
- Vala, F.; Egas, M.; Breeuwer, J.A.J.; Sabelis, M.W. Wolbachia affects oviposition and mating behaviour of its spider mite host. J. Evol. Biol. 2004, 17, 692–700. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Ross, P.A.; Rasic, G. Wolbachia strains for disease control: Ecological and evolutionary considerations. Evol. Appl. 2015, 8, 751–768. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Longdon, B.; Bauer, S.; Chan, Y.S.; Miller, W.J.; Bourtzis, K.; Teixeira, L.; Jiggins, F.M. Symbionts commonly provide broad spectrum resistance to viruses in insects: A comparative analysis of Wolbachia strains. Plos Pathog. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Osborne, S.E.; San Leong, Y.; O’Neill, S.L.; Johnson, K.N. Variation in antiviral protection mediated by different Wolbachia strains in Drosophila simulans. PLoS Pathog. 2009, 5, e1000656. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; White, V.L.; Schlub, T.; Eden, J.-S.; Hoffmann, A.A.; Holmes, E.C. No detectable effect of Wolbachia w Mel on the prevalence and abundance of the RNA virome of Drosophila melanogaster. Proc. R. Soc. B Biol. Sci. 2018, 285, 20181165. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A. Partial cytoplasmic incompatibility between two Australian populations of Drosophila melanogaster. Entomol. Exp. Appl. 1988, 48, 61–67. [Google Scholar] [CrossRef]
- Koch, R.E.; Phillips, J.M.; Camus, M.F.; Dowling, D.K. Maternal age effects on fecundity and offspring egg-to-adult viability are not affected by mitochondrial haplotype. Ecol. Evol. 2018, 8, 10722–10732. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Hercus, M.; Dagher, H. Population dynamics of the Wolbachia infection causing cytoplasmic incompatibility in Drosophila melanogaster. Genetics 1998, 148, 221–231. [Google Scholar] [PubMed]
- Sutton, E.R.; Harris, S.R.; Parkhill, J.; Sinkins, S.P. Comparative genome analysis of Wolbachia strain wAu. BMC Genom. 2014, 15, 928–942. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Experiment | Number of Isofemale Lines Used | Generation Since Isofemale Lines Established | Traits Scored |
|---|---|---|---|
| Comparison of lines on grapes with fungal growth | 23 wAu-infected and 23 uninfected lines | 2 | Number of offspring produced, dry weight, sex ratio |
| Comparisons of lines on laboratory medium | As above | 3 | Fecundity, egg-to-adult emergence, sex ratio, development time, dry weight |
| Toxin effects | As above | 6 | Egg-to-adult emergence, development time, dry weight |
| Variable | Infection (df = 1) | Fungal Level (df = 2) | Infection by Fungal Level (df = 2) | Error MS (df) | |||
|---|---|---|---|---|---|---|---|
| MS | F (P) | MS | F (P) | MS | F (P) | ||
| Offspring number (parents 9–11 days old) | 2431 | 4.708 (0.032) | 2355 | 4.561 (0.012) | 1875 | 3.631 (0.030) | 516 (118) |
| Offspring number (parents 12–14 days old) | 2519 | 3.579 (0.061) | 10921 | 15.52 (<0.001) | 1884 | 2.676 (0.074) | 704 (104) |
| Weight (mg, female) | 0.0000 | 0.001 (0.974) | 0.1577 | 37.99 (<0.001) | 0.0012 | 0.292 (0.747) | 0.0042 (191) |
| Weight (mg, male) | 0.0008 | 0.687 (0.408) | 0.0259 | 21.12 (<0.001) | 0.0005 | 0.386 (0.680) | 0.0012 (193) |
| Variable | Infection (df = 1) | Age (df = 2) | Infection by Age (df = 2) | Error MS (df) | |||
|---|---|---|---|---|---|---|---|
| MS/MRD | F (P) | MS/MRD | F (P) | MS/MRD | F (P) | ||
| Fecundity | 1778.9 | 15.96 (<0.001) | 2206.1 | 19.80 (<0.001) | 39.0 | 0.350 (0.705) | 111.4 (240) |
| Emergence rate (ranked data) | 0.00015 | 0.0012(0.972) | 0.0529 | 0.441 (0.644) | 0.0495 | 0.413 (0.662) | - (234) |
| Development time in days 1 | 1.1435 | 7.393 (0.007) | 0.8082 | 5.225 (0.007) | 0.1354 | 0.875 (0.418) | 0.1585 (229) |
| Weight (mg, female) | 0.0026 | 4.532 (0.034) | 0.0031 | 5.524 (0.005) | 0.0011 | 1.929 (0.148) | 0.5530 (208) |
| Weight (mg, male) | 0.0011 | 4.103 (0.044) | 0.0015 | 5.693 (0.004) | 0.0003 | 1.094 (0.337) | 0.0003 (196) |
| Variable | Infection (df = 1) | OTA (df = 2) | Infection by OTA (df = 2) | Error MS (df) | |||
|---|---|---|---|---|---|---|---|
| MS/MRD | F (P) | MS/MRD | F (P) | MS/MRD | F (P) | ||
| Emergence rate (ranked data) | 0.1289 | 4.733 (0.032) | 0.6885 | 25.277 (<0.001) | 0.0076 | 0.280 (0.756) | - (120) |
| Development time in days | 0.2801 | 25.419 (<0.001) | 0.1027 | 9.318 (<0.001) | 0.0289 | 2.619 (0.077) | 0.0110 (120) |
| Weight (mg, female) | 0.021 | 19.4 (<0.001) | 0.009 | 7.999 (<0.001) | 0.004 | 4.042 (0.020) | 0.0011 (120) |
| Weight (mg, male) | 0.000 | 0.359 (0.550) | 0.001 | 2.846 (0.062) | 0.000 | 1.075 (0.344) | 0.0003 (120) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, L.-J.; Jiang, W.; Hoffmann, A.A. Life History Effects Linked to an Advantage for wAu Wolbachia in Drosophila. Insects 2019, 10, 126. https://doi.org/10.3390/insects10050126
Cao L-J, Jiang W, Hoffmann AA. Life History Effects Linked to an Advantage for wAu Wolbachia in Drosophila. Insects. 2019; 10(5):126. https://doi.org/10.3390/insects10050126
Chicago/Turabian StyleCao, Li-Jun, Weibin Jiang, and Ary A. Hoffmann. 2019. "Life History Effects Linked to an Advantage for wAu Wolbachia in Drosophila" Insects 10, no. 5: 126. https://doi.org/10.3390/insects10050126
APA StyleCao, L.-J., Jiang, W., & Hoffmann, A. A. (2019). Life History Effects Linked to an Advantage for wAu Wolbachia in Drosophila. Insects, 10(5), 126. https://doi.org/10.3390/insects10050126