Seasonal Dynamics of the Leaffooted Bug Leptoglossus zonatus and Its Implications for Control in Almonds and Pistachios


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Annual Generations and Seasonal Development
Egg Strand Production and Egg Development
2.2. Overwintering Survival of Nymphs and Adults
2.3. Data Analysis
3. Results
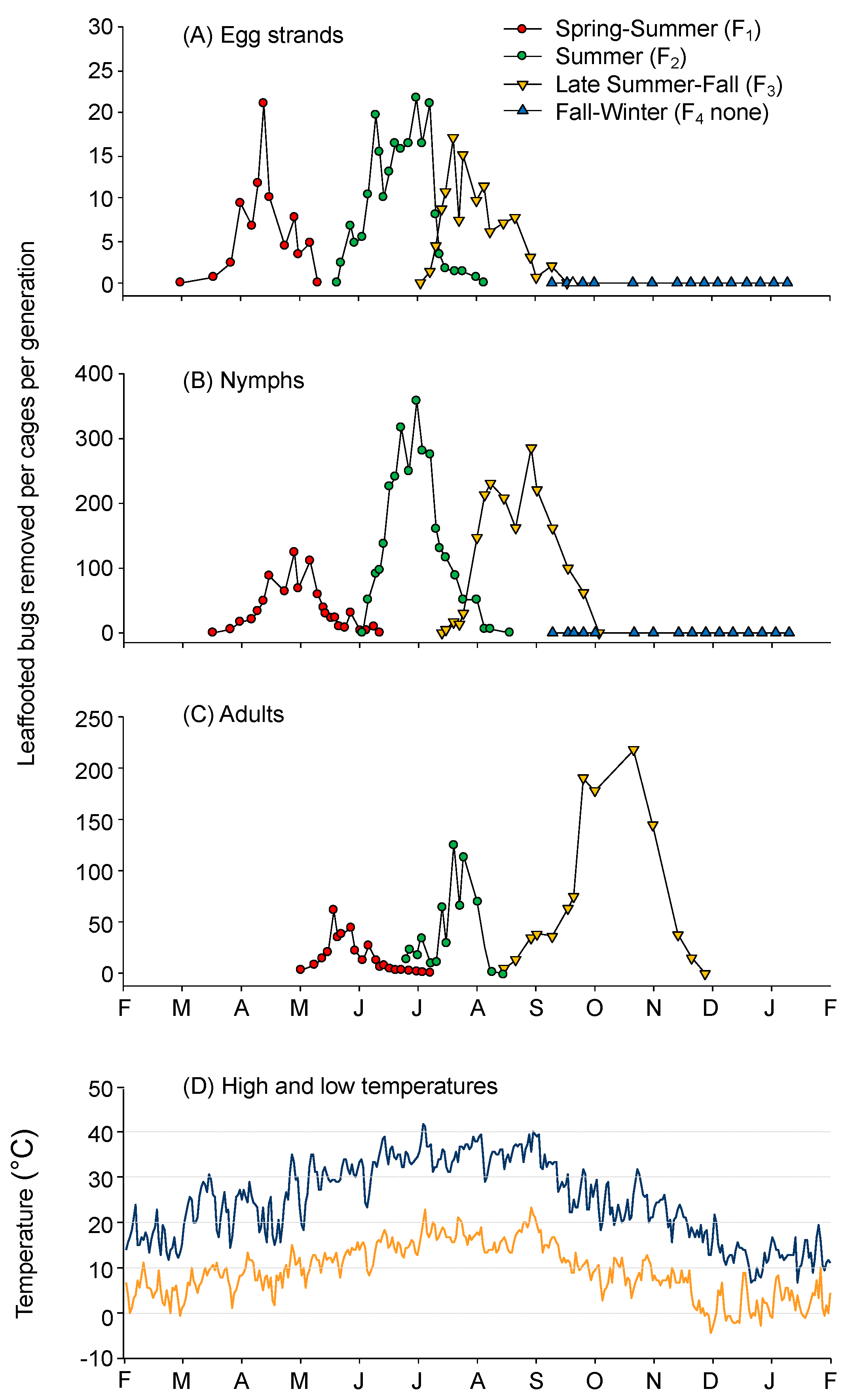
3.1. Annual Generations and Seasonal Development
Egg Strand Production and Egg Development
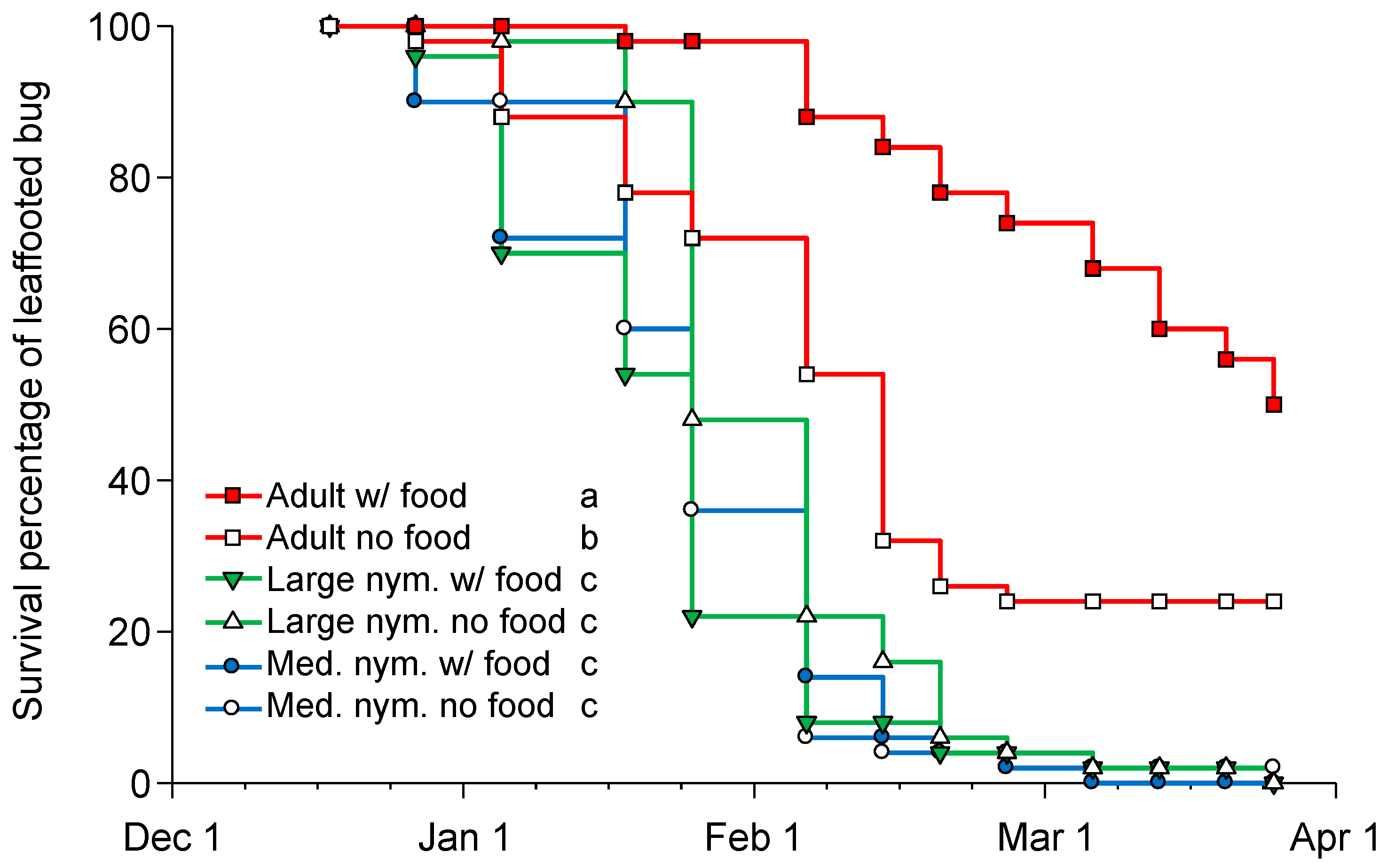
3.2. Overwintering Survival of Nymphs and Adults
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ferguson, L.; Haviland, D.R. Pistachio Production Manual, Publication 3545; University of California, Division of Agriculture and Natural Resources: Oakland, CA, USA, 2016. [Google Scholar]
- Zalom, F.G.; Haviland, D.R.; Symmes, E.J.; Tollerup, K. Almonds: Insects and Mites. In University of California IPM Pest. Management Guidelines, Publication 3431; University of California, Agriculture and Natural Resources: Oakland, CA, USA, 2018. [Google Scholar]
- Haviland, D.R.; Bentley, W.J.; Siegel, J.P.; Holtz, B.A.; Daane, K.M.; Higbee, B.S. Navel orangeworm and obliquebanded leafroller. In Pistachio Production Manual, Publication 3545; Ferguson, L., Haviland, D.R., Eds.; University of California, Agriculture and Natural Resources: Oakland, CA, USA, 2016; pp. 197–210. [Google Scholar]
- Hamby, K.A.; Zalom, F.G. Relationship of almond kernel damage occurrence to navel orangeworm (Lepidoptera: Pyralidae) success. J. Econ. Entomol. 2013, 106, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Daane, K.M.; Yokota, G.Y.; Bentley, W.J.; Weinberger, G.; Millar, J.G.; Beede, R.H. Stink bugs and leaffooted bugs. In Pistachio Production Manual; Ferguson, L., Haviland, D.R., Eds.; University of California, Agriculture and Natural Resources Publication 3545: Oakland, CA, USA, 2016; pp. 225–238. [Google Scholar]
- Bentley, W.J.; Goodell, P.B.; Beede, R.H. Small bugs: Neurocolpus, Phytocoris, Calocoris, and Lygus. In Pistachio Production Manual; Ferguson, L., Haviland, D.R., Eds.; University of California, Agriculture and Natural Resources Publication 3545: Oakland, CA, USA, 2016; pp. 215–224. [Google Scholar]
- Michailides, T.J.; Rice, R.E.; Ogawa, J.M. Succession and significance of several hemipterans attaching a pistachio orchard. J. Econ. Entomol. 1987, 80, 398–406. [Google Scholar] [CrossRef]
- Uyemoto, J.K.; Ogawa, J.M.; Rice, R.E.; Teranishi, H.R.; Bostock, R.M.; Pemberton, W.M. Role of several true bugs (Hemiptera) on incidence and seasonal development of pistachio fruit epicarp lesion disorder. J. Econ. Entomol. 1986, 79, 395–399. [Google Scholar] [CrossRef]
- Purcell, M.; Welter, S.C. Effect of Calocoris norvegicus (Hemiptera: Miridae) on pistachio yields. J. Econ. Entomol. 1991, 84, 114–119. [Google Scholar] [CrossRef]
- Daane, K.M.; Yokota, G.Y.; Krugner, R.; Steffan, S.A.; Silva, P.G.; Beede, R.H.; Bentley, W.J.; Weinberger, G.B. Large bugs damage pistachio nuts most severely during midseason. Calif. Agric. 2005, 59, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Michailides, T.J. The Achilles heel of pistachio fruit. Calif. Agric. 1989, 43, 10–11. [Google Scholar]
- Michailides, T.J.; Morgan, D.P. Association of Botryosphaeria panicle and shoot blight of pistachio with injuries of fruit caused by Hemiptera insects and birds. Plant Dis. 2016, 100, 1405–1413. [Google Scholar] [CrossRef]
- Haviland, D.R.; Bentley, W.J.; Beede, R.H.; Daane, K.M. Pistachios: Insects and Mites. In University of California IPM Pest. Management Guidelines, Publication 3461; University of California, Agriculture and Natural Resources: Oakland, CA, USA, 2018. [Google Scholar]
- Demkovich, M.; Siegel, J.P.; Higbee, B.S.; Berenbaum, M.R. Mechanism of resistance acquisition and potential associated fitness costs in Amyelois transitella (Lepidoptera: Pyralidae) exposed to pyrethroid insecticides. Environ. Entomol. 2015, 44, 855–863. [Google Scholar] [CrossRef]
- Purcell, M.; Welter, S.C. Seasonal phenology and biology of Calocoris norvegicus (Hemipteran: Miridae) in pistachio and associated host plants. J. Econ. Entomol. 1990, 83, 1841–1846. [Google Scholar] [CrossRef]
- Daane, K.M.; Haviland, D.R.; Viveros, M.; Holtz, B.A. Leaffooted bug hammers almonds, pistachios. Pac. Nut Prod. 2006, 12, 4–7. [Google Scholar]
- Michailides, T.J.; Ogawa, J.M.; Rice, R.E. Sites of epicarp lesion and kernel necrosis in relationship to symptoms and phenology of pistachio fruit. J. Econ. Entomol. 1988, 81, 1152–1154. [Google Scholar] [CrossRef]
- Bostock, R.M.; Thomas, C.S.; Ogawa, J.M.; Rice, R.E.; Uyemoto, J.K. Relationship of wound-induced peroxidase activity to epicarp lesion development in maturing pistachio fruit. Phytopathology 1987, 77, 275–282. [Google Scholar] [CrossRef]
- Beck, J.J.; Gee, W.S.; Cheng, L.W.; Higbee, B.S.; Wilson, H.; Daane, K.M. Investigating host plant-based semiochemicals for attracting the leaffooted bug (Hemiptera: Coreidae), an insect pest of California agriculture. In Roles of Natural Products for Biorational Pesticides in Agriculture; Beck, J.J., Rering, C.C., Duke, S.O., Eds.; American Chemical Society: Washington, WA, USA, 2018; Volume 1294, pp. 143–165. [Google Scholar]
- Inoue, K.M.; Vidal, D.M.; Saad, E.B.; Martins, C.B.C.; Zarbin, P.H.G. Identification of the alarm and sex Pheromones of the leaf-footed bug, Leptoglossus zonatus (Heteroptera: Coreidae). J. Braz. Chem. Soc. 2019, 30, 939–947. [Google Scholar] [CrossRef]
- Franco-Archundia, S.L.; Gonzaga-Segura, A.J.; Jimenez-Perez, A.; Castrejon-Gomez, V.R. Behavioral response of Leptoglossus zonatus (Heteroptera: Coreidae) to stimuli based on colors and its aggregation pheromone. Insects 2018, 9, 91. [Google Scholar] [CrossRef]
- Wang, Q.; Millar, J.G. Mating behavior and evidence for male-produced sex pheromones in Leptoglossus clypealis (Heteroptera: Coreidae). Ann. Entomol. Soc. Am. 2000, 93, 972–976. [Google Scholar] [CrossRef]
- Huffaker, C.B.; Berryman, C.A.; Turchin, P. Dynamics and regulation of insect populations. In Ecological Entomology; Huffaker, C.B., Gutierrez, A.P., Eds.; John Wiley and Sons: New York, NY, USA, 1999; pp. 269–280. [Google Scholar]
- Barta, M. Biology and temperature requirements of the invasive seed bug Leptoglossus occidentalis (Heteroptera: Coreidae) in Europe. J. Pest. Sci. 2016, 89, 31–44. [Google Scholar] [CrossRef]
- Xiao, Y.F.; Fadamiro, H.Y. Host preference and development of Leptoglossus zonatus (Hemiptera: Coreidae) on Satsuma Mandarin. J. Econ. Entomol. 2009, 102, 1908–1914. [Google Scholar] [CrossRef]
- Panizzi, A.R. Nymph and adult performance of Leptoglossus zonatus (Dallas 1852) (Hemiptera: Coreidae) on different foods. An. Soc. Entomol. Bras. 1989, 18, 375–390. [Google Scholar]
- Packauskas, R.J.; Schaefer, C.W. Clarification of some taxonomic problems in Anisoscelini and Leptoscelini (Hemiptera: Coreidae: Coreinae). Proc. Entomol. Soc. Wash. 2001, 103, 249–256. [Google Scholar]
- Allen, R.C. A revision of the genus Leptoglossus Guerin (Hemiptera: Coreidae). Entomol. Am. 1969, 45, 35–140. [Google Scholar]
- Foresti, J.; Bastos, C.S.; Fernandes, F.L.; da Silva, P.R. Economic injury levels and economic thresholds for Leptoglossus zonatus (Dallas) (Hemiptera: Coreidae) infesting seed maize. Pest. Manag. Sci. 2018, 74, 149–158. [Google Scholar] [CrossRef]
- Xiao, Y.F.; Fadamiro, H.Y. Evaluation of damage to satsuma mandarin (Citrus unshiu) by the leaffooted bug, Leptoglossus zonatus (Hemiptera: Coreidae). J. Appl. Entomol. 2010, 134, 694–703. [Google Scholar] [CrossRef]
- Tarango Rivero, S.H.; Gonzalez Hernandez, A. Species, seasonal occurrence, and natural enemies of stink bugs and leaffooted bugs (Hemiptera: Pentatomidae, Coreidae, Largidae) in pecans. Southwest. Entomol. 2009, 34, 305–318. [Google Scholar] [CrossRef]
- Grimm, C. Evaluation of damage to physic nut (Jatropha curcas) by true bugs. Entomol. Exp. Appl. 1999, 92, 127–136. [Google Scholar] [CrossRef]
- Joyce, A.L.; Higbee, B.S.; Haviland, D.R.; Brailovsky, H. Genetic variability of two leaffooted bugs, Leptoglossus clypealis and Leptoglossus zonatus (Hemiptera: Coreidae) in the Central Valley of California. J. Econ. Entomol. 2017, 110, 2576–2589. [Google Scholar] [CrossRef]
- Da Silva, P.G.; Daane, K.M. Life history parameters of Chinavia hilaris (Hemiptera: Pentatomidae), a stink bug injurious to pistachios in California. J. Econ. Entomol. 2014, 107, 166–173. [Google Scholar] [CrossRef]
- Yasuda, K.; Tsurumachi, M. Immigration of leaf-footed plant bug, Leptoglossus australis (Fabricius) (Heteroptera, Coreidae), into cucurbit fields on Ishigaki island, Japan, and role of male pheromone. J. Appl. Entomol. Zool. 1994, 38, 161–167. [Google Scholar] [CrossRef]
- Grimm, C.; Somarriba, A. Suitability of physic nut (Jatropha curcas L.) as single host plant for the leaf-footed bug Leptoglossus zonatus Dallas (Het, Coreidae). J. Appl. Entomol. 1999, 123, 347–350. [Google Scholar] [CrossRef]
- Jackson, C.G.; Tveten, M.S.; Figuli, P.J. Development. longevity and fecundity of Leptoglossus zonatus (Heteroptera, Coreidae) on a meridic diet. Southwest. Entomol. 1995, 20, 43–48. [Google Scholar]
- Duarte, I.S.; Nuñez, L.B.; Peña, J.E.; Pinto, L. Biology, parasitoids, and damage of Leptoglossus zonatus and Leptoglossus gonagra (Heteroptera: Coreidae) on citrus. Proc. Fla. State Hortic. Soc. 2008, 121, 5–9. [Google Scholar]
- Bolkan, H.A.; Ogawa, J.M.; Rice, R.E.; Bostock, R.M.; Crane, J.C. Leaffooted bug, Leptoglossus clypealis (Hemiptera: Coreidae) and epicarp lesions of pistachio, Pistacia vera, fruits. J. Econ. Entomol. 1984, 77, 1163–1165. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daane, K.M.; Yokota, G.Y.; Wilson, H. Seasonal Dynamics of the Leaffooted Bug Leptoglossus zonatus and Its Implications for Control in Almonds and Pistachios. Insects 2019, 10, 255. https://doi.org/10.3390/insects10080255
Daane KM, Yokota GY, Wilson H. Seasonal Dynamics of the Leaffooted Bug Leptoglossus zonatus and Its Implications for Control in Almonds and Pistachios. Insects. 2019; 10(8):255. https://doi.org/10.3390/insects10080255
Chicago/Turabian StyleDaane, Kent M., Glenn Y. Yokota, and Houston Wilson. 2019. "Seasonal Dynamics of the Leaffooted Bug Leptoglossus zonatus and Its Implications for Control in Almonds and Pistachios" Insects 10, no. 8: 255. https://doi.org/10.3390/insects10080255
APA StyleDaane, K. M., Yokota, G. Y., & Wilson, H. (2019). Seasonal Dynamics of the Leaffooted Bug Leptoglossus zonatus and Its Implications for Control in Almonds and Pistachios. Insects, 10(8), 255. https://doi.org/10.3390/insects10080255