Ameliorative Effects of Phytochemical Ingestion on Viral Infection in Honey Bees


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Particle Production
2.2. Virus Particle Extraction and Concentration
2.3. Virus Particle Purification and Quantification
2.4. Cage Assays: General Protocol
2.5. Phytochemical Screening
2.5.1. Carvacrol Moderate Dose Treatment (Experiment 1)
2.5.2. Carvacrol Acute Rescue (Experiment 2)
2.5.3. Thymol Acute Rescue, Prophylactic, and Regular Treatment (Experiment 3)
2.6. Caffeine Investigation
2.6.1. Differentiating Caffeine in Multiple Phytochemical Trial (Experiment 4)
2.6.2. Caffeine Trial (Experiment 5)
2.6.3. Caffeine Dose–Responses (Experiments 6A and 6B)
2.7. Sampled Bee Homogenization for Viral Quantification
2.8. Statistical Analyses
3. Results
3.1. Phytochemical Screening
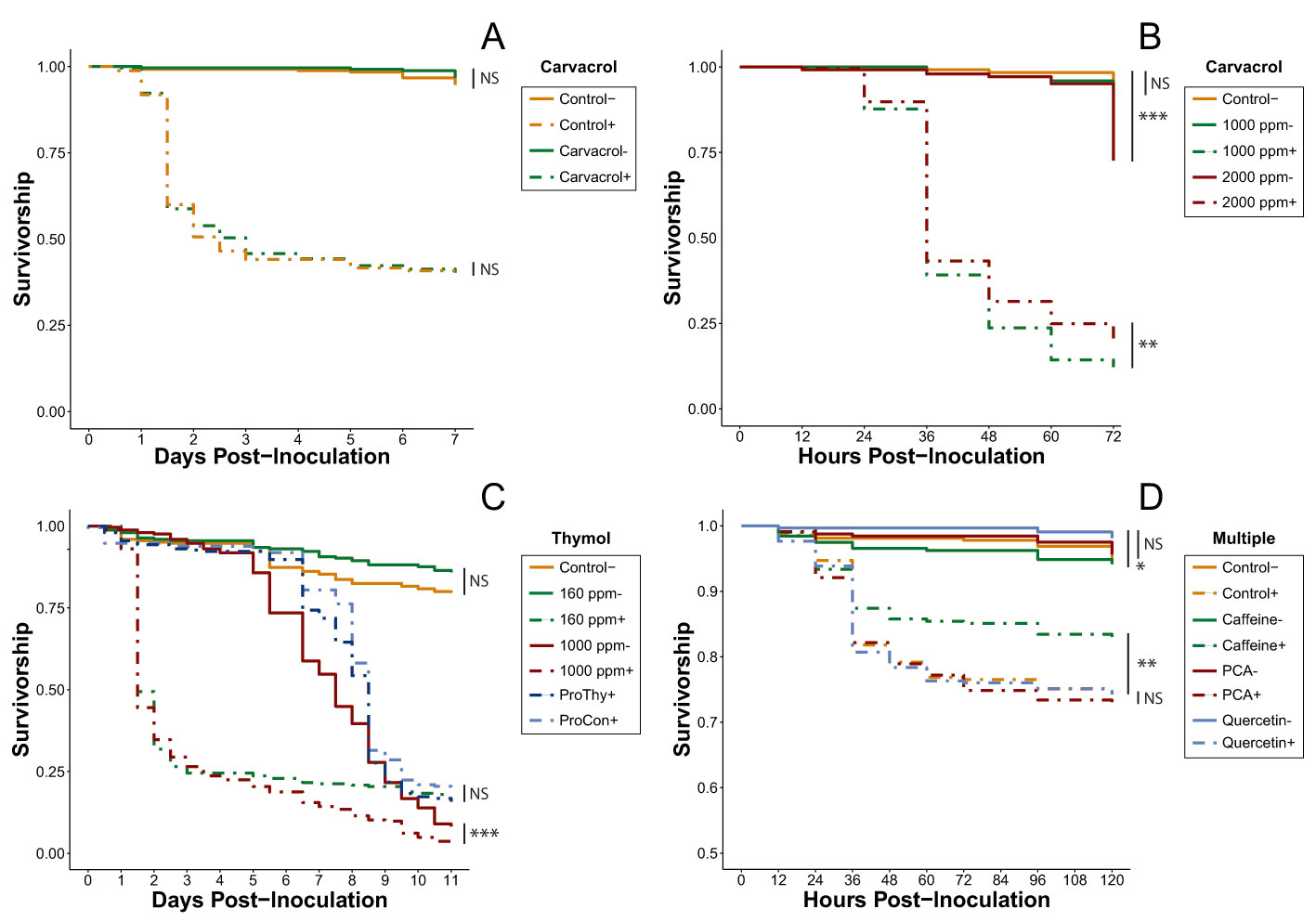
3.1.1. Monoterpene Screening (Experiments 1–3)
3.1.2. Multi-Phytochemical Screening (Experiment 4)
3.2. Caffeine Investigation
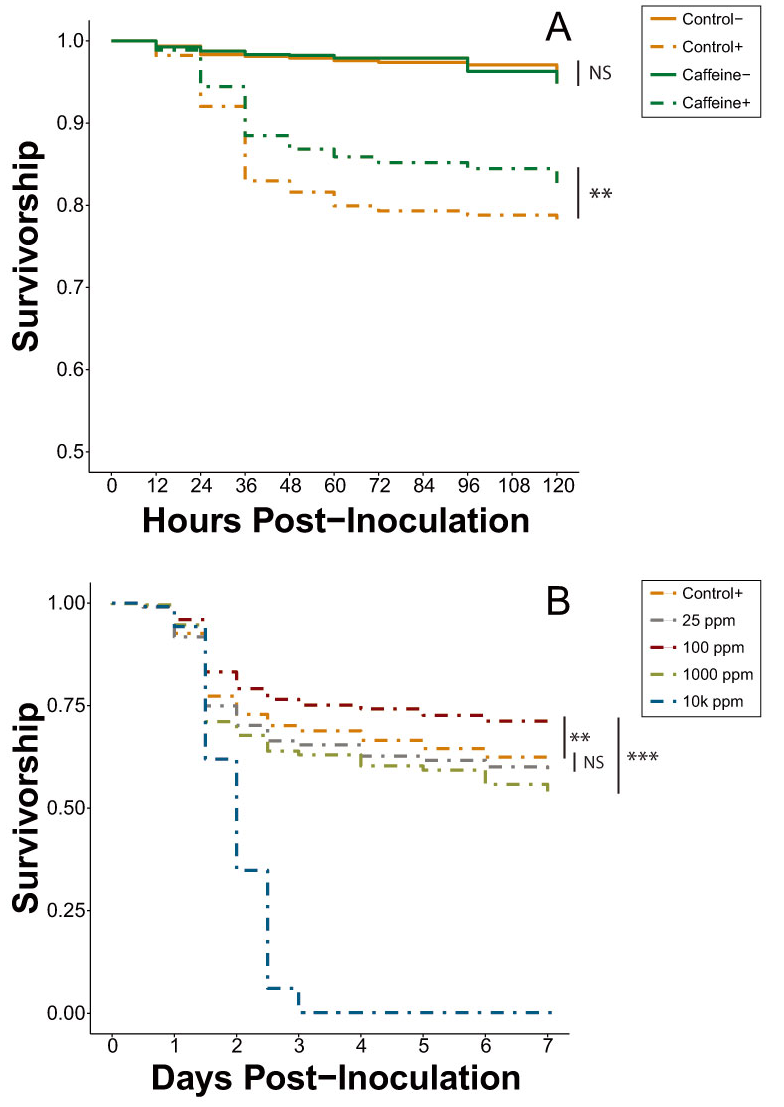
3.2.1. Caffeine Trials (Experiment 5)
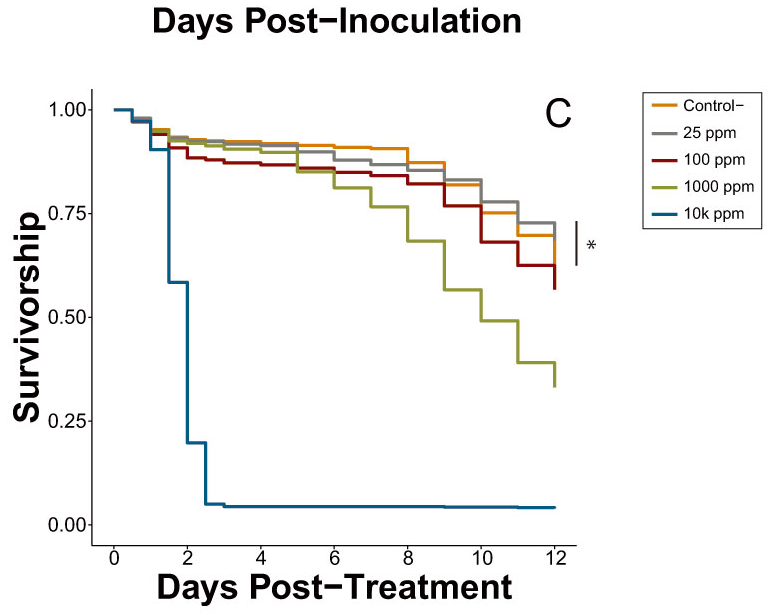
3.2.2. Caffeine Dose-Response (Experiment 6)
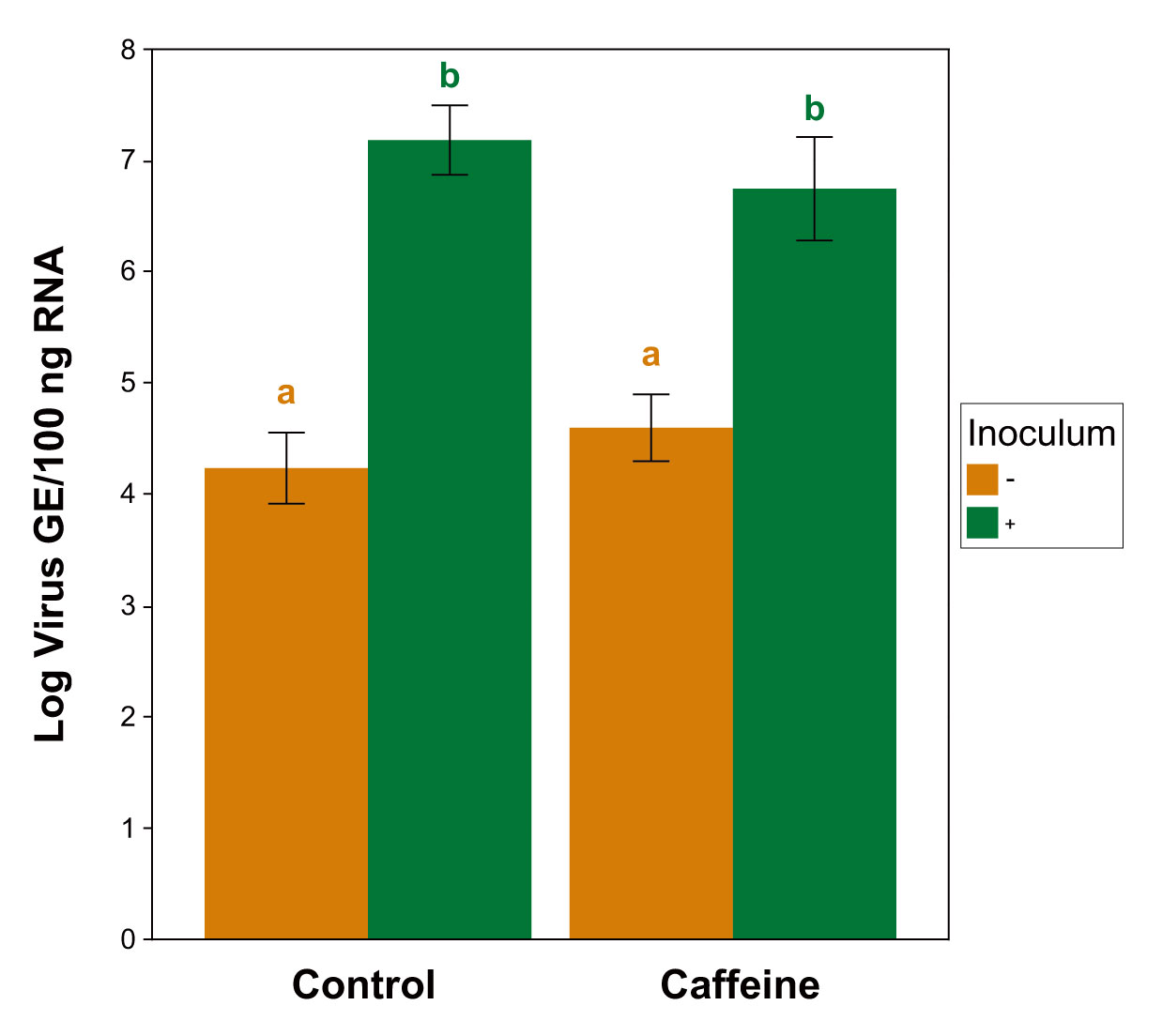
3.2.3. Caffeine Cages Viral Quantification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Smith, T.J.; Saunders, M.E. Honey bees: The queens of mass media, despite minority rule among insect pollinators. Insect Conserv. Divers. 2016, 9, 384–390. [Google Scholar] [CrossRef]
- Aizen, M.A.; Harder, L.D. The Global Stock of Domesticated Honey Bees Is Growing Slower Than Agricultural Demand for Pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Vanengelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Traynor, K.S.; Mondet, F.; De Miranda, J.R.; Techer, M.; Kowallik, V.; Oddie, M.A.; Chantawannakul, P.; McAfee, A. Varroa destructor: A Complex Parasite, Crippling Honey Bees Worldwide. Trends Parasitol. 2020, 36, 592–606. [Google Scholar] [CrossRef]
- Steinhauer, N.A.; Rennich, K.; Wilson, M.E.; Caron, D.M.; Lengerich, E.J.; Pettis, J.S.; Rose, R.; Skinner, J.A.; Tarpy, D.R.; Wilkes, J.T.; et al. A national survey of managed honey bee 2012–2013 annual colony losses in the USA: Results from the Bee Informed Partnership. J. Apic. Res. 2014, 53, 1–18. [Google Scholar] [CrossRef]
- Maori, E.; Paldi, N.; Shafir, S.; Kalev, H.; Tsur, E.; Glick, E.; Sela, I. IAPV, a bee-affecting virus associated with Colony Collapse Disorder can be silenced by dsRNA ingestion. Insect Mol. Biol. 2009, 18, 55–60. [Google Scholar] [CrossRef]
- Hunter, W.B.; Ellis, J.; Vanengelsdorp, D.; Hayes, J.; Westervelt, D.; Glick, E.; Williams, M.; Sela, I.; Maori, E.; Pettis, J.; et al. Large-Scale Field Application of RNAi Technology Reducing Israeli Acute Paralysis Virus Disease in Honey Bees (Apis mellifera, Hymenoptera: Apidae). PLoS Pathog. 2010, 6, e1001160. [Google Scholar] [CrossRef]
- Leonard, S.P.; Powell, J.E.; Perutka, J.; Geng, P.; Heckmann, L.C.; Horak, R.D.; Davies, B.W.; Ellington, A.D.; Barrick, J.E.; Moran, N.A. Engineered symbionts activate honey bee immunity and limit pathogens. Science 2020, 367, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Diet effects on honeybee immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolezal, A.G.; Carrillo-Tripp, J.; Judd, T.M.; Miller, W.A.; Bonning, B.C.; Toth, A.L. Interacting stressors matter: Diet quality and virus infection in honeybee health. R. Soc. Open Sci. 2019, 6, 181803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, P.; Villalobos, E.; Szawarski, N.; Damiani, N.; Gende, L.; Garrido, P.M.; Maggi, M.D.; Quintana, S.; LaMattina, L.; Eguaras, M. Towards Precision Nutrition: A Novel Concept Linking Phytochemicals, Immune Response and Honey Bee Health. Insects 2019, 10, 401. [Google Scholar] [CrossRef] [Green Version]
- Berenbaum, M.R.; Liao, L.-H. Honey Bees and Environmental Stress: Toxicologic Pathology of a Superorganism. Toxicol. Pathol. 2019, 47, 1076–1081. [Google Scholar] [CrossRef]
- Das, L.; Bhaumik, E.; Raychaudhuri, U.; Chakraborty, R. Role of nutraceuticals in human health. J. Food Sci. Technol. 2011, 49, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; Contreras, M.D.M.; Segura-Carretero, A.; Fathi, H.; Nasrabadi, N.N.; Kobarfard, F.; Sharifi-Rad, J. Thymol, thyme, and other plant sources: Health and potential uses. Phytother. Res. 2018, 32, 1688–1706. [Google Scholar] [CrossRef]
- Dauqan, E.M.A.; Abdullah, A. Medicinal and functional values of thyme (Thymus vulgaris L.) herb. J. Appl. Biol. Biotechnol. 2017, 5, 17–22. [Google Scholar]
- Gaur, S. Lipid-Based Nutrient Supplement to Address Child Undernutrition and Enteric Parasitic Infections in India. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Urbana, IL, USA, 2017. [Google Scholar]
- Floris, I.; Satta, A.; Cabras, P.; Garau, V.L.; Angioni, A. Comparison between two thymol formulations in the control of Varroa destructor: Effectiveness, persistence, and residues. J. Econ. Entomol. 2009, 97, 187–191. [Google Scholar] [CrossRef]
- Gregorc, A.; Planinc, I. The control of Varroa destructor in honey bee colonies using the thymol-based acaricide—Apiguard. Am. Bee J. 2005, 145, 672–675. [Google Scholar]
- Palmer-Young, E.C.; Tozkar, C.O.; Schwarz, R.S.; Chen, Y.; E Irwin, R.; Adler, L.S.; Evans, J.D. Nectar and Pollen Phytochemicals Stimulate Honey Bee (Hymenoptera: Apidae) Immunity to Viral Infection. J. Econ. Entomol. 2017, 110, 1959–1972. [Google Scholar] [CrossRef] [PubMed]
- Palmer-Young, E.C.; Sadd, B.M.; Irwin, R.E.; Adler, L.S. Synergistic effects of floral phytochemicals against a bumble bee parasite. Ecol. Evol. 2017, 7, 1836–1849. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Rupasinghe, S.G.; Johnson, R.M.; Zangerl, A.R.; Schuler, M.A.; Berenbaum, M.R. Quercetin-metabolizing CYP6AS enzymes of the pollinator Apis mellifera (Hymenoptera: Apidae). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 154, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera. Proc. Natl. Acad. Sci. USA 2013, 110, 8842–8846. [Google Scholar] [CrossRef] [Green Version]
- Liao, L.-H.; Wu, W.-Y.; Berenbaum, M.R. Impacts of Dietary Phytochemicals in the Presence and Absence of Pesticides on Longevity of Honey Bees (Apis mellifera). Insects 2017, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.J.; Liao, L.-H.; Berenbaum, M.R. Biphasic concentration-dependent interaction between imidacloprid and dietary phytochemicals in honey bees (Apis mellifera). PLoS ONE 2018, 13, e0206625. [Google Scholar] [CrossRef]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. A dietary phytochemical alters caste-associated gene expression in honey bees. Sci. Adv. 2015, 1, e1500795. [Google Scholar] [CrossRef] [Green Version]
- Bernklau, E.; Bjostad, L.B.; Hogeboom, A.; Carlisle, A.; Arathi, S.H. Dietary Phytochemicals, Honey Bee Longevity and Pathogen Tolerance. Insects 2019, 10, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Búrquez, M.D.J.; González-Díaz, F.R.; García-Tovar, C.G.; Carrillo-Miranda, L.; Soto-Zárate, C.I.; Canales-Martínez, M.M.; Penieres-Carrillo, J.G.; Crúz-Sánchez, T.A.; Fonseca-Coronado, S. Comparison between In Vitro Antiviral Effect of Mexican Propolis and Three Commercial Flavonoids against Canine Distemper Virus. Evid.-Based Complement. Altern. Med. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kwon, M.J.; Shin, H.M.; Perumalsamy, H.; Wang, X.; Ahn, Y.-J. Antiviral effects and possible mechanisms of action of constituents from Brazilian propolis and related compounds. J. Apic. Res. 2019, 1–13. [Google Scholar] [CrossRef]
- Grozinger, C.M.; Flenniken, M.L. Bee Viruses: Ecology, Pathogenicity, and Impacts. Annu. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Nathanson, J. Caffeine and related methylxanthines: Possible naturally occurring pesticides. Science 1984, 226, 184–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretschmar, J.A.; Baumann, T.W. Caffeine in Citrus flowers. Phytochemistry 1999, 52, 19–23. [Google Scholar] [CrossRef]
- Wright, G.A.; Baker, D.D.; Palmer, M.J.; Stabler, D.; Mustard, J.A.; Power, E.F.; Borland, A.M.; Stevenson, P.C. Caffeine in Floral Nectar Enhances a Pollinator’s Memory of Reward. Science 2013, 339, 1202–1204. [Google Scholar] [CrossRef] [Green Version]
- Singaravelan, N.; Nee’Man, G.; Inbar, M.; Izhaki, I. Feeding Responses of Free-flying Honeybees to Secondary Compounds Mimicking Floral Nectars. J. Chem. Ecol. 2005, 31, 2791–2804. [Google Scholar] [CrossRef]
- Thomson, J.D.; Draguleasa, M.A.; Tan, M.G. Flowers with caffeinated nectar receive more pollination. Arthropod-Plant Interact. 2015, 9, 1–7. [Google Scholar] [CrossRef]
- Mustard, J.A.; Dews, L.; Brugato, A.; Dey, K.; Wright, G.A. Consumption of an acute dose of caffeine reduces acquisition but not memory in the honey bee. Behav. Brain Res. 2012, 232, 217–224. [Google Scholar] [CrossRef]
- Si, A.; Zhang, S.W.; Maleszka, R. Effects of caffeine on olfactory and visual learning in the honey bee (Apis mellifera). Pharmacol. Biochem. Behav. 2005, 82, 664–672. [Google Scholar] [CrossRef]
- Couvillon, M.J.; Al Toufailia, H.; Butterfield, T.M.; Schrell, F.; Ratnieks, F.L.W.; Schürch, R. Caffeinated forage tricks honeybees into increasing foraging and recruitment behaviors. Curr. Biol. 2015, 25, 2815–2818. [Google Scholar] [CrossRef] [Green Version]
- Balieira, K.V.B.; Mazzo, M.; Bizerra, P.F.V.; Guimarães, A.R.D.J.S.; Nicodemo, D.; Mingatto, F.E. Imidacloprid-induced oxidative stress in honey bees and the antioxidant action of caffeine. Apidologie 2018, 49, 562–572. [Google Scholar] [CrossRef] [Green Version]
- Strachecka, A.; Krauze, M.; Olszewski, K.; Borsuk, G.; Paleolog, J.; Czech, A.; Chobotow, J.; Bajda, M.; Grzywnowicz, K. Unexpectedly strong effect of caffeine on the vitality of western honeybees (Apis mellifera). Biochemistry 2014, 79, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-H.; Wu, C.-P.; Tang, C.-K.; Lin, Y.-H.; Maaroufi, H.O.; Chuang, Y.-C.; Wu, Y.-L. Identification of Immune Regulatory Genes in Apis mellifera through Caffeine Treatment. Insects 2020, 11, 516. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.P.; Pettis, J.S.; Corona, M.; Chen, W.P.; Li, C.; Spivak, M.; Visscher, P.K.; DeGrandi-Hoffman, G.; Boncristiani, H.; Zhao, Y.; et al. Israeli Acute Paralysis Virus: Epidemiology, Pathogenesis and Implications for Honey Bee Health. PLoS Pathog. 2014, 10, e1004261. [Google Scholar] [CrossRef] [PubMed]
- Amiri, E.; Seddon, G.; Smith, W.Z.; Strand, M.K.; Tarpy, D.R.; Rueppell, O. Israeli Acute Paralysis Virus: Honey Bee Queen–Worker Interaction and Potential Virus Transmission Pathways. Insects 2019, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Carrillo-Tripp, J.; Dolezal, A.G.; Goblirsch, M.J.; Miller, W.A.; Toth, A.L.; Bonning, B.C. In vivo and in vitro infection dynamics of honey bee viruses. Sci. Rep. 2016, 6, 22265. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, E.M.; Carrillo-Tripp, J.; Dolezal, A.G. Preparation of Virus-Enriched Inoculum for Oral Infection of Honey Bees (Apis Mellifera). J. Vis. Exp. 2020, 162, e61725. [Google Scholar] [CrossRef]
- De Miranda, J.R.; Bailey, L.; Ball, B.V.; Blanchard, P.; E Budge, G.; Chejanovsky, N.; Chen, Y.-P.; Gauthier, L.; Genersch, E.; De Graaf, D.C.; et al. Standard methods for virus research in Apis mellifera. J. Apic. Res. 2013, 52, 1–56. [Google Scholar] [CrossRef] [Green Version]
- Geffre, A.C.; Gernat, T.; Harwood, G.P.; Jones, B.M.; Gysi, D.M.; Hamilton, A.R.; Bonning, B.C.; Toth, A.L.; Robinson, G.E.; Dolezal, A.G. Honey bee virus causes context-dependent changes in host social behavior. Proc. Natl. Acad. Sci. USA 2020, 117, 10406–10413. [Google Scholar] [CrossRef]
- Raffique, M.K.; Mahmood, R.; Aslam, M.; Sarwar, G. Control of Tropilaelaps clareae mite by using formic acid and thymol in honey bee Apis mellifera L. colonies. Pak. J. Zool. 2012, 44, 1129–1135. [Google Scholar]
- Colin, T.; Lim, M.Y.; Quarrell, S.R.; Allen, G.R.; Barron, A.B. Effects of thymol on European honey bee hygienic behaviour. Apidologie 2019, 50, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Levitt, A.L.; Rajotte, E.G.; Holmes, E.C.; Ostiguy, N.; Vanengelsdorp, D.; Lipkin, W.I.; Depamphilis, C.W.; Toth, A.L.; Cox-Foster, D.L. RNA Viruses in Hymenopteran Pollinators: Evidence of Inter-Taxa Virus Transmission via Pollen and Potential Impact on Non-Apis Hymenopteran Species. PLoS ONE 2010, 5, e14357. [Google Scholar] [CrossRef]
- Dolezal, A.G.; Hendrix, S.D.; Scavo, N.A.; Carrillo-Tripp, J.; Harris, M.A.; Wheelock, M.J.; O’Neal, M.E.; Toth, A.L. Honey Bee Viruses in Wild Bees: Viral Prevalence, Loads, and Experimental Inoculation. PLoS ONE 2016, 11, e0166190. [Google Scholar] [CrossRef]
- Therneau, T.M. Survival: Survival Analysis. Available online: https://cran.r-project.org/web/packages/survival/index.html (accessed on 13 October 2020).
- Kassambara, A.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves Using “ggplot2”. Available online: https://cran.r-project.org/web/packages/survminer/index.html (accessed on 13 October 2020).
- Therneau, T.M. Coxme: Mixed Effects Cox Models. Available online: https://cran.r-project.org/web/packages/coxme/index.html (accessed on 13 October 2020).
- Wickham, H.; François, R.; Henry, L.; Müller, K. Dplyr: A Grammar of Data Manipulation. Available online: https://cran.r-project.org/web/packages/dplyr/index.html (accessed on 13 October 2020).
- Upadhyay, A.; Venkitanarayanan, K. In vivo efficacy of trans-cinnamaldehyde, carvacrol, and thymol in attenuating Listeria monocytogenes infection in a Galleria mellonella model. J. Nat. Med. 2016, 70, 667–672. [Google Scholar] [CrossRef]
- Chang, Y.; Yoon, H.; Kang, D.-H.; Chang, P.-S.; Ryu, S. Endolysin LysSA97 is synergistic with carvacrol in controlling Staphylococcus aureus in foods. Int. J. Food Microbiol. 2017, 244, 19–26. [Google Scholar] [CrossRef]
- Costa, C.; Lodesani, M.; Maistrello, L. Effect of thymol and resveratrol administered with candy or syrup on the development of Nosema ceranae and on the longevity of honeybees (Apis mellifera L.) in laboratory conditions. Apidologie 2009, 41, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Ellis, M.D.; Baxendale, F.P. Toxicity of Seven Monoterpenoids to Tracheal Mites (Acari: Tarsonemidae) and Their Honey Bee (Hymenoptera: Apidae) Hosts When Applied as Fumigants. J. Econ. Entomol. 1997, 90, 1087–1091. [Google Scholar] [CrossRef] [Green Version]
- Schnitzler, P.; Neuner, A.; Nolkemper, S.; Zundel, C.; Nowack, H.; Sensch, K.H.; Reichling, J. Antiviral Activity and Mode of Action of Propolis Extracts and Selected Compounds. Phytother. Res. 2009, 24, S20–S28. [Google Scholar] [CrossRef]
- Johari, J.; Kianmehr, A.; Mustafa, M.R.; Abubakar, S.; Zandi, K. Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus. Int. J. Mol. Sci. 2012, 13, 16785–16795. [Google Scholar] [CrossRef]
- Detzel, A.; Wink, M. Attraction, deterrence or intoxication of bees (Apis mellifera) by plant allelochemicals. Chemoecology 1993, 4, 8–18. [Google Scholar] [CrossRef]
- Dolezal, A.G.; Clair, A.L.S.; Zhang, G.; Toth, A.L.; O’Neal, M.E. Native habitat mitigates feast-famine conditions faced by honey bees in an agricultural landscape. Proc. Natl. Acad. Sci. USA 2019, 116, 25147–25155. [Google Scholar] [CrossRef] [Green Version]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- Wright, G.A.; Nicolson, S.W.; Shafir, S. Nutritional Physiology and Ecology of Honey Bees. Annu. Rev. Entomol. 2018, 63, 327–344. [Google Scholar] [CrossRef]
- Locke, B.; Forsgren, E.; De Miranda, J.R. Increased Tolerance and Resistance to Virus Infections: A Possible Factor in the Survival of Varroa destructor-Resistant Honey Bees (Apis mellifera). PLoS ONE 2014, 9, e99998. [Google Scholar] [CrossRef] [Green Version]
- Laranja, A.T.; Manzatto, A.J.; De Campos Bicudo, H.E.M. Effects of caffeine and used coffee grounds on biological features of Aedes aegypti (Diptera, Culicidae) and their possible use in alternative control. Genet. Mol. Biol. 2003, 26, 419–429. [Google Scholar] [CrossRef]
- Eastep, N.E.; Albert, R.E.; Anderson, J.R. Modulation of La Crosse Virus Infection in Aedes albopictus Mosquitoes Following Larval Exposure to Coffee Extracts. Front. Physiol. 2012, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Njoroge, T. Evaluation of the Role of Plant-Derived Chemicals on Mosquito Ecology and Control. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Urbana, IL, USA, 2020. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Thymol Concentration | IAPV (1%) | |
|---|---|---|---|
| Control–standard | 0 ppm | − | |
| Ad libitum | 160 ppm | − | + |
| Acute Response | 1000 ppm | − | + |
| Prophylactic–Control | 0 ppm; 0 ppm | + | |
| Prophylactic–Inoculated | 160 ppm; 0 ppm | + | |
| Experiment | Description | Figure | Table(s) |
|---|---|---|---|
| Exp. 1 | Carvacrol moderate dose treatments | Figure 1A | Table S1 |
| Exp. 2 | Carvacrol acute dose treatments (1000–2000 ppm) | Figure 1B | Table S2 |
| Exp. 3 | Thymol treatments (acute dose, prophylactic, 160 ppm) | Figure 1C | Table 1 and Table S3 |
| Exp. 4 | Multi-phytochemical trial (quercetin, PCA, caffeine) | Figure 1D | Table S4 |
| Exp. 5 | All 25-ppm caffeine treatments | Figure 2A | Table S5 |
| Exp. 6A | Inoculated caffeine dose-response trial | Figure 2B | Table S6 |
| Exp. 6B | Uninoculated caffeine dose-response trial | Figure 2C | Table S7 |
| Virus Quant. | Viral titers of Experiments 6 and 7 via RT-qPCR | Figure 3 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, E.M.; Berenbaum, M.R.; Dolezal, A.G. Ameliorative Effects of Phytochemical Ingestion on Viral Infection in Honey Bees. Insects 2020, 11, 698. https://doi.org/10.3390/insects11100698
Hsieh EM, Berenbaum MR, Dolezal AG. Ameliorative Effects of Phytochemical Ingestion on Viral Infection in Honey Bees. Insects. 2020; 11(10):698. https://doi.org/10.3390/insects11100698
Chicago/Turabian StyleHsieh, Edward M., May R. Berenbaum, and Adam G. Dolezal. 2020. "Ameliorative Effects of Phytochemical Ingestion on Viral Infection in Honey Bees" Insects 11, no. 10: 698. https://doi.org/10.3390/insects11100698
APA StyleHsieh, E. M., Berenbaum, M. R., & Dolezal, A. G. (2020). Ameliorative Effects of Phytochemical Ingestion on Viral Infection in Honey Bees. Insects, 11(10), 698. https://doi.org/10.3390/insects11100698