A Survey of the Brazilian Dicranocentrus Schött (Collembola, Orchesellidae, Heteromurini) with the Description of a New Species and Notes on the Genus



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
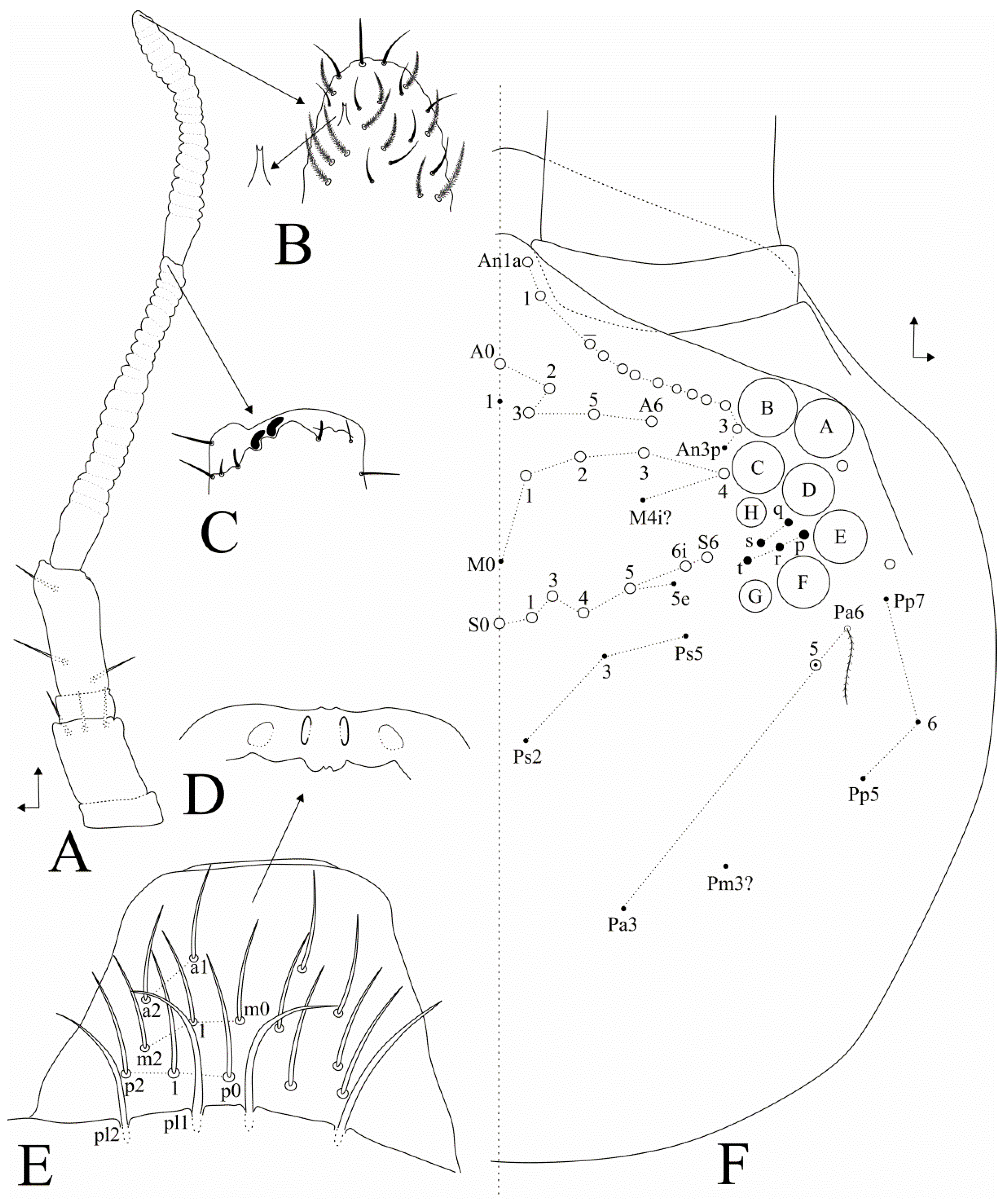
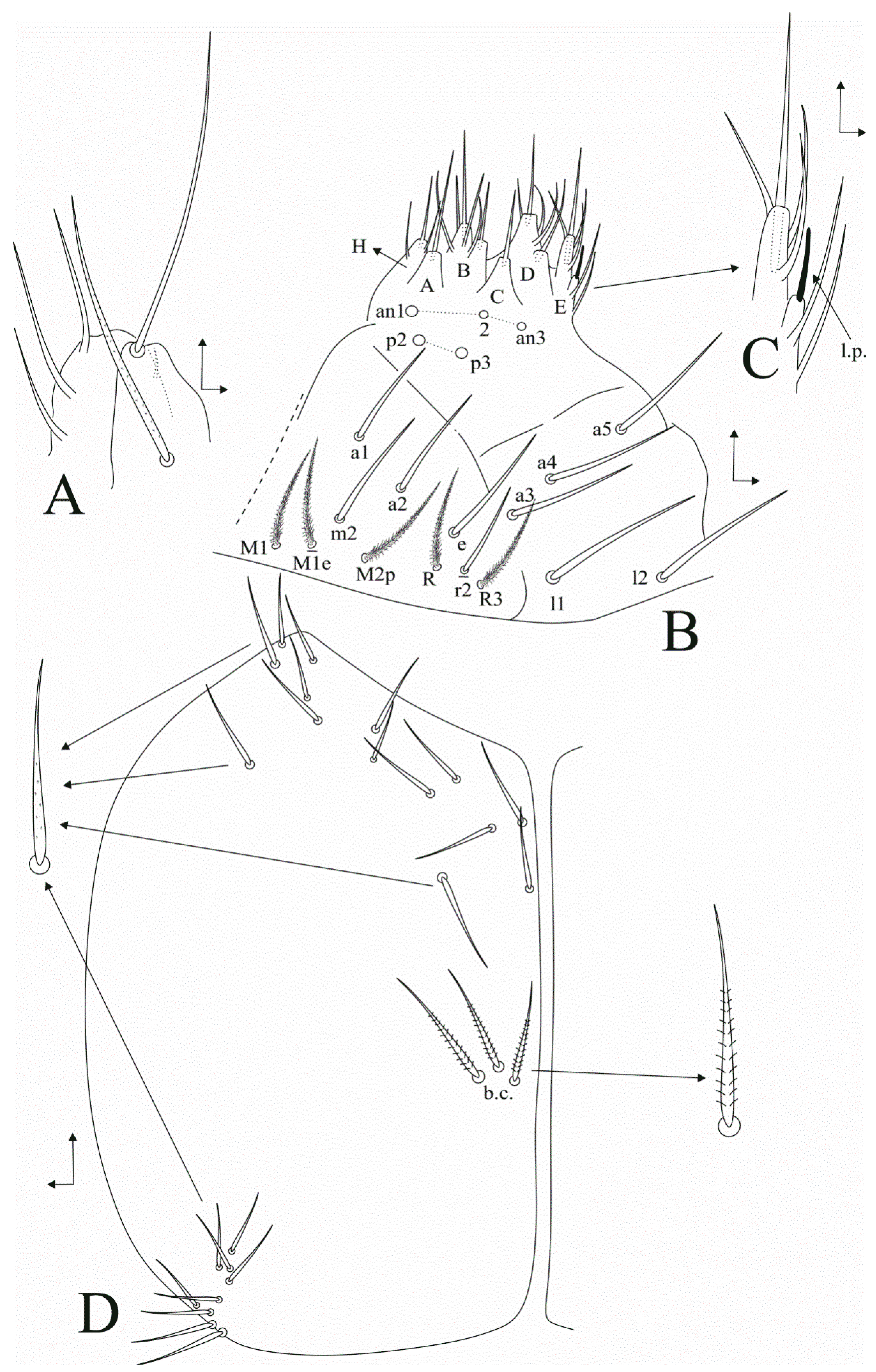
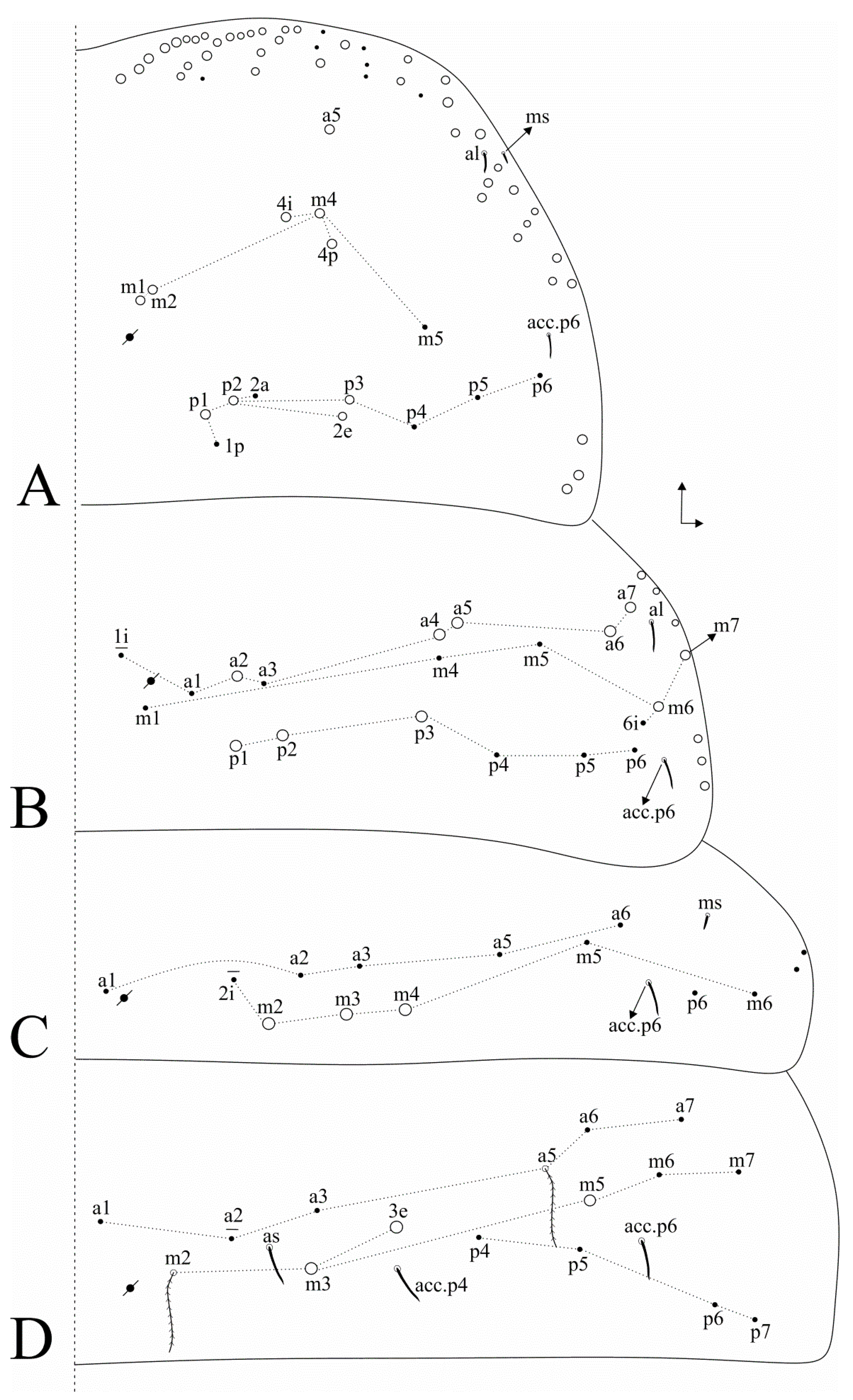
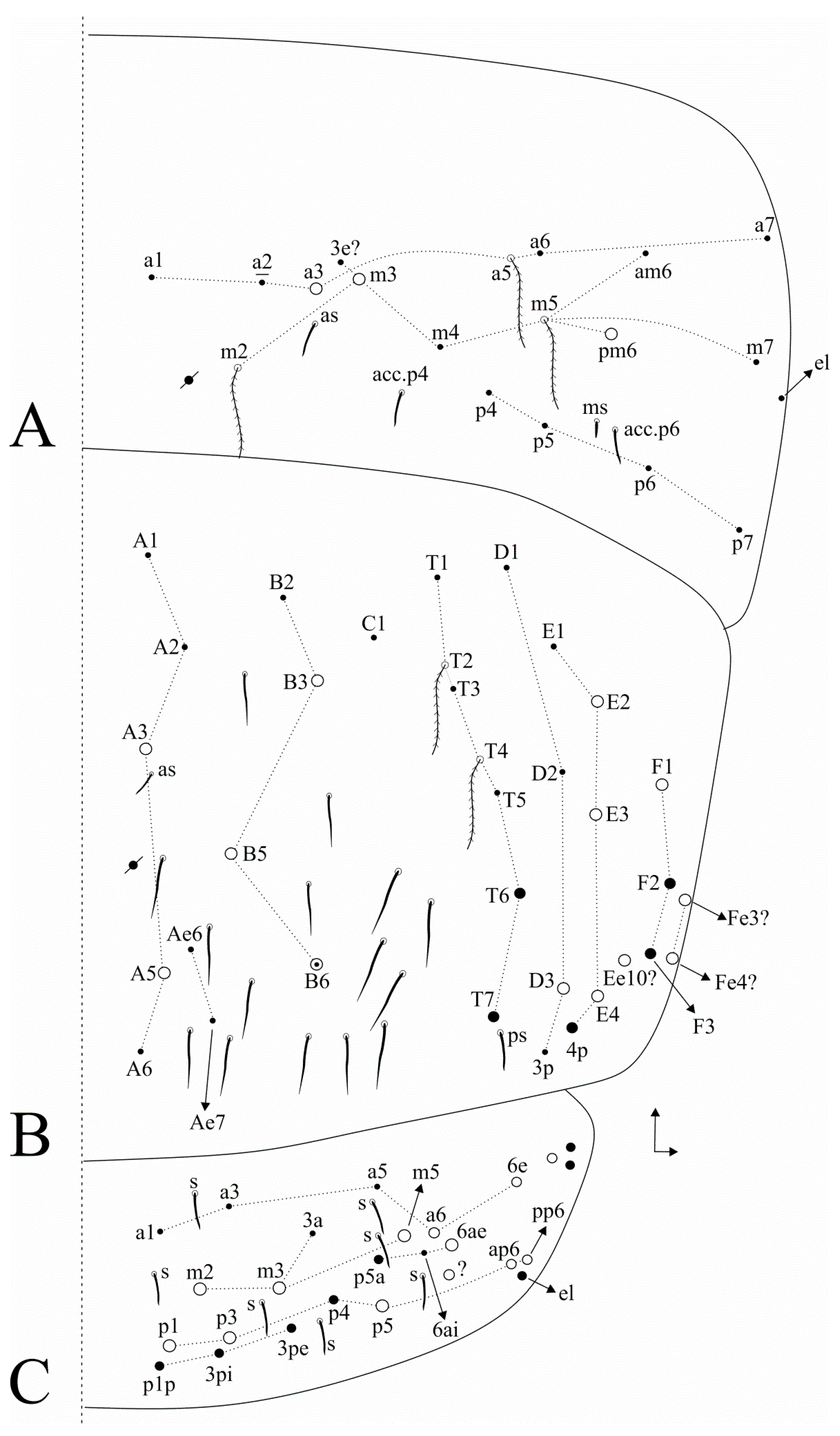
3.1. Taxonomic Summary and Genus Diagnosis
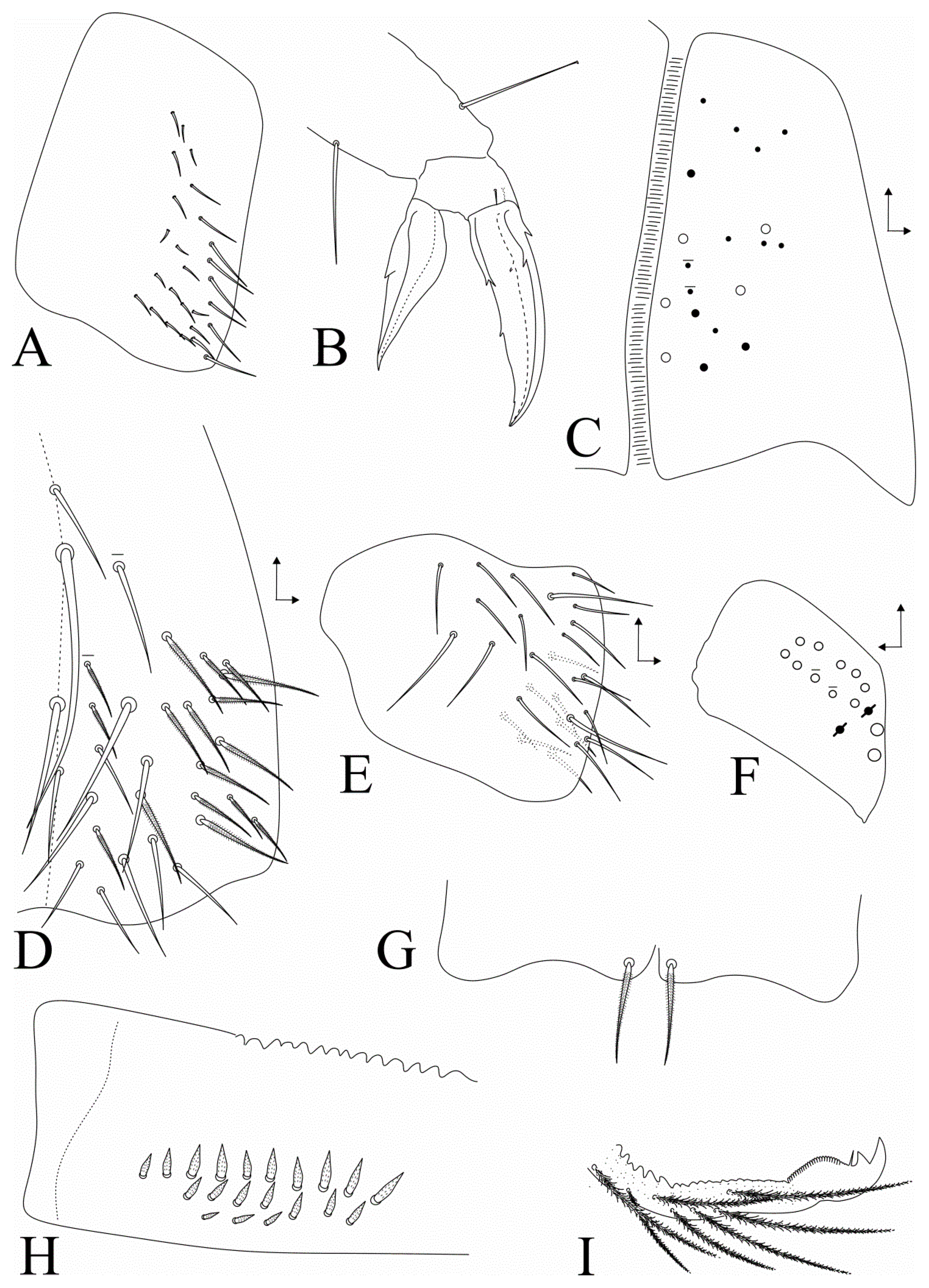
3.2. Dicranocentrus abestado sp. nov. Siqueira, Bellini, and Cipola
4. Discussion
4.1. Brazilian Species of Dicranocentrus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species [References] | gemellus | indicus | inermodentes | luzonensis | nepalensis | solomonensis | wangi | |
|---|---|---|---|---|---|---|---|---|
| [50] | [12,51,54,60] | [12,44,52] | [50] | [53] | [12,44] | [54] | ||
| Type Locality | Finschhafen, | Bandra, | Saipan, | Baguio, | Chainpur, | Guadalcanal, | Guangzhou, | |
| Papua New Guinea | India | Northern Mariana Is. | Philippine Is. | Nepal | Solomon Is. | China | ||
| Characteristics | Species Group | sundanensis | sundanensis? | sundanensis | sundanensis | gracilis | sundanensis | sundanensis |
| Body bluish/violetish | entire body or just | Ant., lateral Th. II | Ant., anterior head | Ant. and legs | Ant. and | entire body, except | Ant., distal | |
| pigmentation on | antennae and legs | and posterior trunk segments | and legs | (except femurs) | anterior head | dorsal head and | femurs and | |
| segments | furcula | tibiotarsi | ||||||
| Interocular chaetae | 4–5 | ? | ? | 3 | ? | ? | ? | |
| Dorsal head mac | S2 | + | ? | + | + | ‒ | + | + |
| Pa1 | + | ? | + | ‒ | + | + | ‒ | |
| Pa2 | + | ? | + | + | + | + | + | |
| Pp3 | + | ? | + | ‒ | + | + | + | |
| Pp5 | + | ? | + | + | ‒ | + | + | |
| Labral papillae | inner | hooked | ? | conical | conical | conical | conical | conical |
| outer | hooked | ? | conical | rounded | conical | conical | conical | |
| Labial basomedian field | M1 | S | ? | C | C | C | S | C |
| M2 | S | ? | C | S | C | S | S | |
| R | S | ? | S/C | S | C | S | S | |
| extra chaetae | 2–3 S | ? | 0–1 S, 3–4 C | 0–2 S, 1–2 C | 2 C | 4–6 S | 0–1 S, 2 C | |
| Th. II mac | p1–3 group | 8 | 8 | 7 | 8 | 6 | 8 | 7–9 |
| p5 mac | ‒ | ‒ | + | ‒ | ‒ | ‒ | + | |
| Th. III central mac | a, m, p rows | 5,1,3 | 5,1,3 | 5,1,3 | 5,1,3 | 5,1,3 | 5,1,5 | 5,2,5 |
| Abd. IV central mac | 4–5 | ? | 3 | 3 | 4 | 4–5 | 4–5 | |
| Trochanteral organ chaetae | 30–36 | 29 | ? | 24–37 | ? | ? | 25–41 | |
| Tibiotarsus III | inner | +(several) | ‒ | ‒ | 15–20 | ‒ | +? | 0–2 |
| smooth chaetae | outer | +(several) | ‒ | ‒ | 1–2 | ‒ | +? | 5–11 |
| Tenent hair | capitate/acuminate | capitate | capitate | acuminate | capitate | acuminate | acuminate | |
| Unguis teeth | ratio | b.t. = m.t.>a.t. | b.t. > m.t. | b.t. = m.t. | b.t. > m.t. = a.t. | b.t. > m.t. = a.t. | b.t. = m.t. | b.t. > m.t. |
| a.t. | +/‒ | – | ‒ | +/‒ | + | ‒ | ‒ | |
| Unguiculus | pe | 0–1 tooth | smooth | smooth | 1 tooth | 1 tooth | 1 tooth | 0–1 tooth |
| Tenaculum chaetae chaetae | 3–4 | 1 | ? | 5–7 | ? | ? | 4–10 | |
| Manubrium dorsal | number | 3–4 | ‒ | ‒ | 5–6 | ‒ | 5 | 23–25 |
| smooth chaeta | distribution | 2 rows | ‒ | ‒ | 2 rows | ‒ | 2 rows | 3 rows |
| Dens (basal) smooth chaeta | 1 | ? | ‒ | 1–3 | ‒ | 1 | 1–4 | |
| Dens (dorsal) blunt mac | ‒ | + | ‒ | ‒ | ‒ | ‒ | ‒ | |
| Dental spines | number | ‒ | ‒ | ‒ | 14–20 | ‒ | ‒ | 8–13 |
| distribution | ‒ | ‒ | ‒ | 1 row | ‒ | ‒ | 1 row | |
| Species [references] | assimilis | inermis | javanus | liuae | meruensis | simplex | |
|---|---|---|---|---|---|---|---|
| [12,44,55] | [12,55] | [56] | [57] | [12,44,58] | [56,59] | ||
| Type Locality | Debundscha, | Debundscha, | Bogor, West | Shitai, | Meru, | Bukit Timah, | |
| Cameroon | Cameroon | Java, Indonesia | China | Kenya | Singapore | ||
| Characteristics | Species Group | gracilis | gracilis | sundanensis | sundanensis | gracilis | sundanensis |
| Body bluish/violetish | Entire body, except | Ant., legs and | Ant. only | Ant., femurs and | Entire body or | Ant., legs and | |
| pigmentation on | dorsal head and trunk | Abd. V–VI | tibiotarsi | just Ant. and legs | ventral body | ||
| and ventral dens | |||||||
| Dorsal head mac | S2 | ‒ | ‒ | + | + | ‒ | + |
| Pa1 | ‒ | ‒ | + | + | ‒ | ‒ | |
| Pp3 | + | ‒ | + | + | + | + | |
| Pp5 | + | ‒ | + | + | ‒ | + | |
| Labial basomedian field | M1 | ? | ? | S | C | C | ? |
| M2 | ? | ? | S | S | S | ? | |
| R | ? | ? | C | S | S | ? | |
| extra chaetae | ? | ? | 3 C | 4 S | 6 C | ? | |
| Th. II mac | m4 group | 3 | 2 | 2 | 2 | 2 | 2 |
| p1–3 group | 6 | 7 | 6 | 8 | 6 | 6–7 | |
| p5 mac | ‒ | ‒ | + | + | ‒ | ‒ | |
| Th. III central mac | a, m, p rows | 4,2,3 | 4,2,3 | 5,1,3 | 5,1,3–4 | 4,2,3 | 4,0,3 |
| Trochanteral organ chaetae | ? | ? | 50 | 30 | ? | 70 | |
| Tibiotarsus III | inner | +? | +? | ‒ | +? | +? | ? |
| smooth chaetae | outer | ‒ | +? | ‒ | ‒? | ‒ | ? |
| Tenent hair | acuminate | capitate | acuminate | acuminate | acuminate | acuminate | |
| Unguis teeth | ratio | b.t. > m.t. | b.t. = m.t. | b.t. > m.t. | b.t. = m.t. | b.t.=m.t. | b.t. only * |
| m.t. | + | + | + | + | + | ‒? | |
| Unguiculus lamellae | ai | acuminate | truncate, 1 tooth | acuminate | acuminate | acuminate | acuminate |
| pe | 1 tooth | 1 tooth | smooth | smooth | 1 tooth | smooth | |
| Tenaculum chaetae | ? | ? | 2 | 2–4 | ? | ? | |
| Manubrium dorsal | number | ? | ? | 6 | +? | +? | 3 |
| smooth chaeta | distribution | ? | ? | 2 rows | +? | +? | 2 rows |
| Dens (basal) smooth chaeta | ? | ? | 1 | 5 | +? | 2 | |
| Dens (dorsal) blunt mac | ‒ | ‒ | + | ‒ | ‒ | ‒ | |
| Dental spines | number | ‒ | ‒ | ‒ | ‒ | ±40–46 | 5–8 |
| distribution | ‒ | ‒ | ‒ | ‒ | 3–4 rows | 1 row | |
4.2. Notes on Dicranoncentrus Chaetotaxy
4.3. Notes on Dicranocentrus Species Groups and Related Genera

4.4. Notes on Heteromurinae Systematics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Womersley, H. A preliminary account of the Collembola—Arthropleona of Australia II (Entomobryoidea). Trans. R. Soc. S. Aust. 1934, 58, 86–138. [Google Scholar]
- Zhang, F.; Bellini, B.C.; Soto-Adames, F.N. New insights into the systematics of Entomobryoidea (Collembola: Entomobryomorpha): First instar chaetotaxy, homology and classification. Zool. Syst. 2019, 44, 249–278. [Google Scholar] [CrossRef]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 31 August 2020).
- Börner, C. Das System der Collembolen-nebst Beschreibung neuer Collembolen des Hamburger Naturhistorischen Museums. Mit. aus dem Nat. Mus. Hambg. 1906, 23, 147–188. [Google Scholar]
- Schäffer, C. Die Collembolen der Umgebung von Hamburg und benachtbaren Gebiete. Mit. aus dem Nat. Mus. Hambg. 1896, 13, 147–216. [Google Scholar]
- Zeppelini, D.; Queiroz, G.C.; Bellini, B.C. Collembola in Catálogo Taxonômico da Fauna do Brasil. PNUD. Available online: http://fauna.jbrj.gov.br/fauna/faunadobrasil/379/ (accessed on 31 August 2020).
- Schött, H. Beiträge zur kenntnis der insektenfauna von Kamerun, Bihang till K. Sven. Vetenskapsakad. Handl. Stockh. 1893, 19, 5–28. [Google Scholar]
- Templeton, R.; Westwood, J.O. Thysanurae Hibernicae, or descriptions of such species of spring-tailed insects (Podura and Lepisma, Linn.,) as have been observed in Ireland. Descriptions of the Irish Species of Thysanura. Trans. Entomol. Soc. Lond. 1836, 1, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Mari-Mutt, J.A.; Bellinger, P.F. A Catalog of Neotropical Collembola, Including Nearctic Areas of Mexico. In Flora and Fauna Handbook 5, 1st ed.; Sandhill Crane Press: Gainesville, FL, USA, 1990; pp. 1–237. [Google Scholar]
- Mari-Mutt, J.A.; Bellinger, P.F. Supplement to the catalog of the Neotropical Collembola. Caribb. J. Sci. 1996, 32, 166–175. [Google Scholar]
- Mari-Mutt, J.A.; Bellinger, P.F.; Janssens, F. Checklist of the Collembola: Supplement to the Catalog of the Neotropical Collembola. Available online: http://www.collembola.org/publicat/neotrcat.htm (accessed on 31 August 2020).
- Mari-Mutt, J.A. A revision of the genus Dicranocentrus Schött (Insecta: Collembola: Entomobryidae). Agric. Exp. Sta. Bull. 1979, 259, 1–79. [Google Scholar]
- Handschin, E. Neue myrmecophile und termitophile Collembolenformen aus Süd Amerika. Neue Beitr. Syst. Insektenk. Berlin 1924, 3, 13–28. [Google Scholar]
- Cipola, N.G.; Oliveira, F.G.L.; Morais, J.W.; Bellini, B.C. The Heteromurini Absolon & Ksenemann (Collembola, Entomobryidae): A review of the genera status and diagnoses, keys for species of Alloscopus Börner and Heteromurtrella Mari-Mutt and description of a new species. Zootaxa 2016, 4084, 151–186. [Google Scholar] [CrossRef]
- Zhang, F.; Cipola, N.G.; Pan, Z.-X.; Ding, Y. New insight into the systematics of Heteromurini (Collembola: Entomobryidae: Heteromurinae) with special reference to Alloscopus and Sinodicranocentrus gen. n. Arth. Syst. Phylogeny 2020, 78, 1–16. [Google Scholar] [CrossRef]
- Arlé, R.; Mendonça, C. Estudo preliminar das espécies de Dicranocentrus Schött, 1893, ocorrentes no Parque Nacional da Tijuca, Rio de Janeiro (Collembola). Rev. Brasil. Biol. 1982, 42, 41–49. [Google Scholar]
- Bellini, B.C.; Morais, J.W.; Oliveira, F.G.L. A new species of Dicranocentrus Schött (Collembola, Entomobryidae, Orchesellinae) from Brazilian Amazon. Zootaxa 2013, 3709, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Xisto, T.; Mendonça, M.C. Redescription of Dicranocentrus heloisae Arlé & Mendonça, 1982 (Collembola: Entomobryidae). J. Insect Biodivers. 2014, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Xisto, T.; Mendonça, M.C.; Abrantes, E.A. Dicranocentrus heloisae Arlé & Mendonça, 1982 (Collembola: Entomobryidae): First record of the genus in Espírito Santo state, Brazil. Check List 2015, 11, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Xisto, T.; Mendonça, M.C. Two new species of Dicranocentrus Schött, 1893 (Collembola: Entomobryidae) from Serra do Gandarela, Minas Gerais State, Brazil. Zootaxa 2016, 4079, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Xisto, T.; Mendonça, M.C. New species and new records of Dicranocentrus Schött, 1893 (Collembola: Entomobryidae) from Southeastern Brazil. Insect Syst. Evol. 2017, 49, 23–58. [Google Scholar] [CrossRef]
- Absolon, K.; Ksenemann, M. Troglopedetini. Vergleichende Studie über eine altertümliche höhlenbewohnende Kollembolengruppe aus den dinarischen Karstgebieten. Stud. Geb. Karst. Höhlenkunde Eiszeitforsch. Nachbargeb. 1942, 16, 5–57. [Google Scholar]
- Mari-Mutt, J.A. Heteromurtrella, a new tropical subgenus of Heteromurus with descriptions of two new species (Insecta: Collembola: Entomobryidae). J. Agric. Univ. Puerto Rico 1979, 63, 214–222. [Google Scholar]
- Absolon, K. Untersuchungen über Apterygoten auf Grund der Sammlungen des Wiener Hofmuseums. Ann. des K.K. Naturhist. Hofmus. 1903, 18, 91–111. [Google Scholar]
- Mari-Mutt, J.A. A classification of the Orchesellinae with a key to tribes, genera and subgenera (Collembola: Entomobryidae). Ann. Entomol. Soc. Am. 1980, 73, 455–459. [Google Scholar] [CrossRef]
- Zhang, F.; Deharveng, L. Systematic revision of Entomobryidae (Collembola) by integrating molecular and new morphological evidence. Zool. Scr. 2015, 44, 298–311. [Google Scholar] [CrossRef]
- Soto-Adames, F.N.; Barra, J.A.; Christiansen, K.; Jordana, R. Suprageneric Classification of Collembola Entomobryomorpha. Ann. Entomol. Soc. Am. 2008, 101, 501–513. [Google Scholar] [CrossRef]
- Nunes, R.C.; Santos-Costa, R.C.; Bellini, B.C. The first Neotropical Capbrya Barra, 1999 (Collembola: Orchesellidae: Nothobryinae) and the reinterpretation of Nothobryinae systematics. Zool. Anz. 2020, 288, 24–42. [Google Scholar] [CrossRef]
- Arlé, R. Novas espécies de colêmbolas aquáticas (Nota preliminar). Atas Soc. Biol. Rio de Janeiro 1961, 5, 34–37. [Google Scholar]
- Silveira, T.C.; Mendonça, M.C. New species of Nothobrya (Collembola: Entomobryidae) from Southeast Brazil. Zoologia 2016, 33, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Nunes, R.C.; Bellini, B.C. A new species of Nothobrya Arlé, 1961 (Collembola: Entomobryidae) from Brazil and notes on key characters for Nothobryinae taxonomy, with an identification key to the species of the subfamily. Zootaxa 2019, 4615, 375–391. [Google Scholar] [CrossRef]
- Barra, J.A. Un nouveau genre Capbrya avec deux nouvelles espèces de la Province du Cap (Rép. Sud Africaine) (Collembola: Entomobryidae). Capbrya n. gen. d’Afrique du Sud (Insecta: Collembola). Bull. Inst. R. Sci. Nat. Belg. 1999, 69, 19–24. [Google Scholar]
- Jordana, R.; Arbea, J.I.; Simón, C.; Luciáñez, M.J. Fauna Iberica Volume 8: Collembola, Poduromorpha, 1st ed.; Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas: Madrid, Spain, 1997; pp. 1–807. [Google Scholar]
- Fjellberg, A. The Labial Palp in Collembola. Zool. Anz. 1999, 237, 309–330. [Google Scholar]
- Gisin, H. Espèces nouvelles et lignées évolutives de Pseudosinella endogés. Mem. Est. Mus. Zool. Univ. Coimbra 1967, 301, 5–25. [Google Scholar]
- Zhang, F.; Pan, Z.-X. Homology of labial chaetae in Entomobryoidea (Collembola). Zootaxa 2020, 4766, 498–500. [Google Scholar] [CrossRef]
- Cipola, N.G.; Morais, J.W.; Bellini, B.C. A new species of Seira (Collembola: Entomobryidae: Seirini) from Northern Brazil, with the addition of new chaetotaxic characters. Zoologia 2014, 31, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Soto-Adames, F.N. Postembryonic development of the dorsal chaetotaxy in Seira dowlingi (Collembola, Entomobryidae); with an analysis of the diagnostic and phylogenetic significance of primary chaetotaxy in Seira. Zootaxa 2008, 1683, 1–31. [Google Scholar] [CrossRef]
- Szeptycki, A. Morpho-systematic studies on Collembola. III. Body chaetotaxy in the first instars of several genera of the Entomobryomorpha. Acta Zool. Cracov. 1972, 17, 341–372. [Google Scholar]
- Szeptycki, A. Chaetotaxy of the Entomobryidae and its Phylogenetical Significance. Morpho-Systematic Studies on Collembola; Polska Akademia Nauk: Kraków, Poland, 1979; Volume IV, pp. 1–219. [Google Scholar]
- Zhang, F.; Yu, D.; Stevens, M.I.; Ding, Y. Colouration, chaetotaxy and molecular data provide species-level resolution in a species complex of Dicranocentrus (Collembola: Entomobryidae). Invertebr. Syst. 2018, 32, 1298–1315. [Google Scholar] [CrossRef]
- Börner, C. Die Familien der Collembolen. Zool. Anz. 1913, 41, 315–322. [Google Scholar]
- Xisto, T.; Mendonça, M.C. Contribuição ao conhecimento da morfologia e taxonomia de Dicranocentrus Schött, 1893, com histórico comentado para a Região Neotropical (Collembola: Entomobryidae). In Anais do III Simpósio de Entomologia do Rio de Janeiro, 1st ed.; Da-Silva, E.R., Passos, M.I.S., Aguiar, V.M., Lessa, C.S.S., Coelho, L.B.N., Eds.; Universidade Federal do Estado do Rio de Janeiro (UNIRIO): Rio de Janeiro, Brazil, 2016; pp. 151–165. [Google Scholar]
- Mari-Mutt, J.A. A new species and complements to the descriptions of nine African or Oriental Dicranocentrus (Collembola: Entomobryidae). J. Agric. Univ. Puerto Rico 1981, 65, 160–170. [Google Scholar]
- Mari-Mutt, J.A. New genus, a new species, and complements to the descriptions of seven Neotropical Dicranocentrus (Collembola: Entomobryidae: Orchesellinae). J. Agric. Univ. Puerto Rico 1981, 65, 90–107. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Mari-Mutt, J.A. Four new species of Dicranocentrus and Heteromurus from the State of Mérida, Venezuela (Collembola: Entomobryidae). J. Agric. Univ. Puerto Rico 1983, 67, 148–164. [Google Scholar]
- Soto-Adames, F.N.; Anderson, E.W. Two new species and new records of springtails (Collembola: Entomobryidae, Paronellidae) from Nevis, Lesser Antilles. Fla. Entomol. 2017, 100, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Wray, D.L. New Collembola from Puerto Rico. J. Agric. Univ. Puerto Rico 1953, 37, 140–150. [Google Scholar]
- Mari-Mutt, J.A. Eight new species of Dicranocentrus and redescriptions for D. thaicus and D. pilosus (Collembola: Entomobryidae: Orchesellinae). J. Agric. Univ. Puerto Rico 1985, 69, 297–322. [Google Scholar]
- Bonet, F. Sur quelques Collemboles de l’Inde. Eos Rev. Esp. Entomol. 1930, 6, 249–273. [Google Scholar]
- Uchida, H. Collembola Von Mikronisien. Bull. Tokyo Sci. Mus. 1944, 17, 1–23. [Google Scholar]
- Mari-Mutt, J.A.; Bhattacharjee, R.K. Four new species of Dicranocentrus from Northeast India and Nepal. Pac. Insects 1980, 22, 162–170. [Google Scholar]
- Ma, Y.; Chen, J.-X. A new Dicranocentrus species (Collembola: Entomobryidae) from China with a key to all species in the genus from Asia. Zootaxa 2007, 1633, 63–68. [Google Scholar] [CrossRef]
- Schött, H. Kamerunische Collembolen. Linköping 1927, 5, 1–40. [Google Scholar]
- Yoshii, R.; Suhardjono, Y.R. Notes on the Collembolan Fauna of Indonesia and its vicinities. I. Miscellaneous Notes, with special references to Seirini and Lepidocyrtini. Acta Zool. Asiae Orient. 1989, 1, 23–90. [Google Scholar]
- Xu, G.-L.; Zhang, F. A new species of Dicranocentrus from China with comments on the systematic position of the genus. ZooKeys 2014, 417, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wahlgren, E. Übersicht der aus dem Kilimandjaro-Mern-Gebiete bisher bekannten Collembolen. Sjöstedts Kilimandjaro-Meru Exp. 1908, 18, 1–10. [Google Scholar]
- Yosii, R. Studies on the Collembolan Fauna of Malay and Singapore with special reference to the genera: Lobella, Lepidocyrtus and Callyntrura. Contrib. Biol. Lab. Kyoto Univ. 1959, 10, 1–65. [Google Scholar]
- Yosii, R. On some Collembola of Afghanistan, India and Ceylon, collected by the Kuphe-Expedition, 1960. R. Kyoto U. Sci. Exp. Karakoram Hindukush 1955 1966, 8, 333–405. [Google Scholar]
- Cassagnau, P. Collemboles d’Amérique du Sud, II. Orchesellini, Paronellinae, Cyphoderinae. Biol. Amérique Australe 1963, 2, 127–148. [Google Scholar]
- Bellini, B.C. Fauna de Collembola (Arthropoda) em áreas úmidas do semiárido. In Artrópodes do Semiárido: Biodiversidade e Conservação, 1st ed.; Bravo, F., Calor, A., Eds.; Printmídia: Feira de Santana, Brazil, 2014; pp. 57–68. [Google Scholar]
- Abrantes, E.A.; Bellini, B.C.; Bernardo, A.N.; Fernandes, L.H.; Mendonça, M.C.; Oliveira, E.P.; Queiroz, G.C.; Sautter, K.D.; Silveira, T.C.; Zeppelini, D. Errata Corrigenda and update for the “Synthesis of Brazilian Collembola: An update to the species list.” ABRANTES et al. (2010) Zootaxa, 2388: 1–22. Zootaxa 2012, 3168, 1–21. [Google Scholar] [CrossRef]
- Bellini, B.C.; Godeiro, N.N. Novos registros de Collembola (Arthropoda, Hexapoda) para áreas úmidas do semiárido do Brasil. In Artrópodes do Semiárido II: Biodiversidade e conservação, 1st ed.; Bravo, F., Ed.; Métis: São Paulo, Brazil, 2017; pp. 28–53. [Google Scholar]
- Denis, J.R. Contributo alla conoscenza del “Microgenton” di Costa Rica. II –Collemboles de Costa Rica avec une contribution au species de l’ordre. Boll. Lab. Entomol. Agric. 1931, 25, 69–170. [Google Scholar]
- Bonet, F. Colémbolos de La República Argentina. Eos Rev. Esp. de Entomol. 1934, 9, 123–194. [Google Scholar]
- Bellini, B.C.; Cipola, N.G. The Neotropical genera of Paronellinae (Collembola, Entomobryoidea, Paronellidae) with description of two new species and redescription of Campylothorax mitrai. Zootaxa 2017, 4300, 151–179. [Google Scholar] [CrossRef]
- Prabhoo, N.R. Soil and litter Collembola of South India I-Arthropleona. Orient. Insects 1971, 5, 1–46. [Google Scholar] [CrossRef]
- Yosii, R. Collembola of Khumbu Himal. Khumbu Himal 1971, 4, 80–130. [Google Scholar]
- Mandal, G.P. A new genus of Heteromurini (Collembola: Entomobryidae) with dental base falcate macrochaetae, from India. Halteres 2018, 9, 74–85. [Google Scholar]
- Christiansen, K.A.; Bellinger, P. Biogeography of Hawaiian Collembola: The simple principles and complex reality. Orient. Insects 1994, 28, 309–351. [Google Scholar] [CrossRef]
- Zhang, F.; Sun, D.; Yu, D.; Wang, B. Molecular phylogeny supports S-chaetae as a key character better than jumping organs and body scales in classification of Entomobryoidea (Collembola). Sci. Rep. 2015, 5, 12471. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Pan, Z.; Wu, J.; Ding, Y.; Yu, D.; Wang, B. Dental scales could occur in all scaled subfamilies of Entomobryidae (Collembola): New definition of Entomobryinae with description of a new genus and three new species. Invertebr. Syst. 2016, 30, 598–615. [Google Scholar] [CrossRef]
- Zhang, F.; Ma, Y.; Greenslade, P. New Australian Paronellidae (Collembola) reveal anomalies in existing tribal diagnoses. Invertebr. Syst. 2017, 31, 375–393. [Google Scholar] [CrossRef]
- Nunes, R.C.; Godeiro, N.N.; Pacheco, G.; Liu, S.; Gilbert, M.T.P.; Alvarez-Valin, F.; Zhang, F.; Bellini, B.C. The discovery of Neotropical Lepidosira (Collembola, Entomobryidae) and its systematic position. Zool. Scr. 2019, 48, 783–800. [Google Scholar] [CrossRef]
- Santos, N.M.C.; Santos-Costa, R.C.; Siqueira, O.J.R.; Godeiro, N.N.; Bellini, B.C. Two new species of Entomobrya Rondani (Collembola, Entomobryidae) from northeastern Brazil and comments on the genus. Zootaxa 2020, 4731, 43–62. [Google Scholar] [CrossRef]
- Godeiro, N.N.; Pacheco, G.; Liu, S.; Cipola, N.G.; Berbel-Filho, W.M.; Zhang, F.; Gilbert, M.T.P.; Bellini, B.C. Phylogeny of Neotropical Seirinae (Collembola, Entomobryidae) based on mitochondrial genomes. Zool. Scr. 2020, 49, 329–339. [Google Scholar] [CrossRef]






| Taxa | Known Distribution in Brazil (Biomes) |
|---|---|
| Heteromurinae Absolon and Ksenemann, 1942 [22] | |
| Heteromurini Absolon and Ksenemann, 1942 [22] | |
| Heteromurtrella Mari-Mutt, 1979 [23] | |
| H. anae Cipola, 2016 * [14] | Amazon Forest |
| Dicranocentrus Schött, 1893 [7] | |
| D. albicephalus Xisto and Mendonça, 2017 * [21] | Atlantic Forest |
| D. amazonicus Bellini, Moraes and Oliveira, 2013 * [17] | Amazon Forest |
| D. bicolor Handschin, 1924 *,*** [13] | Atlantic Forest |
| D. cuprum Xisto and Mendonça, 2016 * [20] | Atlantic Forest |
| D. heloisae Arlé and Mendonça, 1982 * [16] | Atlantic Forest |
| D. magnus Xisto and Mendonça, 2017 * [21] | Atlantic Forest |
| D. marimutti Xisto and Mendonça, 2017 * [21] | Atlantic Forest |
| D. melinus Xisto and Mendonça, 2016 * [20] | Atlantic Forest |
| D. pikachu Xisto and Mendonça, 2017 * [21] | Atlantic Forest |
| D. silvestrii Absolon, 1903 **,*** [24] | Atlantic Forest |
| D. termitophilus Handschin, 1924 *,*** [13] | Atlantic Forest? |
| Mastigocerini Mari-Mutt, 1980 [25] | |
| Mastigoceras Handschin, 1924 * [13] | |
| M. camponoti Handschin, 1924 *,** [13] | Atlantic Forest, Amazon Forest, Caatinga |
| Nothobryinae Zhang and Deharveng, 2015 [26] | |
| Nothobryini Soto-Adames et al., 2008 [27]sensu Nunes et al. 2020 [28] | |
| Nothobrya Arlé, 1961 * [29] | |
| N. arlei Silveira and Mendonça, 2016 *[30] | Atlantic Forest |
| N. schubarti Arlé, 1961 * [29] | Cerrado, Caatinga |
| N. sertaneja Nunes and Bellini, 2019 * [31] | Caatinga |
| Capbryini Nunes et al. 2020 [28] | |
| Capbrya Barra, 1999 [32] | |
| Capbrya brasiliensis Nunes, Santos-Costa and Bellini, 2020 * [28] | Caatinga, Cerrado, Atlantic Forest |
| Species [References] | abestado | antillensis | icelosmarias | marias | paramoensis | |
|---|---|---|---|---|---|---|
| sp. nov. | [12,45] | [48] | [12,45,49] | [47] | ||
| Type Locality: | Chapada Diamantina | Camp Perrin, | Brick Kiln, | Las Marias, | Páramo de Mucubají, | |
| Characteristics | Bahia, Brazil | Haiti | Saint Kitts and Nevis | Puerto Rico | Mérida, Venezuela | |
| Body bluish/violetish | Ant., anterior head, lateral Th., dorsal Abd. VI and part of legs | Ant., anterior head, part of legs, lateral Abd. I–IV and posterior Abd. III–IV | Ant., anterior head, lateral trunk, legs or entirely pigmented | Entire body and appendages, except furca | ||
| Pigmentation on: | Unpigmented ** | |||||
| Interocular chaetae | 5 | ? | 3–4 | ? | ? | |
| Dorsal head mac | Pp5 | − | − | −? | − | + |
| Basomedian and | R (smaller) | C | − | S | S | S/C |
| basolateral labial fields | L1 | S | ? | S | ? | C |
| L2 | S | ? | S | ? | C | |
| extra chaetae * | 0–1 S, 2–3 C | − | 3–4 S, 1C | 1–3 S, 1–2 C | 0–1 S, 6–7 C | |
| scales | − | + | ? | − | − | |
| Cephalic groove b.c. | 3 | ? | 3 | ? | 2 | |
| Th. II mac | m4 group | 3 | 2 | 3 | 3 | 2 |
| p row | 4 | 5 | 4 | 4 | 5 | |
| Th. III central mac | a, m, p rows | 3,0,3 | 4,1,3 | 3,0,3 | 3,0,3 | 2,1,3 |
| Abd. IV central mac | B5 | + | − | + | + | + |
| B6 | +/− | + | + | +/− | − | |
| Trochanteral organ chaetae | 22–27 | ? | ±23 | ? | 35–52 | |
| Tibiotarsus III inner smooth chaetae | 5 | − | + | + | 6 | |
| Tenent hair | acuminate/capitate | acuminate/capitate | capitate | acuminate/capitate | acuminate | |
| Unguis teeth | ratio | b.t.=m.t. > a.t. | b.t. = m.t. | b.t. > m.t.. | b.t. > m.t.. | b.t. > m.t. > a.t. |
| a.t. | + | − | – | − | +/− | |
| Unguiculus lamellae | ai | wide | normal | normal | normal | normal |
| pe | 1 tooth | 1 tooth | smooth | 1 tooth | 1 tooth | |
| Manubrium dorsal smooth mac | 3 + 3 | − | 4 + 4 | ? | 4 + 4 | |
| Dental spines | number | 16–20 | ? | 20 | 16–42 | ±16 |
| distribution | 2–3 rows grouped | variable | 2 rows grouped | 2–5 rows | 2 rows grouped | |
| Species [References] | abestado sp. nov. | albicephalus [21] | amazonicus [17] | bicolor [13] | cuprum [20] | heloisae [16,18] | magnus [21] | marimutti [21] | melinus [20] | pikachu [21] | termitophilus [13] | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Type Locality | Chapada Diamantina, Bahia | Serra dos Órgãos, Rio de Janeiro | Reserva Ducke, Amazonas | Blumenau, Santa Catarina | Serra da Gandarela, Minas Gerais | Floresta da Tijuca, Rio de Janeiro | Serra do Japi, São Paulo | Serra dos Órgãos, Rio de Janeiro | Serra da Gandarela, Minas Gerais | Serra do Japi, São Paulo | Minas Gerais | |
| Characteristics | Species Group | marias | gracilis | marias | ? | gracilis | gracilis | gracilis | gracilis | gracilis | marias | ? |
| Body bluish/violetish pigmentation on: | Ant. | + | + | + | Ant. II–IV | partially | + | partially | Ant. II–IV | + | partially | mostly |
| Head (– eyes) | anteriorly | anteriorly | weakly | − | + | + | partially | − | anteriorly | − | anteriorly | |
| Th. (dorsally) | − | + | − | − | + | − | + | Th. III | − | − | partially | |
| Abd. (dorsally) | Abd. VI | + | − | − | + | − | partially | + | − | − | partially | |
| Legs | partially | partially | weakly | mostly | proximally | −/+ | mostly | tibiotarsi | + | tibiotarsi | –? | |
| Ant. IIb smooth mac | 3 | −? | ? | ? | −? | 6? | 1? | 4? | −? | −? | ? | |
| Interocular chaetae | 5 | 3 | 3 | ? | 3 | 3 | 3 | 4 | 4 | 3 | ? | |
| Dorsal head mac | An row | 11–12 | 14–15 | ±11 | ? | 14 | 12 | 13 | 15 | 13 | 17 | ? |
| S6i | + | + | + | ? | + | + | + | + | − | + | ? | |
| posterior | 1 | 5 | 1 * | ? | 2 | 2 | 4 | 4 | 5 | 1 | ? | |
| Maxillary outer lobe b.c. | acuminate | blunt | acuminate | ? | acuminate | acuminate | acuminate | acuminate | acuminate | blunt | ? | |
| Labial basomedian field | M1 | C | C | C | ? | S | C | C | C | C | C | ? |
| R (smaller) | C | C | S | ? | S | S | S | C | S | C | ? | |
| extra chaetae | 0–1 S, 2–3 C | − | 2 S, 1 C | ? | − | 1 S | 1 S | 1 C *** | 1 S, 1 C | 1 C | ? | |
| scales | − | − | − | ? | − | + | − | − | − | − | ? | |
| Th. II mac | p row | 4 | 6 | 4 | ? | 6 | 7 | 7 | 6 | 6 | 5–6 | ? |
| Th. III central mac | a,m,p rows | 3,0,3 | 5,1,3 | 3,0,3 | ? | 5,1,3 | 5,1,3 | 5,1,3 | 5,1,3 | 5,1,3 | 5,1,3 | ? |
| Abd. I mac | 3 | 5 | 2 | ? | 5 | 5 | 5 | 5 | 5 | 5 | ? | |
| Abd. II mac | central | 2 | 2 | 1 | ? | 2 | 2 | 2 | 2 | 2 | 2 | ? |
| Abd. III mac | lateral | 1 | 1 | 2 | ? | 2 | 2 | 2 | 2 | 2 | 2 | ? |
| Abd. IV central mac | 4–5 | 5 | 3 | ? | 5 | 5 | 5 | 5 | 5 | 5 | ? | |
| Trochanteral organ chaetae | 22–27 | ±40 | ±30 | ? | ±70 | ±100 | ±100 | ±60 | ±65 | ±80 | ? | |
| Tenent hair | acuminate /capitate | acuminate | capitate | capitate | acuminate | acuminate/capitate | acuminate | acuminate | capitate | capitate | acuminate | |
| Unguis teeth | ratio | b.t. = m.t.>a.t. | b.t. = m.t. | b.t. = m.t. | b.t. = m.t. | b.t. = m.t. | b.t. = m.t.>a.t. | b.t. = m.t.>a.t. | b.t. = m.t. > a.t. | b.t. = m.t. | b.t. < m.t. | b.t. = m.t. |
| a.t. | + | − | − | − | − | + | + | + | − | − | − | |
| Unguiculus lamellae | ai | wide | normal | normal | normal | normal | normal | normal | narrow | normal | narrow | normal |
| pe | 1 tooth | 1 tooth | S | 1 tooth | 1 tooth | 1 tooth | 1 tooth | 1 tooth | 1 tooth ** | 1 tooth | S | |
| Dental spines | number | 16–20 | ±50 | ±19 | ±30? | ±50 | ±60 | ±60 | ±110 | ±55 | ±50 | 20–30 |
| distribution | 2–3 rows grouped | 5–6 rows grouped | 2 rows grouped | 4 rows grouped | grouped | 7 rows | 6–7 rows | variable | mostly grouped | grouped | 3 rows grouped | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellini, B.C.; Cipola, N.G.; Siqueira, O.J.R. A Survey of the Brazilian Dicranocentrus Schött (Collembola, Orchesellidae, Heteromurini) with the Description of a New Species and Notes on the Genus. Insects 2020, 11, 709. https://doi.org/10.3390/insects11100709
Bellini BC, Cipola NG, Siqueira OJR. A Survey of the Brazilian Dicranocentrus Schött (Collembola, Orchesellidae, Heteromurini) with the Description of a New Species and Notes on the Genus. Insects. 2020; 11(10):709. https://doi.org/10.3390/insects11100709
Chicago/Turabian StyleBellini, Bruno C., Nikolas G. Cipola, and Orquianne J. R. Siqueira. 2020. "A Survey of the Brazilian Dicranocentrus Schött (Collembola, Orchesellidae, Heteromurini) with the Description of a New Species and Notes on the Genus" Insects 11, no. 10: 709. https://doi.org/10.3390/insects11100709
APA StyleBellini, B. C., Cipola, N. G., & Siqueira, O. J. R. (2020). A Survey of the Brazilian Dicranocentrus Schött (Collembola, Orchesellidae, Heteromurini) with the Description of a New Species and Notes on the Genus. Insects, 11(10), 709. https://doi.org/10.3390/insects11100709