Chemosensory-Related Gene Family Members of the Horn Fly, Haematobia irritans irritans (Diptera: Muscidae), Identified by Transcriptome Analysis

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Horn Fly Tissue Collection and Total RNA Isolation
2.2. Library Preparation, Sequencing, and Pre-Processing
2.3. De Novo Transcriptome Assembly, Annotation, and Functional Categorization
2.4. Differential Gene Expression and Gene Ontology Enrichment Analyses
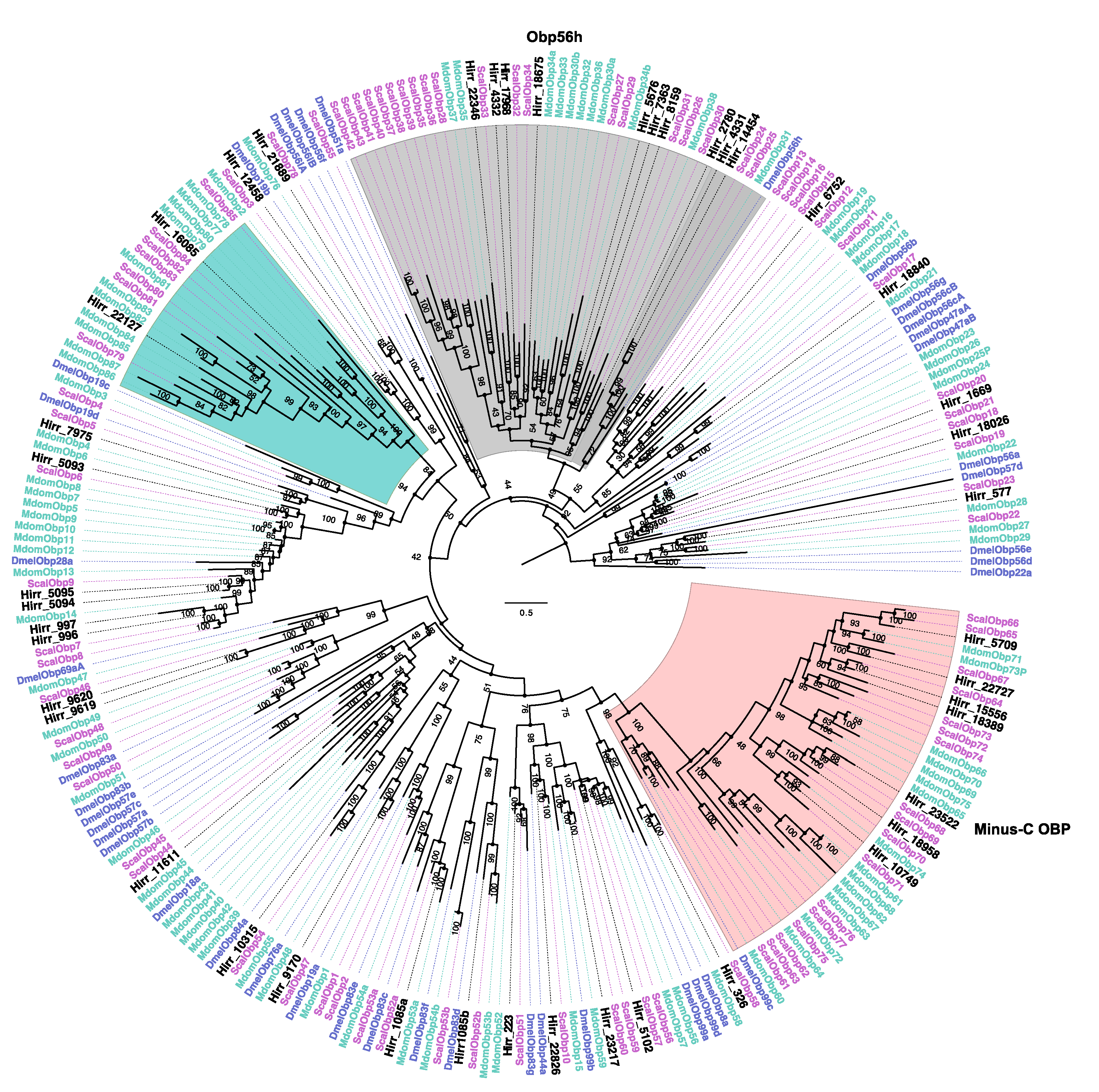
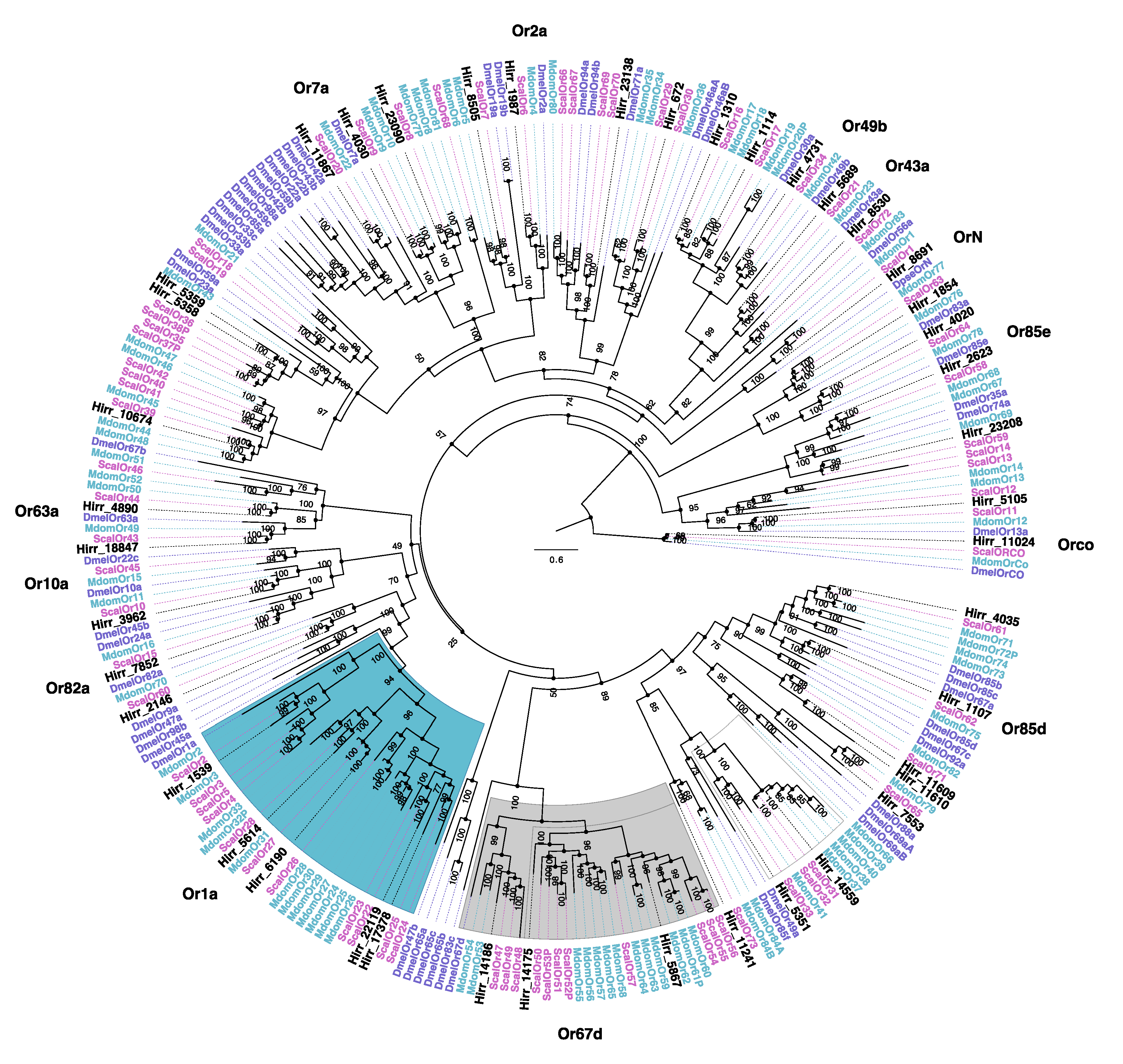
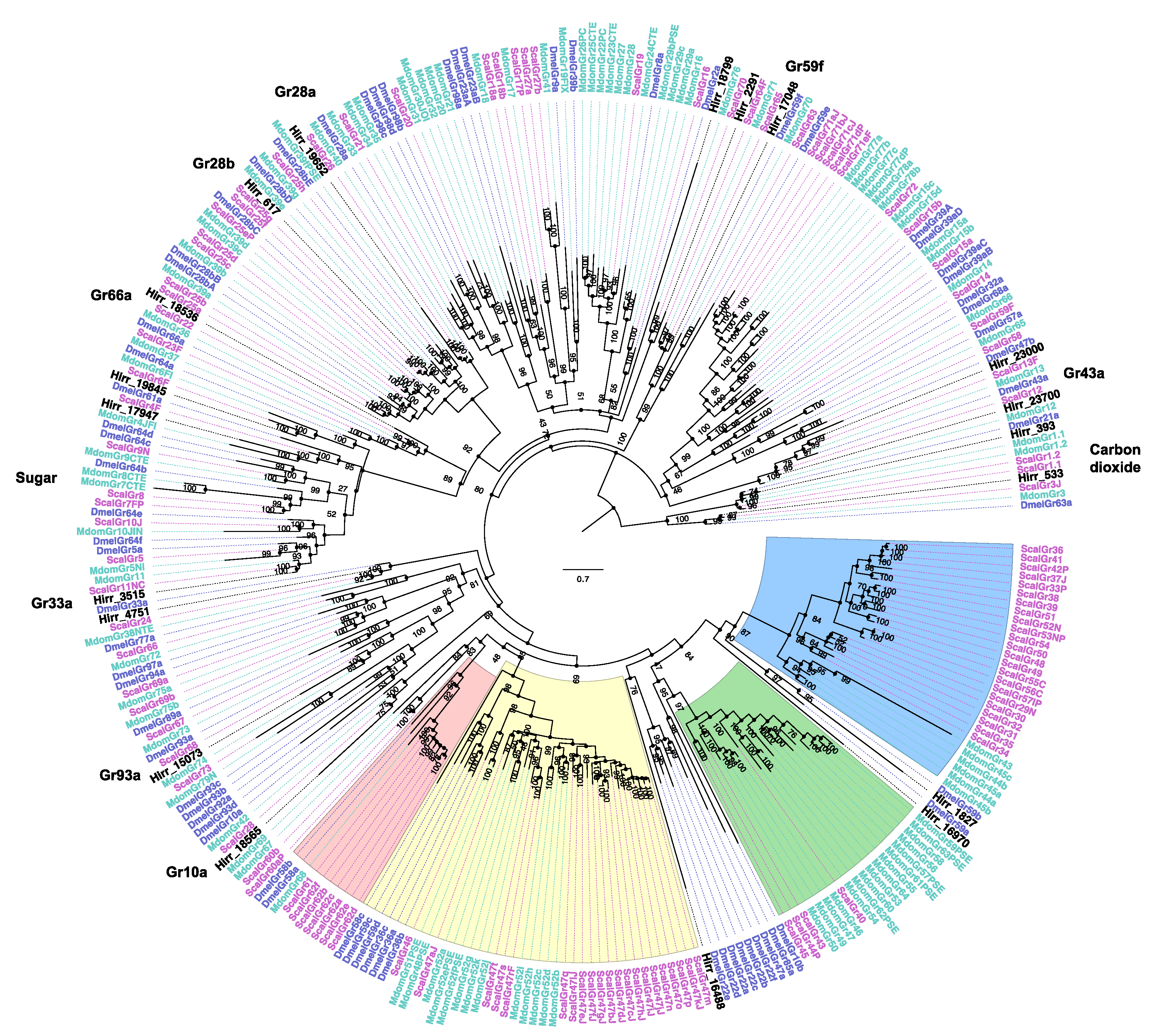
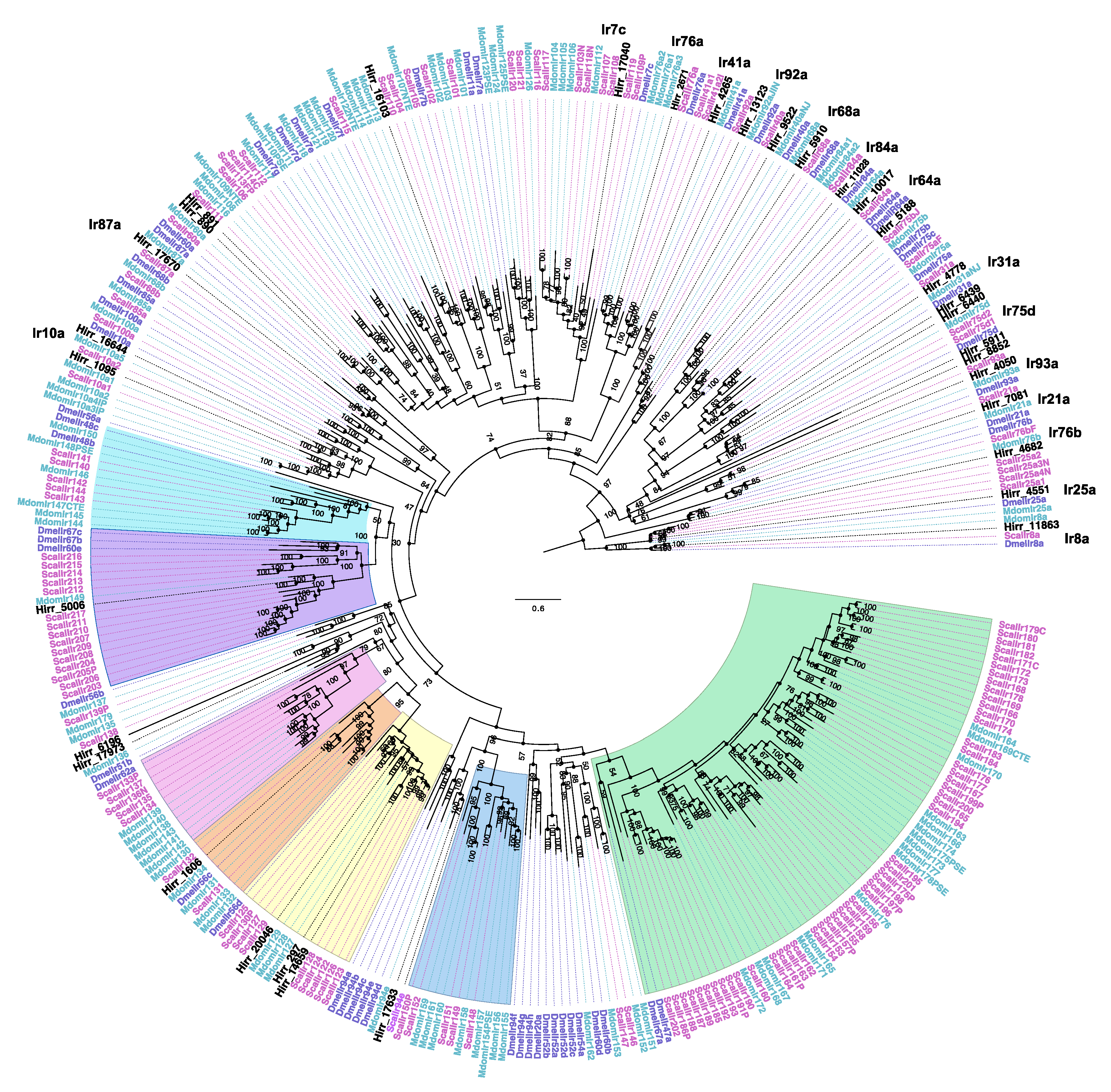
2.5. Phylogenetic Analysis
3. Results
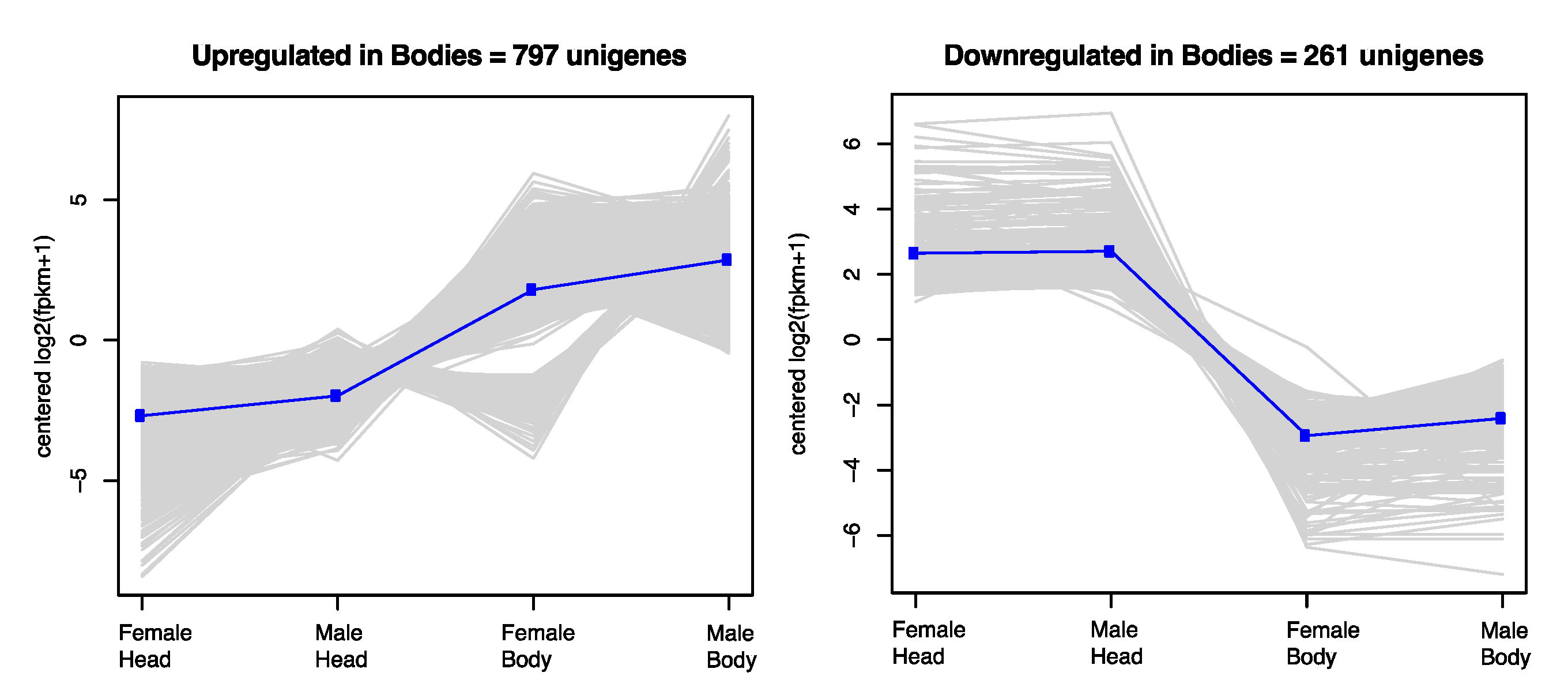
3.1. Transcriptome Sequencing, Assembly, and Differential Expression Analysis
3.2. Enriched Gene Ontology Categories of Differentially Expressed and Tissue-Specific Trancripts
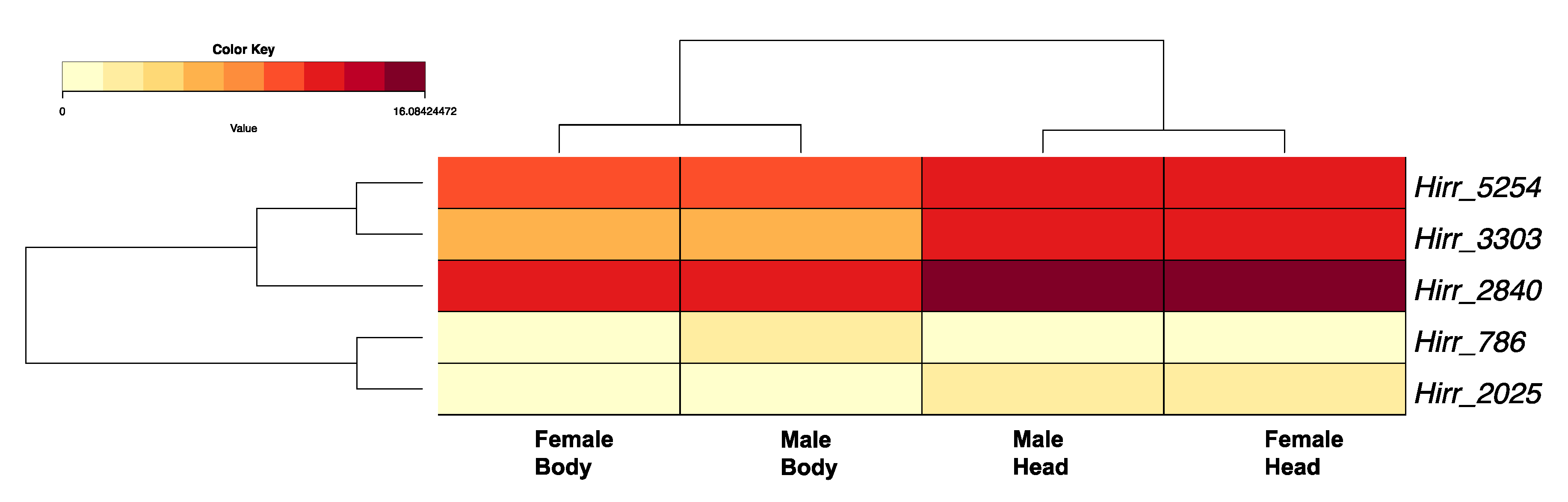
3.3. Candidate Horn Fly Chemosensory Gene Family Members Identified from Transcriptome Analysis of Pooled Head Appendages and Headless Bodies
3.3.1. Horn Fly Odorant Binding Proteins (OBP) and Chemosensory Proteins (CSP)
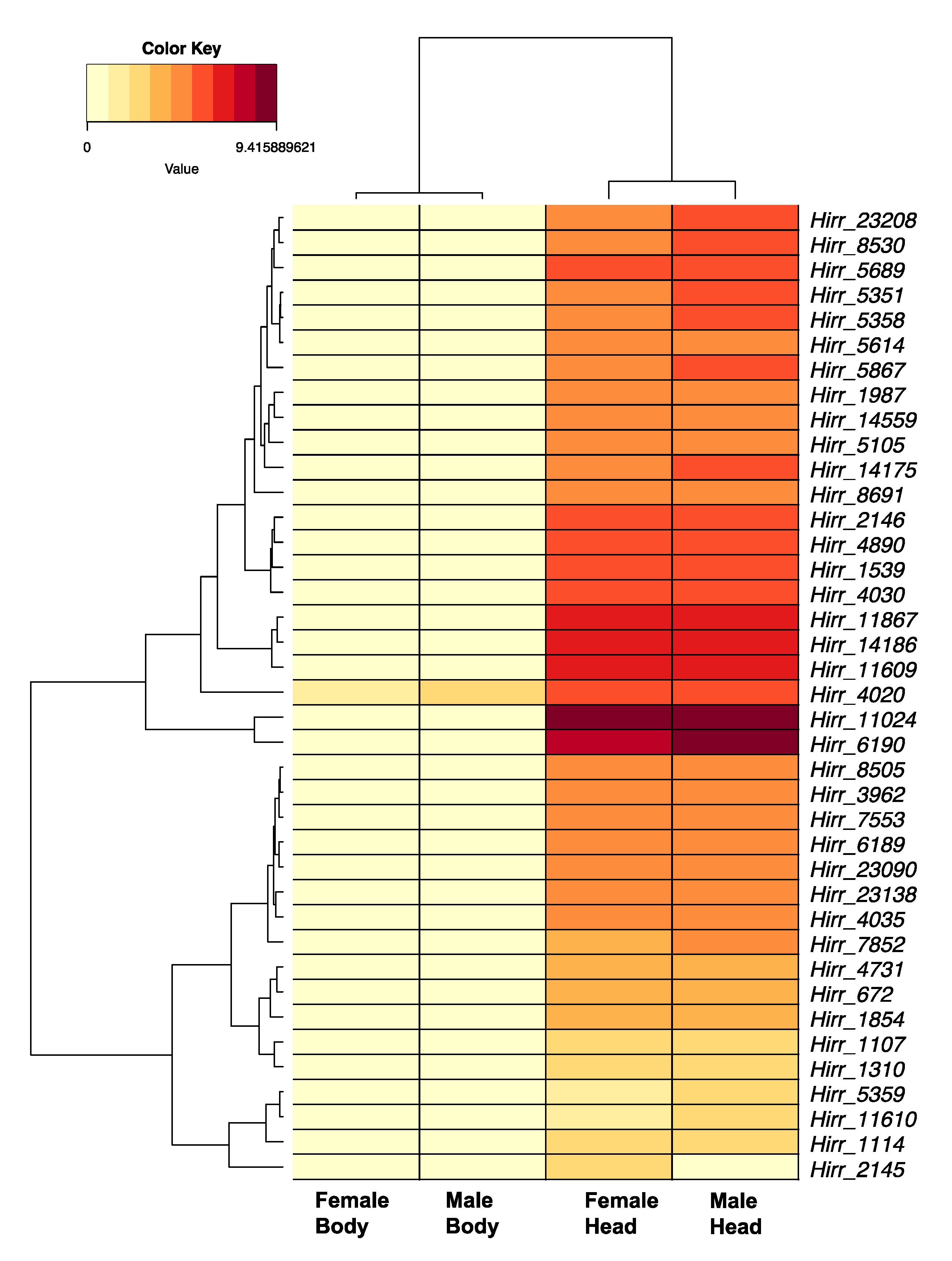
3.3.2. Horn Fly Odorant Receptors (OR)
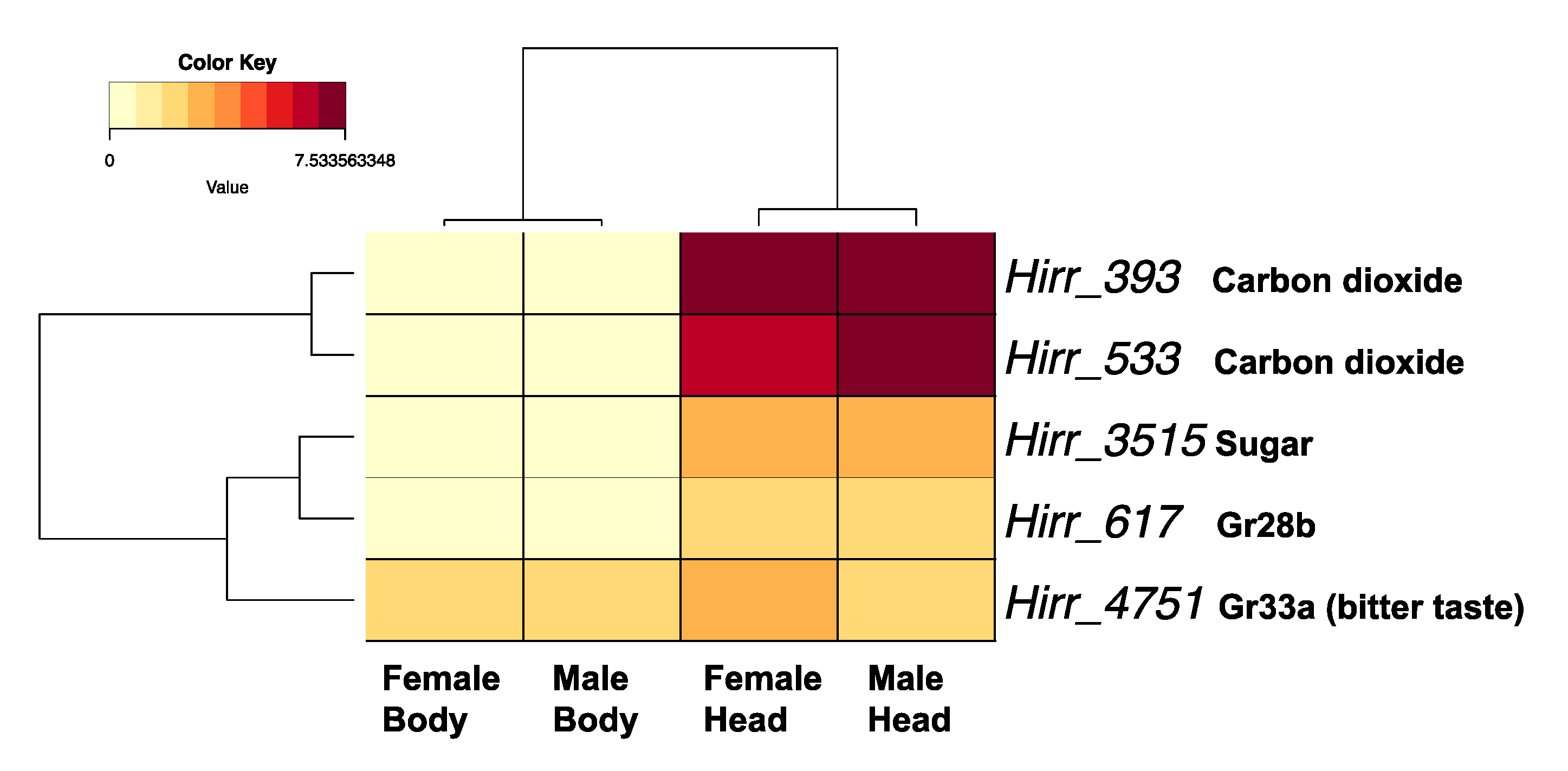
3.3.3. Horn Fly Gustatory Receptors (GR)
3.3.4. Horn Fly Ionotropic Receptors (IR)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swiger, S.L.; Tomberlein, J.K. Protecting Cattle from Horn Flies; Texas A&M AgriLife Extension: La Grange, TX, USA, 2011; p. E-208. [Google Scholar]
- Oyarzun, M.P.; Quiroz, A.; Birkett, M.A. Insecticide resistance in the horn fly: Alternative control strategies. Med. Vet. Entomol. 2008, 22, 188–202. [Google Scholar] [CrossRef] [PubMed]
- Domingues, L.N.; Guerrero, F.D.; Foil, L.D. Impacts of long-term insecticide treatment regimes on skdr and kdr pyrethroid resistance alleles in horn fly field populations. Parasitol. Res. 2019, 118, 2485–2497. [Google Scholar] [CrossRef] [PubMed]
- Steelman, C.D.; McNew, R.W.; Simpson, R.B.; Rorie, R.W.; Phillips, J.M.; Rosenkrans, C.F., Jr. Evaluation of alternative tactics for management of insecticide-resistant horn flies (Diptera: Muscidae). J. Econ. Entomol. 2003, 96, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Breijo, M.; Rocha, S.; Ures, X.; Pastro, L.; Alonzo, P.; Fernandez, C.; Meikle, A. Evaluation of Hematobin as a Vaccine Candidate to Control Haematobia irritans (Diptera: Muscidae) Loads in Cattle. J. Econ. Entomol. 2017, 110, 1390–1393. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M.; Debat, H.J.; Boiani, M.; Ures, X.; Rocha, S.; Breijo, M. An insight into the sialome, mialome and virome of the horn fly, Haematobia irritans. BMC Genom. 2019, 20, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denning, S.S.; Washburn, S.P.; Watson, D.W. Development of a novel walk-through fly trap for the control of horn flies and other pests on pastured dairy cows. J. Dairy Sci. 2014, 97, 4624–4631. [Google Scholar] [CrossRef]
- Zhu, J.J.; Cermak, S.C.; Kenar, J.A.; Brewer, G.; Haynes, K.F.; Boxler, D.; Baker, P.D.; Wang, D.; Wang, C.; Li, A.Y.; et al. Better than DEET: Repellent compounds derived from coconut oil. Sci. Rep. 2018, 8, 14053. [Google Scholar] [CrossRef]
- Lachance, S.; Grange, G. Repellent effectiveness of seven plant essential oils, sunflower oil and natural insecticides against horn flies on pastured dairy cows and heifers. Med. Vet. Entomol. 2014, 28, 193–200. [Google Scholar] [CrossRef]
- Zhu, J.J.; Brewer, G.J.; Boxler, D.J.; Friesen, K.; Taylor, D.B. Comparisons of antifeedancy and spatial repellency of three natural product repellents against horn flies, Haematobia irritans (Diptera: Muscidae). Pest Manag. Sci. 2015, 71, 1553–1560. [Google Scholar] [CrossRef] [Green Version]
- Mullens, B.A.; Reifenrath, W.G.; Butler, S.M. Laboratory trials of fatty acids as repellents or antifeedants against houseflies, horn flies and stable flies (Diptera: Muscidae). Pest Manag. Sci. 2009, 65, 1360–1366. [Google Scholar] [CrossRef]
- Mullens, B.A.; Soto, D.; Gerry, A.C. Sex ratios and mating status of the horn fly on pastured cattle as a function of repellent-oil treatments. J. Med. Entomol. 2018, 55, 1325–1329. [Google Scholar] [CrossRef] [PubMed]
- Mullens, B.A.; Soto, D.; Gerry, A.C.; Fowler, F.E.; Diniz, A.N. Effects of fatty acid and geraniol repellent-oil mixtures applied to cattle on blood feeding and reproductive parameters in field populations of Haematobia irritans (Diptera: Muscidae). J. Med. Entomol. 2018, 55, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Mullens, B.A.; Watson, D.W.; Gerry, A.C.; Sandelin, B.A.; Soto, D.; Rawls, D.; Denning, S.; Guisewite, L.; Cammack, J. Field trials of fatty acids and geraniol applied to cattle for suppression of horn flies, Haematobia irritans (Diptera: Muscidae), with observations on fly defensive behaviors. Vet. Parasitol. 2017, 245, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.N.; Newcomb, R.D. Pest control compounds targeting insect chemoreceptors: Another silent spring? Front. Ecol. Evol. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Sparks, J.T.; Bohbot, J.D.; Dickens, J.C. Olfactory disruption: Toward controlling important insect vectors of disease. Prog. Mol. Biol. Transl. Sci. 2015, 130, 81–108. [Google Scholar] [CrossRef]
- Venthur, H.; Zhou, J.J. Odorant Receptors and Odorant-Binding Proteins as Insect Pest Control Targets: A Comparative Analysis. Front. Physiol. 2018, 9, 1163. [Google Scholar] [CrossRef]
- Clark, J.T.; Ray, A. Olfactory mechanisms for discovery of odorants to reduce insect-host contact. J. Chem. Ecol. 2016, 42, 919–930. [Google Scholar] [CrossRef]
- Kuramochi, K. Studies on the Reproductive-Biology of the Horn Fly, Hematobia-Irritans (L.) (Diptera, Muscidae): 3. Mating-Behavior of the Fly in the Field. Appl. Entomol. Zool. 1989, 24, 326–333. [Google Scholar] [CrossRef]
- Zorka, T.J.; Bay, D.E. The courtship behavior of the horn fly. Southwest Entomol. 1980, 5, 196–200. [Google Scholar]
- Bolton, H.T.; Butler, J.F.; Carlson, D.A. A mating stimulant pheromone of the horn fly, Haematobia irritans (L.): Demonstration of biological activity in separated cuticular components. J. Chem. Ecol. 1980, 6, 951–964. [Google Scholar] [CrossRef]
- Kunz, S.E.; Blume, R.R.; Hogan, B.F.; Matter, J.J. Biological and ecological investigations of horn flies in Central Texas: Influence of time of manure deposition on oviposition. J. Econ. Entomol. 1970, 63, 930–933. [Google Scholar] [CrossRef]
- Kuramochi, K. Ovipositional behavior of the horn fly (Diptera: Muscidae) in the field. J. Med. Entomol. 2000, 37, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Birkett, M.A.; Agelopoulos, N.; Jensen, K.M.; Jespersen, J.B.; Pickett, J.A.; Prijs, H.J.; Thomas, G.; Trapman, J.J.; Wadhams, L.J.; Woodcock, C.M. The role of volatile semiochemicals in mediating host location and selection by nuisance and disease-transmitting cattle flies. Med. Vet. Entomol. 2004, 18, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Bay, D.E.; Meola, S.M.; White, S.L. Scanning electron microscopy of the ovipositor of the horn fly. Southwest Entomol. 1996, 21, 337–339. [Google Scholar]
- Fernandes, F.D.; Pimenta, P.F.P. Typology and distribution of antennal sensory organs of adults Haematobia irritans of the Midwest region of Brazil. Pesqui. Vet. Bras. 2018, 38, 183–188. [Google Scholar] [CrossRef]
- Fernandes, F.F.; Bahia, A.C.; Secundino, N.F.C.; Pimenta, P.F.P. Ultrastructural analysis of mouthparts of adult horn fly (Diptera: Muscidae) from the Brazilian Midwest region. J. Med. Entomol. 2020, 57, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- White, S.L.; Bay, D.E. Antennal olfactory sensilla of the horn fly, Haematobia irritans (L.) (Diptera: Muscidae). J. Kans. Entomol. Soc. 1980, 53, 641–652. [Google Scholar]
- Olafson, P.U. Molecular characterization and immunolocalization of the olfactory co-receptor Orco from two blood-feeding muscid flies, the stable fly (Stomoxys calcitrans, L.) and the horn fly (Haematobia irritans irritans, L.). Insect Mol. Biol. 2013, 22, 131–142. [Google Scholar] [CrossRef]
- Domingues, L.N.; Guerrero, F.D.; Cameron, C.; Farmer, A.; Bendele, K.G.; Foil, L.D. The assembled transcriptome of the adult horn fly, Haematobia irritans. Data Brief 2018, 19, 1933–1940. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Bairoch, A.; Apweiler, R. The SWISS-PROT protein sequence database and its supplement TrEMBL in 2000. Nucleic Acids Res. 2000, 28, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.D.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. PANTHER: A library of protein families and subfamilies indexed by function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olafson, P.U.; Aksoy, S.; Attardo, G.M.; Buckmeier, G.; Chen, X.; Coates, C.J.; Davis, M.; Dykema, J.; Emrich, S.J.; Friedrich, M.; et al. Functional genomics of the stable fly, Stomoxys calcitrans, reveals mechanisms underlying reproduction, host interactions, and novel targets for pest control. bioRxiv 2019. [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutierrez, S.; Silla-Martinez, J.M.; Gabaldon, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Rimal, S.; Lee, Y. The multidimensional ionotropic receptors of Drosophila melanogaster. Insect Mol. Biol. 2018, 27, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.G.; Warren, W.C.; Beukeboom, L.W.; Bopp, D.; Clark, A.G.; Giers, S.D.; Hediger, M.; Jones, A.K.; Kasai, S.; Leichter, C.A.; et al. Genome of the house fly, Musca domestica L., a global vector of diseases with adaptations to a septic environment. Genome Biol. 2014, 15. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Ma, H.; Xie, H.; Xuan, N.; Guo, X.; Fan, Z.; Rajashekar, B.; Arnaud, P.; Offmann, B.; Picimbon, J.F. Biotype Characterization, Developmental Profiling, Insecticide Response and Binding Property of Bemisia tabaci Chemosensory Proteins: Role of CSP in Insect Defense. PLoS ONE 2016, 11, e0154706. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, P.; Zhu, J.; Knoll, W. From radioactive ligands to biosensors: Binding methods with olfactory proteins. Appl. Microbiol. Biotechnol. 2018, 102, 8213–8227. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Xuan, N.; Rajashekar, B.; Arnaud, P.; Offmann, B.; Picimbon, J.F. Comprehensive history of CSP genes: Evolution, phylogenetic distribution and functions. Genes 2020, 11, 413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, F.G.; Rozas, J. Comparative genomics of the odorant-binding and chemosensory protein gene families across the Arthropoda: Origin and evolutionary history of the chemosensory system. Genome Biol. Evol. 2011, 3, 476–490. [Google Scholar] [CrossRef] [PubMed]
- Campanacci, V.; Spinelli, S.; Lartigue, A.; Lewandowski, C.; Brown, K.; Tegoni, M.; Cambillau, C. Recombinant chemosensory protein (CSP2) from the moth Mamestra brassicae: Crystallization and preliminary crystallographic study. Acta Crystallogr. Sect. D Biol. Crystallogr. 2001, 57, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Xu, X.; Xu, W.; Ishida, Y.; Leal, W.S.; Ames, J.B.; Clardy, J. Crystal and solution structures of an odorant-binding protein from the southern house mosquito complexed with an oviposition pheromone. Proc. Natl. Acad. Sci. USA 2010, 107, 19102–19107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olafson, P.U.; Lohmeyer, K.H.; Dowd, S.E. Analysis of expressed sequence tags from a significant livestock pest, the stable fly (Stomoxys calcitrans), identifies transcripts with a putative role in chemosensation and sex determination. Arch. Insect Biochem. Physiol. 2010, 74, 179–204. [Google Scholar] [CrossRef]
- Wanner, K.W.; Willis, L.G.; Theilmann, D.A.; Isman, M.B.; Feng, Q.; Plettner, E. Analysis of the insect OS-D-like gene family. J. Chem. Ecol. 2004, 30, 889–911. [Google Scholar] [CrossRef]
- Pitts, R.J.; Rinker, D.C.; Jones, P.L.; Rokas, A.; Zwiebel, L.J. Transcriptome profiling of chemosensory appendages in the malaria vector Anopheles gambiae reveals tissue- and sex-specific signatures of odor coding. BMC Genom. 2011, 12, 271. [Google Scholar] [CrossRef] [Green Version]
- Younus, F.; Chertemps, T.; Pearce, S.L.; Pandey, G.; Bozzolan, F.; Coppin, C.W.; Russell, R.J.; Maibeche-Coisne, M.; Oakeshott, J.G. Identification of candidate odorant degrading gene/enzyme systems in the antennal transcriptome of Drosophila melanogaster. Insect Biochem. Mol. Biol. 2014, 53, 30–43. [Google Scholar] [CrossRef] [Green Version]
- Shorter, J.R.; Dembeck, L.M.; Everett, L.J.; Morozova, T.V.; Arya, G.H.; Turlapati, L.; St. Armour, G.E.; Schal, C.; Mackay, T.F.C.; Anholt, R.R.H. Obp56h modulates mating behavior in Drosophila melanogaster. G3 Genes Genom. Genet. 2016, 6, 3335–3342. [Google Scholar] [CrossRef] [Green Version]
- Hekmat-Scafe, D.S.; Scafe, C.R.; McKinney, A.J.; Tanouye, M.A. Genome-wide analysis of the odorant-binding protein gene family in Drosophila melanogaster. Genome Res. 2002, 12, 1357–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinelli, S.; Lagarde, A.; Iovinella, I.; Legrand, P.; Tegoni, M.; Pelosi, P.; Cambillau, C. Crystal structure of Apis mellifera OBP14, a C-minus odorant-binding protein, and its complexes with odorant molecules. Insect Biochem. Mol. Biol. 2012, 42, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.D.; Dahanukar, A. Recent advances in the genetic basis of taste detection in Drosophila. Cell. Mol. Life Sci. 2020, 77, 1087–1101. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, J.; Pregitzer, P.; Breer, H.; Krieger, J. Access to the odor world: Olfactory receptors and their role for signal transduction in insects. Cell. Mol. Life Sci. 2018, 75, 485–508. [Google Scholar] [CrossRef]
- Robertson, H.M. The insect chemoreceptor superfamily in Drosophila pseudoobscura: Molecular evolution of ecologically-relevant genes over 25 million years. J. Insect Sci. 2009, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- van Naters, W.G.; Carlson, J.R. Receptors and neurons for fly odors in Drosophila. Curr. Biol. 2007, 17, 606–612. [Google Scholar] [CrossRef] [Green Version]
- Kurtovic, A.; Widmer, A.; Dickson, B.J. A single class of olfactory neurons mediates behavioural responses to a Drosophila sex pheromone. Nature 2007, 446, 542–546. [Google Scholar] [CrossRef]
- Masuda-Nakagawa, L.M.; Gendre, N.; O’Kane, C.J.; Stocker, R.F. Localized olfactory representation in mushroom bodies of Drosophila larvae. Proc. Natl. Acad. Sci. USA 2009, 106, 10314–10319. [Google Scholar] [CrossRef] [Green Version]
- Fishilevich, E.; Domingos, A.I.; Asahina, K.; Naef, F.; Vosshall, L.B.; Louis, M. Chemotaxis behavior mediated by single larval olfactory neurons in Drosophila. Curr. Biol. 2005, 15, 2086–2096. [Google Scholar] [CrossRef] [Green Version]
- Couto, A.; Alenius, M.; Dickson, B.J. Molecular, anatomical, and functional organization of the Drosophila olfactory system. Curr. Biol. 2005, 15, 1535–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, A.L.; van Naters, W.G.; Lessing, D.; Warr, C.G.; Carlson, J.R. Coexpression of two functional odor receptors in one neuron. Neuron 2005, 45, 661–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, W.D.; Cayirlioglu, P.; Kadow, I.G.; Vosshall, L.B. Two chemosensory receptors together mediate carbon dioxide detection in Drosophila. Nature 2007, 445, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Yavuz, A.; Slone, J.; Jagge, C.; Song, X.; Amrein, H. Drosophila sugar receptors in sweet taste perception, olfaction, and internal nutrient sensing. Curr. Biol. 2015, 25, 621–627. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Moon, S.J.; Wang, X.; Ren, Q.; Montell, C. Gr64f is required in combination with other gustatory receptors for sugar detection in Drosophila. Curr. Biol. 2008, 18, 1797–1801. [Google Scholar] [CrossRef] [Green Version]
- Slone, J.; Daniels, J.; Amrein, H. Sugar receptors in Drosophila. Curr. Biol. 2007, 17, 1809–1816. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Tanaka, K.; Touhara, K. Sugar-regulated cation channel formed by an insect gustatory receptor. Proc. Natl. Acad. Sci. USA 2011, 108, 11680–11685. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Slone, J.; Song, X.; Amrein, H. A fructose receptor functions as a nutrient sensor in the Drosophila brain. Cell 2012, 151, 1113–1125. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.J.; Kottgen, M.; Jiao, Y.; Xu, H.; Montell, C. A taste receptor required for the caffeine response in vivo. Curr. Biol. 2006, 16, 1812–1817. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.J.; Lee, Y.; Jiao, Y.; Montell, C. A Drosophila gustatory receptor essential for aversive taste and inhibiting male-to-male courtship. Curr. Biol. 2009, 19, 1623–1627. [Google Scholar] [CrossRef] [Green Version]
- Thorne, N.; Amrein, H. Atypical expression of Drosophila gustatory receptor genes in sensory and central neurons. J. Comp. Neurol. 2008, 506, 548–568. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Bronk, P.; Chang, E.C.; Lowell, A.M.; Flam, J.O.; Panzano, V.C.; Theobald, D.L.; Griffith, L.C.; Garrity, P.A. A gustatory receptor paralogue controls rapid warmth avoidance in Drosophila. Nature 2013, 500, 580–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, D.; Thorne, N.; Miyamoto, C.; Jagge, C.; Amrein, H. The taste of ribonucleosides: Novel macronutrients essential for larval growth are sensed by Drosophila gustatory receptor proteins. PLoS Biol. 2018, 16, e2005570. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Diaz, C.; Martin, F.; Garcia-Fernandez, J.M.; Alcorta, E. The Two Main Olfactory Receptor Families in Drosophila, ORs and IRs: A Comparative Approach. Front. Cell. Neurosci. 2018, 12, 253. [Google Scholar] [CrossRef] [PubMed]
- Abuin, L.; Bargeton, B.; Ulbrich, M.H.; Isacoff, E.Y.; Kellenberger, S.; Benton, R. Functional architecture of olfactory ionotropic glutamate receptors. Neuron 2011, 69, 44–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knecht, Z.A.; Silbering, A.F.; Ni, L.; Klein, M.; Budelli, G.; Bell, R.; Abuin, L.; Ferrer, A.J.; Samuel, A.D.; Benton, R.; et al. Distinct combinations of variant ionotropic glutamate receptors mediate thermosensation and hygrosensation in Drosophila. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Klein, M.; Svec, K.V.; Budelli, G.; Chang, E.C.; Ferrer, A.J.; Benton, R.; Samuel, A.D.; Garrity, P.A. The Ionotropic Receptors IR21a and IR25a mediate cool sensing in Drosophila. Elife 2016, 5. [Google Scholar] [CrossRef]
- Koh, T.W.; He, Z.; Gorur-Shandilya, S.; Menuz, K.; Larter, N.K.; Stewart, S.; Carlson, J.R. The Drosophila IR20a clade of ionotropic receptors are candidate taste and pheromone receptors. Neuron 2014, 83, 850–865. [Google Scholar] [CrossRef] [Green Version]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
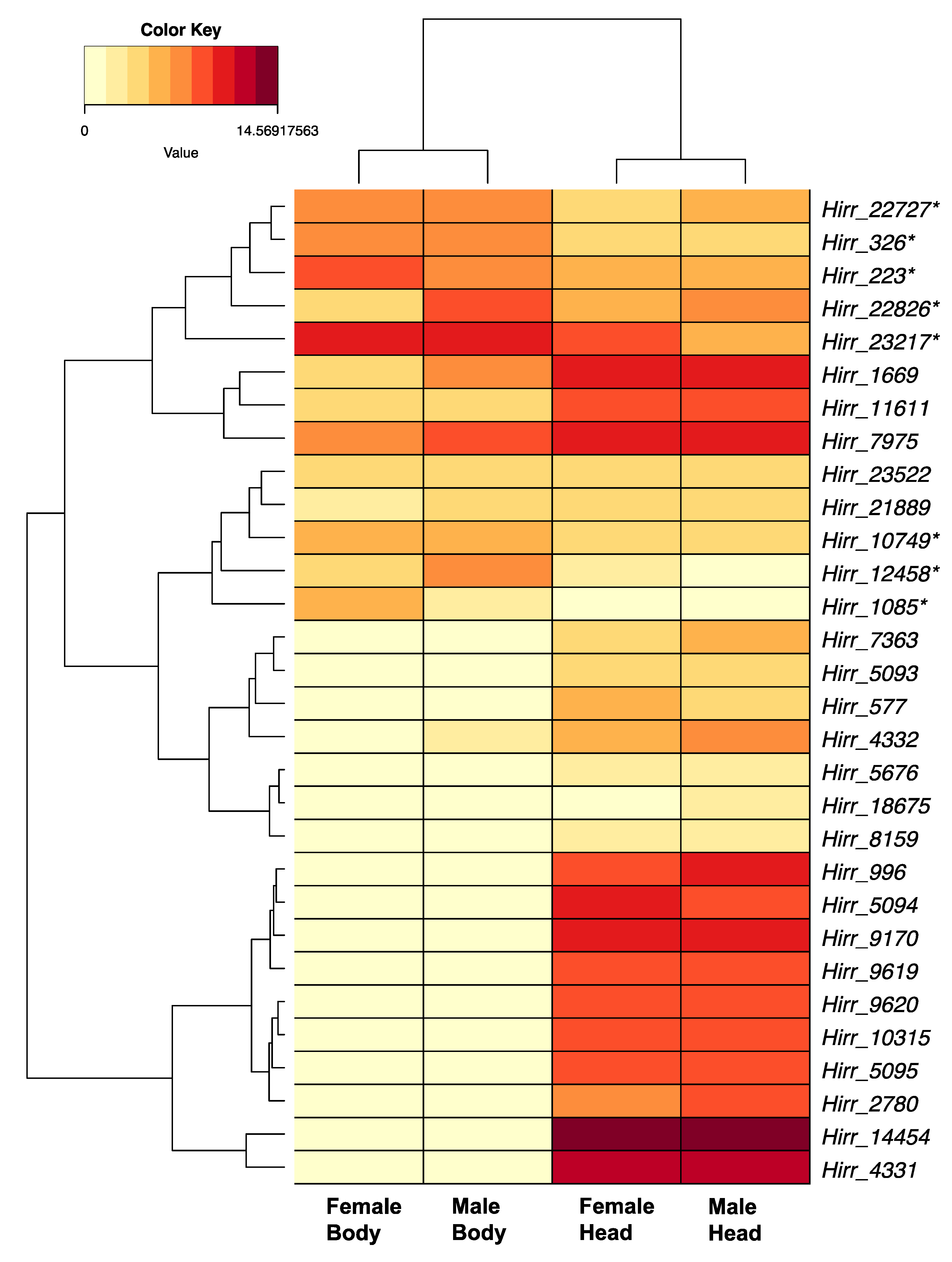{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession (Sequence Read Archive) | Sample Dataset Pool | Total Number Read Pairs | Total Number Bases |
|---|---|---|---|
| SRR12763132 | Female head appendage | 19,605,733 | 4,901,433,250 |
| SRR12763131 | Female headless bodies | 28,525,717 | 7,131,429,250 |
| SRR12763130 | Male head appendage | 20,632,837 | 5,158,209,250 |
| SRR12763129 | Male headless bodies | 51,464,662 | 12,866,165,500 |
| For de novo transcriptome | 120,228,949 | 30,057,237,250 |
| Sequence ID | Condition a | Log2 Fold Change b | ||
|---|---|---|---|---|
| A | B | A | B | |
| Hirr_10315 | fh > fb | mh > mb | 8.04 | 8.21 |
| Hirr_11611 | fh > fb | mh > mb | 3.95 | 3.64 |
| Hirr_14454 | fh > fb | mh > mb | 12.98 | 14.15 |
| Hirr_1669 | fh > fb | mh > mb | 6.99 | 3.15 |
| Hirr_18675 | mh > mb | 2.38 | ||
| Hirr_2780 | fh > fb | mh > mb | 7.67 | 7.86 |
| Hirr_4331 | fh > fb | mh > mb | 11.84 | 12.01 |
| Hirr_4332 | fh > fb | mh > mb | 5.43 | 3.99 |
| Hirr_5093 | fh > fb | mh > mb | 3.82 | 3.87 |
| Hirr_5094 | fh > fb | mh > mb | 9.55 | 9.09 |
| Hirr_5095 | fh > fb | mh > mb | 8.73 | 7.18 |
| Hirr_5676 | mh > mb | 2.6 | ||
| Hirr_577 | fh > fb | mh > mb | 5.92 | 3.8 |
| Hirr_7363 | fh > fb | mh > mb | 3.85 | 5.11 |
| Hirr_7975 | fh > fb | 2.69 | ||
| Hirr_8159 | fh > fb | 2.06 | ||
| Hirr_9170 | fh > fb | mh > mb | 8.87 | 9.14 |
| Hirr_9619 | fh > fb | mh > mb | 8.61 | 9.2 |
| Hirr_9620 | fh > fb | mh > mb | 8.35 | 8.58 |
| Hirr_996 | fh > fb | mh > mb | 9.19 | 9.6 |
| Hirr_10749 | fb > fh | mb > mh | 2.1 | 3.02 |
| Hirr_1085 | fb > fh | mb > mh | 5.06 | 2.61 |
| Hirr_12458 | fb > fh | mb > mh | 3.04 | 6.25 |
| Hirr_223 | fb > fh | mb > mh | 2.82 | 2.68 |
| Hirr_22727 | fb > fh | mb > mh | 3.09 | 3.6 |
| Hirr_22826 | mb > mh | 2.68 | ||
| Hirr_23217 | fb > fh | mb > mh | 2.19 | 5.08 |
| Hirr_23217 | fh > mh | 3.91 | ||
| Hirr_326 | fb > fh | mb > mh | 3.95 | 3.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olafson, P.U.; Saski, C.A. Chemosensory-Related Gene Family Members of the Horn Fly, Haematobia irritans irritans (Diptera: Muscidae), Identified by Transcriptome Analysis. Insects 2020, 11, 816. https://doi.org/10.3390/insects11110816
Olafson PU, Saski CA. Chemosensory-Related Gene Family Members of the Horn Fly, Haematobia irritans irritans (Diptera: Muscidae), Identified by Transcriptome Analysis. Insects. 2020; 11(11):816. https://doi.org/10.3390/insects11110816
Chicago/Turabian StyleOlafson, Pia Untalan, and Christopher A. Saski. 2020. "Chemosensory-Related Gene Family Members of the Horn Fly, Haematobia irritans irritans (Diptera: Muscidae), Identified by Transcriptome Analysis" Insects 11, no. 11: 816. https://doi.org/10.3390/insects11110816
APA StyleOlafson, P. U., & Saski, C. A. (2020). Chemosensory-Related Gene Family Members of the Horn Fly, Haematobia irritans irritans (Diptera: Muscidae), Identified by Transcriptome Analysis. Insects, 11(11), 816. https://doi.org/10.3390/insects11110816