The Invasion and Encapsulation of the Entomopathogenic Nematode, Steinernema abbasi, in Aedes albopictus (Diptera: Culicidae) Larvae


Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing
2.2. Nematode Propagation
2.3. Infection Assay of Ae. albopictus
2.4. Observations on Routes of Entry of S. abbasi Into Ae. albopictus
2.5. Observations on Encapsulation of Invading Nematodes in Ae. albopictus
3. Results
3.1. Virulence of S. abbasi and S. carpocapsae to Ae. albopictus Larvae
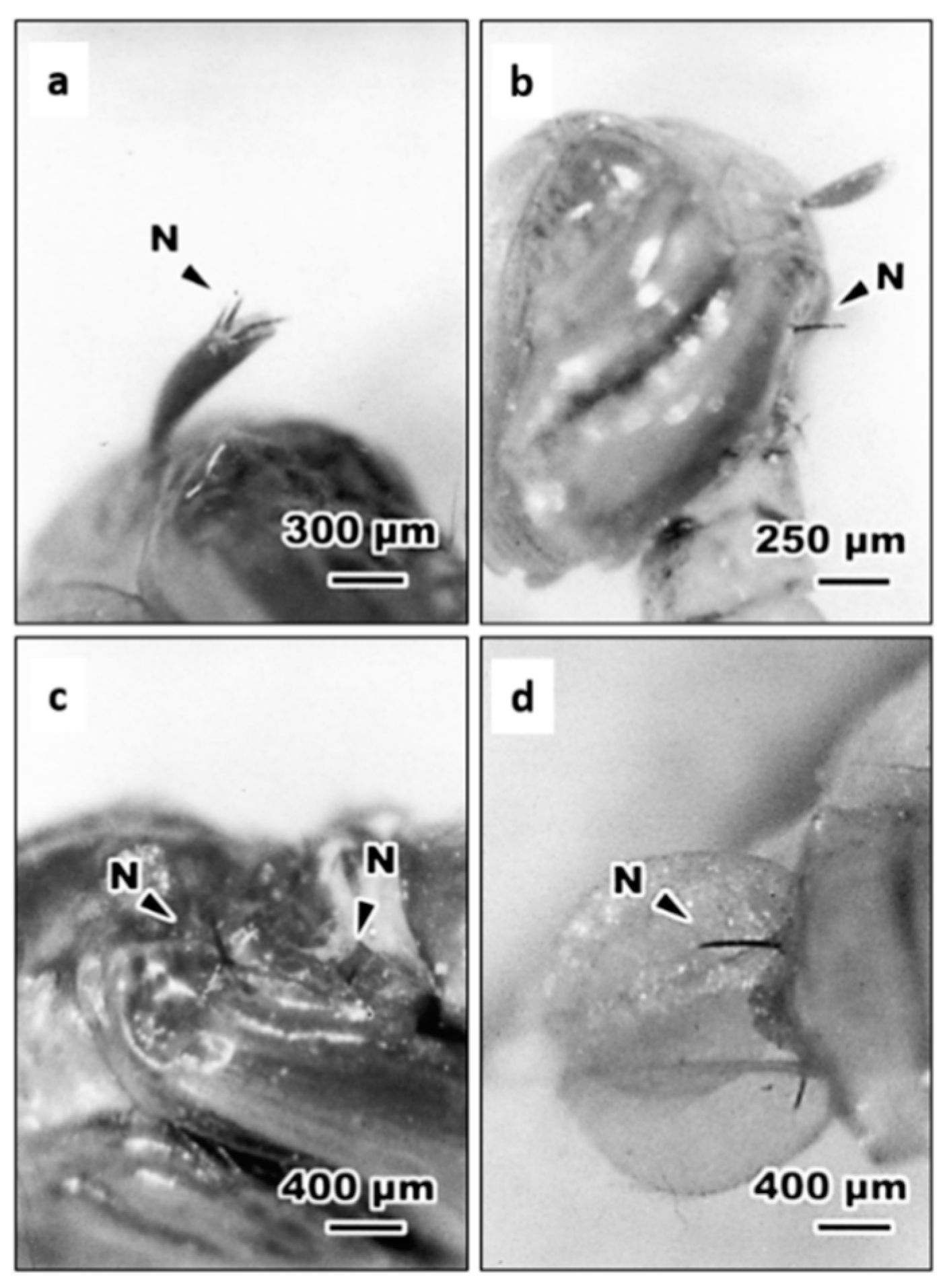
3.2. Routes of Entry of S. abbasi Into Ae. albopictus
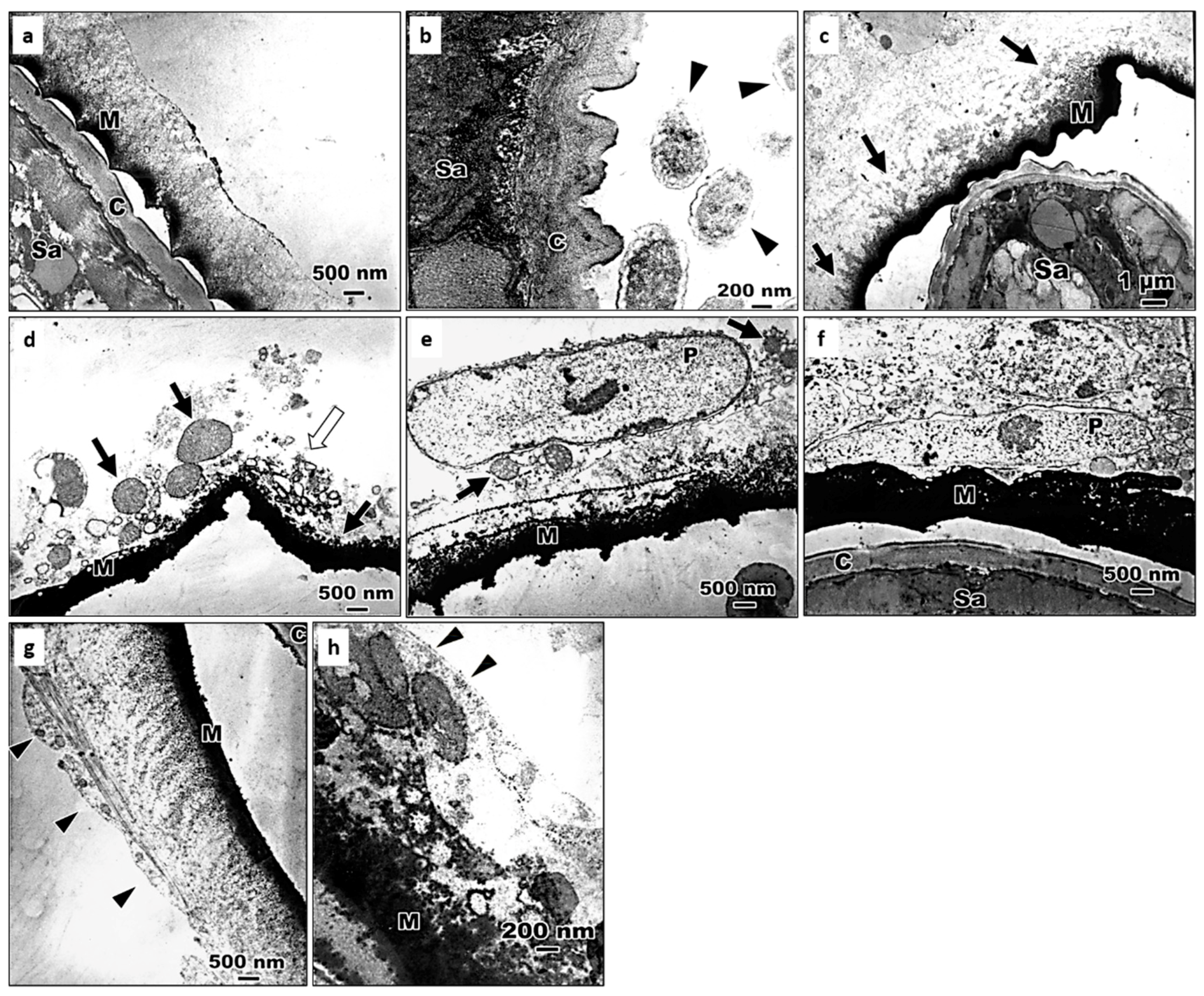
3.3. The Encapsulation of S. abbasi in the 4th Instar Larvae of Ae. albopictus
3.4. The Processes of Encapsulation of S. abbasi in Ae. albopictus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Leta, S.; Beyene, T.J.; De Clercq, E.M.; Amenu, K.; Kraemer, M.U.G.; Revie, C.W. Global risk mapping for major diseases transmitted by Aedes aegypti and Aedes albopictus. Int. J. Infect. Dis. 2018, 67, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonizzoni, M.; Gasperi, G.; Chen, X.; James, A.A. The invasive mosquito species Aedes albopictus: Current knowledge and future perspectives. Trends Parasitol. 2013, 29, 460–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, T.W.; Lorenz, L.H.; Weaver, S.C. Susceptibility of Aedes albopictus to infection with eastern equine encephalomyelitis virus. J. Am. Mosq. Control. Assoc. 1990, 6, 274–278. [Google Scholar]
- Mitchell, C.J.; Niebylski, M.L.; Smith, G.C.; Karabatsos, N.; Martin, D.; Mutebi, J.P.; Craig, G.B., Jr.; Mahler, M.J. Isolation of eastern equine encephalitis virus from Aedes albopictus in Florida. Science 1992, 257, 526–527. [Google Scholar] [CrossRef]
- Sardelis, M.R.; Turell, M.J.; O’Guinn, M.L.; Andre, R.G.; Roberts, D.R. Vector competence of three north American strains of Aedes albopictus for West Nile virus. J. Am. Mosq. Control. Assoc. 2002, 18, 284–289. [Google Scholar]
- De Wispelaere, M.; Desprès, P.; Choumet, V. European Aedes albopictus and Culex pipiens are competent vectors for Japanese encephalitis virus. PLoS Negl. Trop. Dis. 2017, 11, e0005294. [Google Scholar] [CrossRef]
- Morchón, R.; Carretón, E.; González-Miguel, J.; Mellado-Hernández, I. Heartworm disease (Dirofilaria immitis) and their vectors in Europe—New distribution trends. Front. Physiol. 2012, 3, 196. [Google Scholar] [CrossRef] [Green Version]
- Neelakanta, G.; Sultana, H. Transmission-blocking vaccines: Focus on anti-vector vaccines against tick-borne diseases. Arch. Immunol. Ther. Exp. 2015, 63, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yooyangket, T.; Muangpat, P.; Polseela, R.; Tandhavanant, S.; Thanwisai, A.; Vitta, A. Identification of entomopathogenic nematodes and symbiotic bacteria from Nam Nao National Park in Thailand and larvicidal activity of symbiotic bacteria against Aedes aegypti and Aedes albopictus. PLoS ONE 2018, 13, e0195681. [Google Scholar] [CrossRef]
- Agumba, S.; Gimnig, J.E.; Ogonda, L.; Ombok, M.; Kosgei, J.; Munga, S.; Guyah, B.; Omondi, S.; Ochomo, E. Diagnostic dose determination and efficacy of chlorfenapyr and clothianidin insecticides against Anopheles malaria vector populations of western Kenya. Malar. J. 2019, 18, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chareonviriyaphap, T.; Bangs, M.J.; Suwonkerd, W.; Kongmee, M.; Corbel, V.; Ngoen-Klan, R. Review of insecticide resistance and behavioral avoidance of vectors of human diseases in Thailand. Parasites Vectors 2013, 6, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, J.P.; Ismail, H.M.; Chandor-Proust, A.; Paine, M.J. Role of cytochrome P450s in insecticide resistance: Impact on the control of mosquito-borne diseases and use of insecticides on Earth. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120429. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Matviishyn, T.M.; Husak, V.V.; Storey, J.M.; Storey, K.B. Pesticide toxicity: A mechanistic approach. EXCLI J. 2018, 17, 1101–1136. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Georgis, R. Entomopathogenic nematodes for control of insect pests above and below ground with comments on commercial production. J. Nematol. 2012, 44, 218–225. [Google Scholar]
- Abate, B.A.; Wingfield, M.J.; Slippers, B.; Hurley, B.P. Commercialisation of entomopathogenic nematodes: Should import regulations be revised? Biocontrol. Sci. Technol. 2017, 27, 149–168. [Google Scholar] [CrossRef]
- Labaude, S.; Griffin, C.T. Transmission success of entomopathogenic nematodes used in pest control. Insects 2018, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elgawad, M.M.M. Towards optimization of entomopathogenic nematodes for more service in the biological control of insect pests. Egypt. J. Biol. Pest. Control. 2019, 29, 77. [Google Scholar] [CrossRef]
- San-Blas, E.; Campos-Herrera, R.; Dolinski, C.; Monteiro, C.; Andaló, V.; Leite, L.G.; Rodríguez, M.G.; Morales-Montero, P.; Sáenz-Aponte, A.; Cedano, C.; et al. Entomopathogenic nematology in Latin America: A brief history, current research and future prospects. J. Invertebr. Pathol. 2019, 165, 22–45. [Google Scholar] [CrossRef]
- Askary, T.H.; Nermut, J.; Ahmad, M.J.; Ganai, M.A. Future thrusts in expanding the use of entomopathogenic and slug parasitic nematodes in agriculture. In Biocontrol Agents: Entomopathogenic and Slug Parasitic Nematodes; Abd-Elgawad, M.M.M., Askary, T.H., Coupland, J., Eds.; CAB International: Wallingford, UK, 2017; pp. 620–627. [Google Scholar]
- Caoili, B.L.; Latina, R.A.; Sandoval, R.F.C.; Orajay, J.I. Molecular identification of entomopathogenic nematode isolates from the philippines and their biological control potential against lepidopteran pests of corn. J. Nematol. 2018, 50, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Hazir, S.; Kaya, H.K.; Stock, S.P.; Keskin, N. Entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) for biological control of soil pests. Turk. J. Biol. 2003, 27, 181–202. [Google Scholar]
- Shapiro-Ilan, D.I.; Han, R.; Dolinksi, C. Entomopathogenic nematode production and application technology. J. Nematol. 2012, 44, 206–217. [Google Scholar] [PubMed]
- Bedding, R.A.; Molyneux, A.S.; Akhurst, R.J. Heterorhabditis spp., Neoaplectana spp., and Steinernema kraussei: Interspecific and intraspecific differences in infectivity for insects. Exp. Parasitol. 1983, 55, 249–257. [Google Scholar] [CrossRef]
- Bedding, R.A.; Molyneux, A.S. Penetration of insect cuticle by infective juveniles of Heterorhabditis spp. (Heterorhabditidae: Nematoda). Nematologica 1982, 28, 354–359. [Google Scholar] [CrossRef] [Green Version]
- Poinar, G.O., Jr.; Georgis, R. Characterization and field application of Heterorhabditis bacteriophora strain HP88 (Heterorhabditidae: Rhabditida). Revue Nématol. 1990, 13, 387–393. [Google Scholar]
- Burnell, A.M.; Stock, S.P. Heterorhabditis, Steinernema and their bacterial symbionts-lethal pathogens of insects. Nematology 2000, 2, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Dziedziech, A.; Shivankar, S.; Theopold, U. High-resolution infection kinetics of entomopathogenic nematodes entering Drosophila melanogaster. Insects 2020, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Dillman, A.R.; Chaston, J.M.; Adams, B.J.; Ciche, T.A.; Goodrich-Blair, H.; Stock, S.P.; Sternberg, P.W. An entomopathogenic nematode by any other name. PLoS Pathog. 2012, 8, e1002527. [Google Scholar] [CrossRef] [Green Version]
- Grewal, P.S.; Nardo, E.A.B.D.; Aguillera, M.M. Entomopathogenic nematodes: Potential for exploration and use in south America. Neotrop. Entomol. 2001, 30, 191–205. [Google Scholar] [CrossRef] [Green Version]
- Sharmila, R.; Subramanian, S.; Poornima, K. Host range of entomopathogenic nematodes. J. Entomol. Zool. Stud. 2018, 6, 1310–1312. [Google Scholar]
- Wong, J.; Stoddard, S.T.; Astete, H.; Morrison, A.C.; Scott, T.W. Oviposition site selection by the dengue vector Aedes aegypti and its implications for dengue control. PLoS Negl. Trop. Dis. 2011, 5, e1015. [Google Scholar] [CrossRef]
- Kampen, H.; Werner, D. Out of the bush: The asian bush mosquito Aedes japonicus japonicus (Theobald, 1901) (Diptera, Culicidae) becomes invasive. Parasites Vectors 2014, 7, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Getachew, D.; Tekie, H.; Gebre-Michael, T.; Balkew, M.; Mesfin, A. Breeding sites of Aedes aegypti: Potential dengue vectors in Dire Dawa, east Ethiopia. Interdiscip. Perspect. Infect. Dis. 2015, 2015, 706276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dom, N.C.; Madzlan, M.F.; Nur, S.; Hasnan, A.; Misran, N. Water quality characteristics of dengue vectors breeding containers. Int. J. Mosq. Res. 2016, 3, 2529. [Google Scholar]
- Poinar, G.O., Jr.; Kaul, H.N. Parasitism of the mosquito Culex pipiens by the nematode Heterorhabditis bacteriophora. J. Invertebr. Pathol. 1982, 39, 382–387. [Google Scholar] [CrossRef]
- Molta, N.B.; Hominick, W.M. Dose- and time-response assessments of Heterorhabditis heliothidis and Steinernema feltiae [Nem.: Rhabitida] against Aedes aegypti larvae. Entomophaga 1989, 34, 485–493. [Google Scholar] [CrossRef]
- Zohdy, Z.; Shamseldean, M.; El-Samiee, E.A.; Hamama, H.M. Efficacy of the Steinernematid and Heterorhabditid nematodes for controlling the mosquito, Culex quinquefasciatus Say (Diptera: Culicidae). J. Mosq. Res. 2013, 3, 33–44. [Google Scholar] [CrossRef]
- Peschiutta, M.; Cagnolo, S.R.; Almirón, W.R. Susceptibilidad de larvas de Aedes aegypti (Linnaeus) (Diptera: Culicidae) al nematodo entomopatógeno Heterorhabditis bacteriophora (Poinar) (Rhabditida: Heterorhabditidae). Rev. Soc. Entomol. Argent. 2014, 73, 99–108. [Google Scholar]
- Cagnolo, S.R.; Almirón, W.R. Capacity of the terrestrial entomopathogenic nematode Steinernema rarum (Rhabditida: Steinernematidae) to parasite Culex apicinus larvae (Diptera: Culicidae). Rev. Soc. Entomol. Argent. 2010, 69, 141–145. [Google Scholar]
- Silva, B.; Almeida, A.M.; Dolinski, C.; Souza, R.M. Efficacy of Heterorhabdits indica LPP35 against Aedes aegypti in domiciliary oviposition sites. J. Nematol. 2019, 51, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Edmunds, C.V.; Wilding, C.S.; Rae, R. Susceptibility of Chironomus plumosus larvae (Diptera: Chironomidae) to entomopathogenic nematodes (Rhabditida: Steinernematidae and Heterorhabditidae): Potential for control. Eur. J. Entomol. 2017, 114, 526–532. [Google Scholar] [CrossRef] [Green Version]
- Ulvedal, C.; Bertolotti, M.A.; Cagnolo, S.R.; Almirón, W.R. Ensayos de sensibilidad de larvas de Aedes aegypti y Culex quinquefasciatus frente al nematodo Heterorhabditis bacteriophora en condiciones de laboratorio. Biomédica 2017, 37, 67–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.Y.; Tang, L.C.; Pai, C.F.; Hsiao, W.F.; Briscoe, B.R.; Hou, R.F. A new isolate of the entomopathogenic nematode, Steinernema abbasi (Nematoda: Steinernematidae), from Taiwan. J. Invertebr. Pathol. 2001, 77, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Beresky, M.A.; Hall, D.W. The influence of phenylthiourea on encapsulation, melanization, and survival in larvae of the mosquito Aedes aegypti parasitized by the nematode Neoaplectana carpocapsae. J. Invertebr. Pathol. 1977, 29, 74–80. [Google Scholar] [CrossRef]
- Welch, H.E.; Bronskill, J.F. Parasitism of mosquito larvae by the nematode, DD136 (Nematoda: Neoaplectanidae). Can. J. Zool. 1962, 40, 1263–1268. [Google Scholar] [CrossRef]
- Dadd, R.H. Size limitations on the infectibility of mosquito larvae by nematodes during filter-feed. J. Invertebr. Pathol. 1971, 18, 246–251. [Google Scholar] [CrossRef]
- Fenton, A.; Norman, R.; Fairbairn, J.P.; Hudson, P.J. Evaluating the efficacy of entomopathogenic nematodes for the biological control of crop pests: A nonequilibrium approach. Am. Nat. 2001, 158, 408–425. [Google Scholar] [CrossRef]
- Chaudhary, M.Z.; Majeed, S.; Tayyib, M.; Javed, N.; Farzand, A.; Moosa, A.; Shehzad, M.; Mushtaq, F. Antagonistic potential of Steinernema kraussei and Heterorhabditis bacteriophora against dengue fever mosquito Aedes aegypti. J. Entomol. Zool. Stud. 2017, 5, 865–869. [Google Scholar]
- Dilipkumar, A.; Raja Ramalingam, K.; Chinnaperumal, K.; Govindasamy, B.; Paramasivam, D.; Dhayalan, A.; Pachiappan, P. Isolation and growth inhibition potential of entomopathogenic nematodes against three public health important mosquito vectors. Exp. Parasitol. 2019, 197, 76–84. [Google Scholar] [CrossRef]
- Tsai, M.H.; Tang, L.C.; Hou, R.F. The bacterium associated with the entomopathogenic nematode Steinernema abbasi (Nematoda: Steinernematidae) isolated from Taiwan. J. Invertebr. Pathol. 2008, 99, 242–245. [Google Scholar] [CrossRef]
- Vitta, A.; Thimpoo, P.; Meesil, W.; Yimthin, T.; Fukruksa, C.; Polseela, R.; Mangkit, B.; Tandhavanant, S.; Thanwisai, A. Larvicidal activity of Xenorhabdus and Photorhabdus bacteria against Aedes aegypti and Aedes albopictus. Asian Pac. J. Trop. Biomed. 2018, 8, 31–36. [Google Scholar] [CrossRef]
- Gaugler, R.; Kaplan, B.; Alvarado, C.; Montoya, J.; Ortega, M. Assessment of Bacillus thuringiensis serotype 14 and Steinernema feltiae [Nematoda: Steinernematidae] for control of the Simulium vectors of onchocerciasis in Mexico. Entomophaga 1983, 28, 309–315. [Google Scholar] [CrossRef]
- Poinar, G.O., Jr. Taxonomy and biology of Steinernematidae and Heterorhabdititidae. In Entomopathogenic Nematodes in Biological Control; CRC Press: Boca Raton, FL, USA, 1990; pp. 23–61. [Google Scholar]
- Walsh, K.T.; Webster, J.M. Interaction of microbial populations in Steinernema (Steinernematidae, Nematoda) infected Galleria mellonella larvae. J. Invertebr. Pathol. 2003, 83, 118–126. [Google Scholar] [CrossRef]
- Poinar, G.O., Jr.; Grewal, P.S. History of entomopathogenic nematology. J. Nematol. 2012, 44, 153–161. [Google Scholar]
- Peters, A.; Ehlers, R.U. Susceptibility of leather jackets (Tipula paludosa and T. oleracea, Tipulidae: Nematocera) to the entomopathogenic nematode Steinernema feltiae. J. Invertebr. Pathol. 1994, 69, 163–171. [Google Scholar] [CrossRef]
- Abu Hatab, M.; Selvan, S.; Gaugler, R. Role of proteases in penetration of insect gut by the entomopathogenic nematode Steinernema glaseri (Nematoda: Steinernematidae). J. Invertebr. Pathol. 1995, 66, 125–130. [Google Scholar] [CrossRef]
- Chen, C.C.; Laurence, B.R. An ultrastructural study on the encapsultion of microfilaria of Brugia pahangi in the haemocoel of Anopheles quadrimaculatus. Int. J. Parasitol. 1985, 15, 421–428. [Google Scholar] [CrossRef]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef] [Green Version]
- Schmit, A.R.; Ratcliffe, N.A. The encapsulation of foreign tissue implants in Galleria mellonella larvae. J. Insect. Physiol. 1977, 23, 175–184. [Google Scholar] [CrossRef]
- Götz, P.; Vey, A. Humoral encapsulation in Diptera (Insecta): Defense reactions of Chironomus larvae against fungi. Parasitology 1974, 68, 193–205. [Google Scholar] [CrossRef]
- Browne, N.; Heelan, M.; Kavanagh, K. An analysis of the structural and functional similarities of insect hemocytes and mammalian phagocytes. Virulence 2013, 4, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.T.; Hou, R.F.; Chen, C.C. Formation of basement membrane-like structure terminates the cellular encapsulation of microfilariae in the haemocoel of Anopheles quadrimaculatus. Parasitology 1998, 116, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Barreaux, A.M.; Barreaux, P.; Koella, J.C. Overloading the immunity of the mosquito Anopheles gambiae with multiple immune challenges. Parasites Vectors 2016, 9, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elawad, A.S.; Ahmad, W.; Reid, A.P. Steinernema abbasi sp. n. (Nematoda: Steinernematidae) from the Sultanate of Oman. Fundam. Appl. Nematol. 1997, 20, 435–442. [Google Scholar]
- Aasha, R.; Chaubey, A.K.; Bhat, A.H. Notes on Steinernema abbasi (Rhabditida: Steinernematidae) strains and virulence tests against lepidopteran and coleopterans pests. J. Entomol. Zool. Stud. 2019, 7, 954–964. [Google Scholar]
- Sunanda, B.S. Effect of temperature on the life cycle of entomopathogenic nematodes, Steinernema abbasi and Heterorhabditis indica. J. Biol. Control. 2009, 23, 185–189. [Google Scholar] [CrossRef]
- Yoshida, M. A new distribution record of Steinernema abbasi Elawad, Ahmad and Reid, 1997 (Rhabditida: Steinernematidae) in Japan with a note on its pathogenicity against the turnip moth larvae, Agrotis segetum (Lepidoptera: Noctuidae). Jpn. J. Nematol. 2007, 37, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Nagesh, M.; Balachander, M.; Shivalingaswamy, T.M.; Patil, J.; Shylesha, A.N.; Raghavendra, A. Variability in foraging behaviour, thermal requirement and virulence of entomopathogenic nematodes against sod webworm, Herpetogramma phaeopteralis Gueneè (Lepidoptera: Crambidae). J. Biol. Control. 2019, 33, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Hussaini, S.S.; Singh, S.P.; Parthasarathy, R.; Shakeela, V. Storage effects on activity of native Steinernema and Heterorhabditis spp. Indian J. Nematol. 2000, 30, 231–232. [Google Scholar]
- Sunanda, B.S.; Siddiqui, A.U.; Sharma, S. Effect of temperature on longevity of entomopathogenic nematodes, Steinernema abbasi and Heterorhabditis indica. Indian J. Nematol. 2012, 42, 17–19. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nematode Species | Mosquito Instars | Lethal Time (LT50 and LT90) Values (h) | |||
|---|---|---|---|---|---|
| 1 × 103 (IJs/mL) a | 1 × 104 (IJs/mL) | ||||
| LT50 | LT90 | LT50 | LT90 | ||
| S. abbasi | 3rd | 37.9 (35.7–40.1) b | 69.1 (67.2–70.9) | 31.2 (30.2–32.2) | 60.6 (59.7–61.6) |
| 4th | 47.1 (44.6–49.6) | >72 c | 34.9 (33.3–36.5) | 57.1 (56.1–58.1) | |
| S. carpocapsae | 3rd | 46.4 (45.3–47.4) | >72 | 31.6 (29.8–33.5) | 62.9 (59.8–66) |
| 4th | 47.8 (45.6–49.9) | >72 | 26.9 (25.7–28.1) | 53.1 (51.4–54.8) | |
| Number of Encapsulated Nematode in a Mosquito Larva | Number of Mosquito Larvae | |
|---|---|---|
| Dead | Survived | |
| 0 | 0 | 7 |
| 1 | 25 | 6 |
| 2 | 47 | 4 |
| 3 | 28 | 0 |
| 4 | 25 | 1 |
| 5 | 10 | 0 |
| 6 | 9 | 0 |
| 7 | 7 | 0 |
| 8 | 3 | 0 |
| 9 | 2 | 0 |
| 10 | 1 | 0 |
| 11 | 1 | 0 |
| 13 | 1 | 0 |
| 14 | 1 | 0 |
| 16 | 1 | 0 |
| 17 | 1 | 0 |
| 20 | 1 | 0 |
| 31 | 1 | 0 |
| Total | 164 | 18 |
| Percentage (%) | 90.11 | 9.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.-T.; Chen, T.-L.; Hou, R.F.; Chen, C.-C.; Tu, W.-C. The Invasion and Encapsulation of the Entomopathogenic Nematode, Steinernema abbasi, in Aedes albopictus (Diptera: Culicidae) Larvae. Insects 2020, 11, 832. https://doi.org/10.3390/insects11120832
Liu W-T, Chen T-L, Hou RF, Chen C-C, Tu W-C. The Invasion and Encapsulation of the Entomopathogenic Nematode, Steinernema abbasi, in Aedes albopictus (Diptera: Culicidae) Larvae. Insects. 2020; 11(12):832. https://doi.org/10.3390/insects11120832
Chicago/Turabian StyleLiu, Wei-Ting, Tien-Lai Chen, Roger F. Hou, Cheng-Chen Chen, and Wu-Chun Tu. 2020. "The Invasion and Encapsulation of the Entomopathogenic Nematode, Steinernema abbasi, in Aedes albopictus (Diptera: Culicidae) Larvae" Insects 11, no. 12: 832. https://doi.org/10.3390/insects11120832