Characteristics of the Two Asian Bumblebee Species Bombus friseanus and Bombus breviceps (Hymenoptera: Apidae)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Investigating Food Plants
2.2. Monitoring the Life History
2.3. Bumblebees Laboratory Rearing
2.4. Recording the Colony Development
2.5. Observing the Mating Behaviour
3. Results
3.1. The Food Plants of B. friseanus and B. breviceps
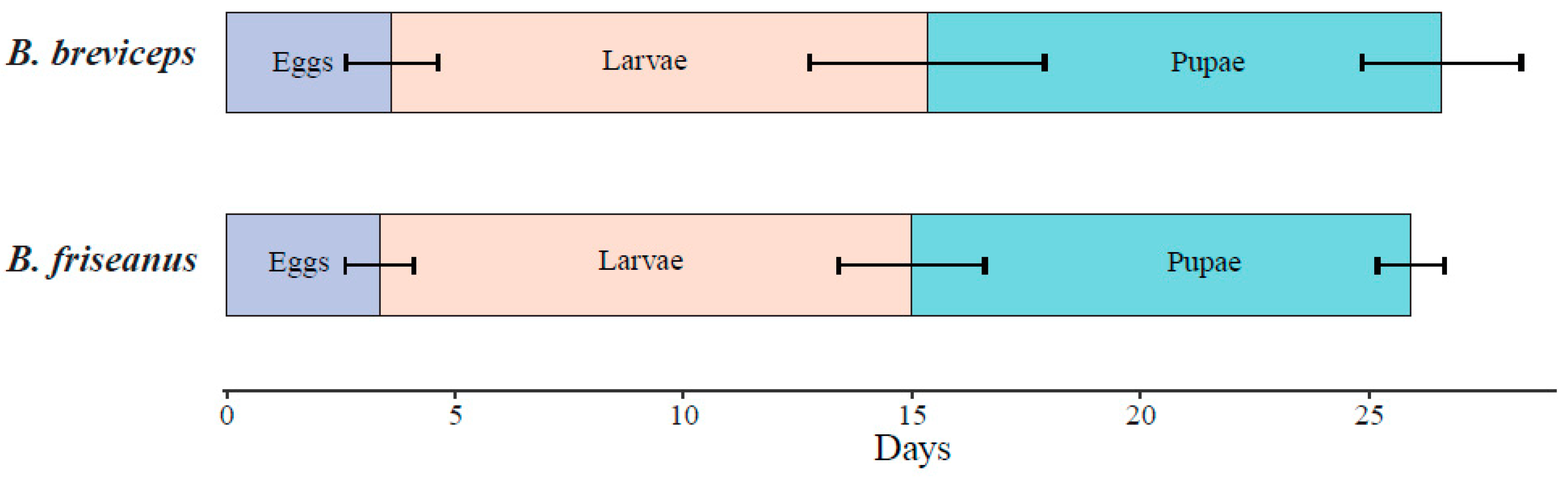
3.2. The Life History of B. friseanus and B. breviceps
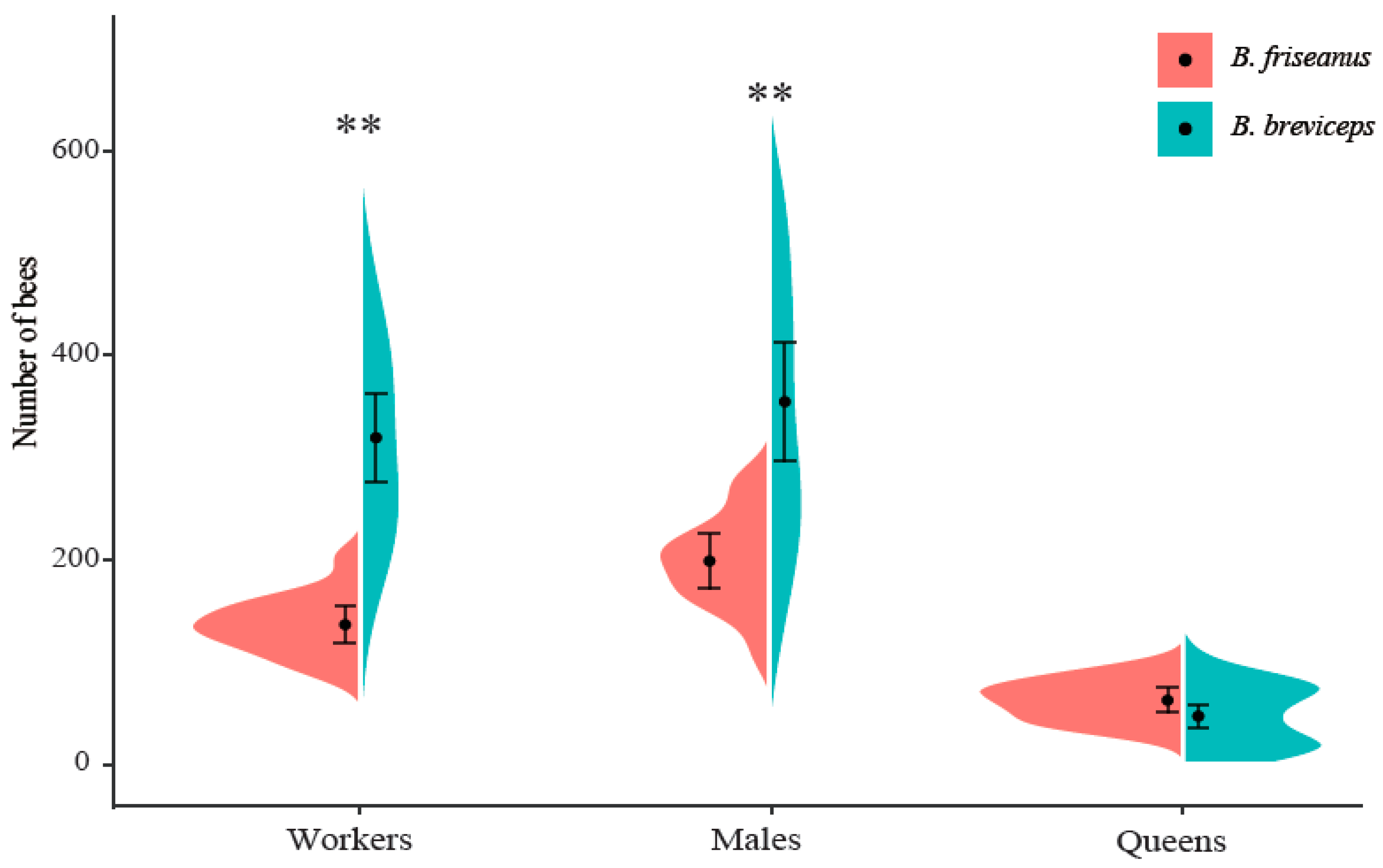
3.3. The Colony Development of B. friseanus and B. breviceps
3.4. The Mating Behaviour of B. friseanus and B. breviceps
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Williams, P.; Huang, J.; An, J. Bear wasps of the Middle Kingdom: A decade of discovering China’s bumblebees. Antenna 2017, 41, 21–24. [Google Scholar]
- Beekman, M.; Van Stratum, P. Does the diapause experience of bumblebee queens Bombus terrestris affect colony characteristics? Ecol. Entomol. 2000, 25, 1–6. [Google Scholar] [CrossRef]
- Velthuis, H.H.W.; Van Doorn, A. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Rasmont, P.; Coppee, A.; Michez, D.; De Meulemeester, T. An overview of the Bombus terrestris (L. 1758) subspecies (Hymenoptera: Apidae). Ann. Soc. Entomol. Fr. 2008, 44, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Hingston, A.B.; Marsden-Smedley, J.; Driscoll, D.A.; Corbett, S.; Fenton, J.; Anderson, R.; Plowman, C.; Mowling, F.; Jenkin, M.; Matsui, K.; et al. Extent of invasion of Tasmanian native vegetation by the exotic bumblebee Bombus terrestris (Apoidea: Apidae). Austral Ecol. 2002, 27, 162–172. [Google Scholar] [CrossRef]
- Matsumura, C.; Yokoyama, J.; Washitani, I. Invasion status and potential ecological impacts of an invasive alien bumblebee, Bombus terrestris L. (Hymenoptera: Apidae) naturalized in Southern Hokkaido, Japan. Glob. Environ. Res. 2004, 8, 51–66. [Google Scholar]
- Inoue, M.N.; Yokoyama, J.; Washitani, I. Displacement of Japanese native bumblebees by the recently introduced Bombus terrestris (L.) (Hymenoptera: Apidae). J. Insect Conserv. 2008, 12, 135–146. [Google Scholar] [CrossRef]
- Tsuchida, K.; Kondo, N.I.; Inoue, M.N.; Goka, K. Reproductive disturbance risks to indigenous Japanese bumblebees from introduced Bombus terrestris. Appl. Entomol. Zool. 2010, 45, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Dohzono, I.; Kunitake, Y.K.; Yokoyama, J.; Goka, K. Alien bumble bee affects native plant reproduction through interactions with native bumble bees. Ecology 2008, 89, 3082–3092. [Google Scholar] [CrossRef]
- Sáez, A.; Morales, C.L.; Garibaldi, L.A.; Aizen, M.A. Invasive bumble bees reduce nectar availability for honey bees by robbing raspberry flower buds. Basic Appl. Ecol. 2017, 19, 26–35. [Google Scholar] [CrossRef]
- Jones, C.M.; Brown, M.J.F. Parasites and genetic diversity in an invasive bumblebee. J. Anim. Ecol. 2014, 83, 1428–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid-Hempel, R.; Eckhardt, M.; Goulson, D.; Heinzmann, D.; Lange, C.; Plischuk, S.; Escudero, L.R.; Salathe, R.; Scriven, J.J.; Schmid-Hempel, P. The invasion of southern South America by imported bumblebees and associated parasites. J. Anim. Ecol. 2014, 83, 823–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanbe, Y.; Okada, I.; Yoneda, M.; Goka, K.; Tsuchida, K. Interspecific mating of the introduced bumblebee Bombus terrestris and the native Japanese bumblebee Bombus hypocrita sapporoensis results in inviable hybrids. Naturwissenschaften 2008, 95, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, K.; Yamaguchi, A.; Kanbe, Y.; Goka, K. Reproductive interference in an introduced bumblebee: Polyandry may mitigate negative reproductive impact. Insects 2019, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfarlane, R.P.; Patten, K.D.; Royce, L.A.; Wyatt, B.K.W.; Mayer, D.F. Management potential of sixteen North American bumble bee species. Melanderia 1994, 50, 1–12. [Google Scholar]
- Mah, Y.I.; Lee, M.Y.; Bilinski, M. Some charateristics of korean indigenous bumblebee species (hymenoptera; Bombus spp.) under laboratory conditions. Acta Hortic. 2001, 561, 287–291. [Google Scholar] [CrossRef]
- Yoon, H.J.; Kim, S.E.; Kim, Y.S. Temperature and humidity favorable for colony development of the indoor-reared bumblebee, Bombus ignitus. Appl. Entomol. Zool. 2002, 37, 419–423. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Mitsuhata, M.; Sasaki, M. Use of introduced Bombus terrestris worker helpers for rapid development of Japanese native B. hypocrita colonies (Hymenoptera, Apidae). Appl. Entomol. Zool. 1994, 29, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Hannan, M.D.; Maeta, Y.; Hoshikawa, K. Colony development of two species of Japanese bumble bees Bombus ignitus and Bombus hypocrita reared under artificial condition. Jpn. J. Entomol. 1997, 65, 343–354. [Google Scholar]
- Asada, S.; Ono, M. Difference in colony development of two Japanese bumblebees, Bombus hypocrita and B. ignitus (Hymenoptera: Apidae). Appl. Entomol. Zool. 2000, 35, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; An, J. Species diversity, pollination application and strategy for conservation of the bumblebees of China. Biodivers. Sci. 2018, 26, 486–497. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Z.; Huang, J.; Yuan, X.; Ding, G.; An, J. Queen traits and colony size of four bumblebee species of China. Insects Soc. 2018, 65, 537–547. [Google Scholar] [CrossRef]
- Williams, P.; Tang, Y.; Yao, J.; Cameron, S. The bumblebees of Sichuan (Hymenoptera: Apidae, Bombini). Syst. Biodivers. 2009, 7, 101–189. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Zhang, X.; Huang, J.; Song, W.; Zhang, H.; Luo, W.; An, J. Biogeography and species diversity of bumblebees in Yunnan, Southwest China. Chin. J. Appl. Entomol. 2018, 55, 1045–1053. [Google Scholar] [CrossRef]
- Tang, Y.; Xie, J.; Sun, H. Pollination ecology of Pedicularis muscoides H. L. Li subsp. himalayca Yamazaki from alpine areas of western Sichuan, China. Arct. Antarct. Alp. Res. 2007, 39, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Li, D.; Wang, H. Highly efficient pollination by bumblebees ensures seed production in Pedicularis lachnoglossa (Orobanchaceae), an early-flowering Himalayan plant. J. Syst. Evol. 2012, 50, 218–226. [Google Scholar] [CrossRef]
- Corbet, S.A.; Huang, S.Q. Buzz pollination in eight bumblebee-pollinated Pedicularis species: Does it involve vibration-induced triboelectric charging of pollen grains? Ann. Bot. 2014, 114, 1665–1674. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Jin, X.; Wang, Q.; Yang, C.; Inouye, D.W. Nectar replenishment maintains the neutral effects of nectar robbing on female reproductive success of Salvia przewalskii (Lamiaceae), a plant pollinated and robbed by bumble bees. Ann. Bot. 2017, 119, 1053–1059. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Jin, X.; Inouye, D.W.; Wang, Q.; Yang, C. Variation in composition of two bumble bee species across communities affects nectar robbing but maintains pollinator visitation rate to an alpine plant, Salvia przewalskii. Ecol. Entomol. 2018, 43, 363–370. [Google Scholar] [CrossRef]
- Deka, T.N.; Sudharshan, M.R.; Saju, K.A. New record of bumble bee, Bombus breviceps Smith as a pollinator of large cardamom. Curr. Sci. 2011, 100, 926–928. [Google Scholar]
- Deka, T.N.; Gudade, B.; Saju, K.A.; Vijayan, A.; Chhetri, P. Foraging behaviour of bumble bee (Bombus breviceps) in large cardamom (Amomum subulatum Roxb.) at Dzongu, North Sikkim. Ecol. Env. Cons. 2014, 20, 197–199. [Google Scholar]
- Goulson, D.; Hawson, S.A.; Stout, J.C. Foraging bumblebees avoid flowers already visited by conspecifics or by other bumblebee species. Anim. Behav. 1998, 55, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, R.L. Nestmate recognition as an inbreeding avoidance mechanism in bumble bees (Hymenoptera: Apidae). J. Kans. Entomol. Soc. 1992, 65, 238–243. [Google Scholar]
- Duvoisin, N.; Baer, B.; Schmid-Hempel, P. Sperm transfer and male competition in a bumblebee. Anim. Behav. 1999, 58, 743–749. [Google Scholar] [CrossRef]
- Brown, M.J.F.; Baer, B.; Schmid-Hempel, R.; Schmid-Hempel, P. Dynamics of multiple-mating in the bumble bee Bombus hypnorum. Insects Soc. 2002, 49, 315–319. [Google Scholar] [CrossRef]
- Estoup, A.; Scholl, A.; Pouvreau, A.; Solignac, M. Monoandry and polyandry in bumble bees (Hymenoptera; Bombinae) as evidenced by highly variable microsatellites. Mol. Ecol. 1995, 4, 89–93. [Google Scholar] [CrossRef]
- Schmid-Hempel, R.; Schmid-Hempel, P. Female mating frequencies in Bombus spp. from Central Europe. Insects Soc. 2000, 47, 36–41. [Google Scholar] [CrossRef]
- Kokuvo, N.; Toquenaga, Y.; Goka, K. Effective paternity in natural colonies of Japanese native bumble bees. Ecol. Res. 2009, 24, 1111–1115. [Google Scholar] [CrossRef]
- Paxton, R.J.; Thoren, P.A.; Estoup, A.; Tengö, J. Queen-worker conflict over male production and the sex ratio in a facultatively polyandrous bumblebee, Bombus hypnorum: The consequences of nest usurpation. Mol. Ecol. 2001, 10, 2489–2498. [Google Scholar] [CrossRef]
- Payne, C.M.; Laverty, T.M.; Lachance, M.A. The frequency of multiple paternity in bumble bee (Bombus) colonies based on microsatellite DNA at the B10 locus. Insects Soc. 2003, 50, 375–378. [Google Scholar] [CrossRef]
- Baer, B.; Morgan, E.D.; Schmid-Hempel, P. A nonspecific fatty acid within the bumblebee mating plug prevents females from remating. Proc. Natl. Acad. Sci. USA 2001, 98, 3926–3928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauter, A.; Brown, M.J.F. To copulate or not? The importance of female status and behavioural variation in predicting copulation in a bumblebee. Anim. Behav. 2001, 62, 221–226. [Google Scholar] [CrossRef]
- Inouye, D.W. The effect of proboscis and corolla tube lengths on patterns and rates of flower visitation by bumblebees. Oecologia 1980, 45, 197–201. [Google Scholar] [CrossRef]
- Irwin, R.E.; Bronstein, J.L.; Manson, J.S.; Richardson, L. Nectar robbing: Ecological and evolutionary perspectives. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 271–292. [Google Scholar] [CrossRef]
- Irwin, R.E.; Maloof, J.E. Variation in nectar robbing over time, space, and species. Oecologia 2002, 133, 525–533. [Google Scholar] [CrossRef]
- Maloof, J.; Inouye, D. Are nectar robbers cheaters or mutualists? Ecology 2000, 81, 2651–2661. [Google Scholar] [CrossRef]
- Gösterit, A.; Gürel, F. Comparison of development patterns of imported and native Bombus terrestris L. (Hymenoptera: Apidae) colonies in the Mediterranean coastal region. Turk. J. Vet. Anim. Sci. 2005, 29, 393–398. [Google Scholar]
- Müller, C.B.; Schmid-Hempel, P. Variation in life-history pattern in relation to worker mortality in the bumble-bee, Bombus lucorum. Funct. Ecol. 1992, 6, 48–56. [Google Scholar] [CrossRef]
- Cnaani, J.; Schmid-Hempel, R.; Schmidt, J.O. Colony development, larval development and worker reproduction in Bombus impatiens Cresson. Insects Soc. 2002, 49, 164–170. [Google Scholar] [CrossRef]


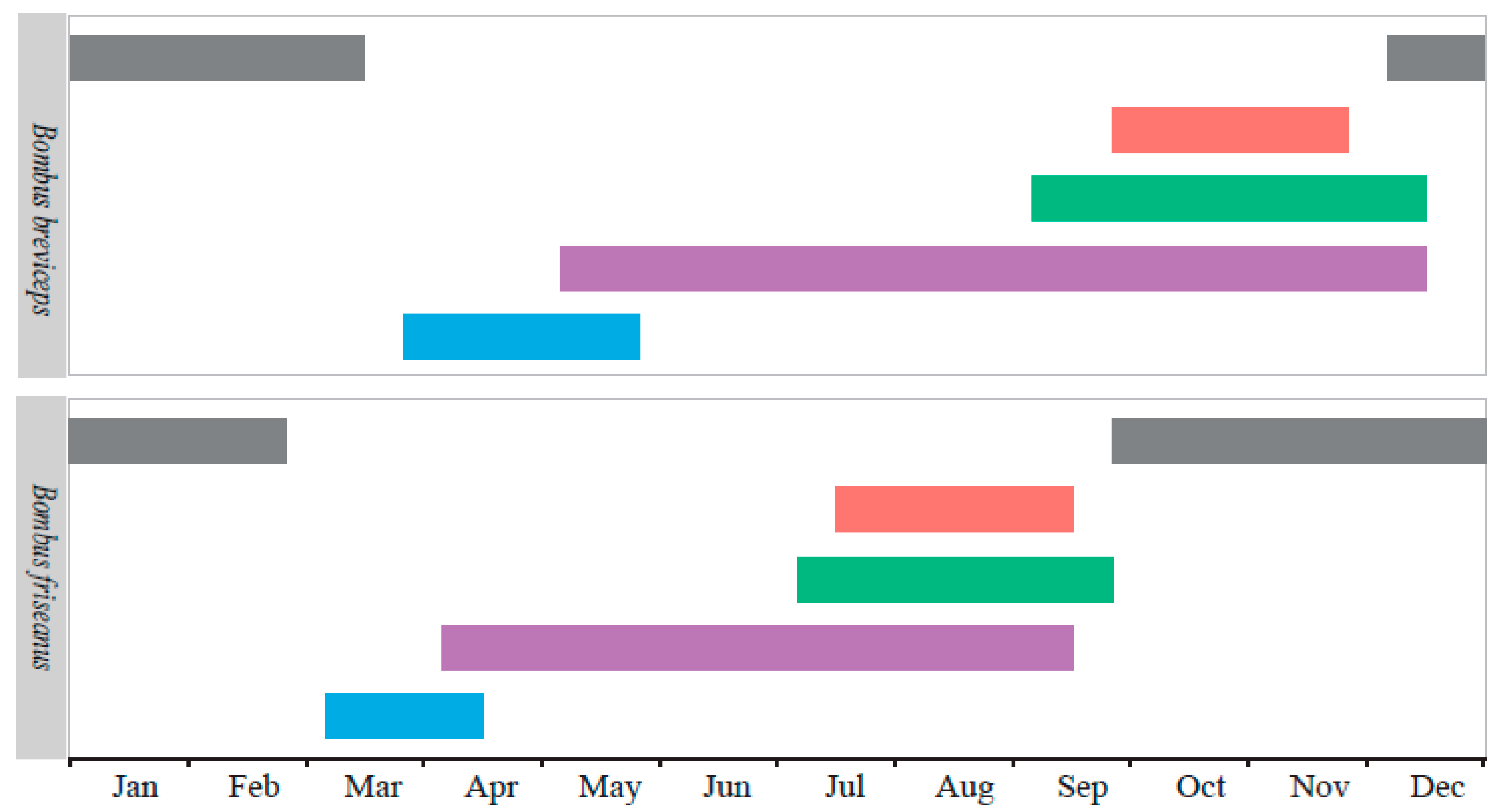
 ) indicate the seasonal occurrence of hibernated queens, purple bars (
) indicate the seasonal occurrence of hibernated queens, purple bars (  ) for workers, green bars (
) for workers, green bars (  ) for males, red bars (
) for males, red bars (  ) for gynes, and gray bars (
) for gynes, and gray bars (  ) for mated queens in hibernation.
) indicate the seasonal occurrence of hibernated queens, purple bars ( ) for workers, green bars ( ) for males, red bars ( ) for gynes, and gray bars ( ) for mated queens in hibernation.
) for mated queens in hibernation.
) indicate the seasonal occurrence of hibernated queens, purple bars ( ) for workers, green bars ( ) for males, red bars ( ) for gynes, and gray bars ( ) for mated queens in hibernation.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bumblebee Species | Number of Colonies | Initial Oviposition | First Worker Emergence | Colony Size of 30 Workers | Colony Size of 60 Workers | Colony Size of 80 Workers | First Male Emergence | First Gyne Emergence | Colony Lifetime |
|---|---|---|---|---|---|---|---|---|---|
| Bombus friseanus | 13 | 8.9 ± 7.7 a | 25.4 ± 1.8 a | 38.8 ± 5.0 a | 46.2 ± 5.2 a | 53.6 ± 5.5 a | 60.6 ± 5.1 a | 73.5 ± 5.9 a | 132.5 ± 15.5 a |
| Bombus breviceps | 20 | 13.6 ± 7.0 a | 26.5 ± 8.1 a | 62.1 ± 13.2 b | 76.2 ± 15.0 b | 87.8 ± 17.8 b | 120.4 ± 12.7 b | 141.2 ± 16.0 b | 217.8 ± 21.7 b |
| Bumblebee Species | Mating Success | Mating Duration | Queens Re-mated | |||
|---|---|---|---|---|---|---|
| Number of Queens | Mating Rate (%) | Number of Mating Pairs | Mating Duration ± S. E. (min) | Number of Queens Observed | Queens Re-mated (%) | |
| Bombus friseanus | 259 | 62.96 a | 30 | 27.44 ± 10.97 a | 149 | 0 |
| Bombus breviceps | 216 | 57.53 a | 30 | 1.54 ± 0.62 b | 136 | 10.07 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, C.; Ding, G.; Huang, J.; Zhang, X.; Miao, C.; An, J. Characteristics of the Two Asian Bumblebee Species Bombus friseanus and Bombus breviceps (Hymenoptera: Apidae). Insects 2020, 11, 163. https://doi.org/10.3390/insects11030163
Liang C, Ding G, Huang J, Zhang X, Miao C, An J. Characteristics of the Two Asian Bumblebee Species Bombus friseanus and Bombus breviceps (Hymenoptera: Apidae). Insects. 2020; 11(3):163. https://doi.org/10.3390/insects11030163
Chicago/Turabian StyleLiang, Cheng, Guiling Ding, Jiaxing Huang, Xuewen Zhang, Chunhui Miao, and Jiandong An. 2020. "Characteristics of the Two Asian Bumblebee Species Bombus friseanus and Bombus breviceps (Hymenoptera: Apidae)" Insects 11, no. 3: 163. https://doi.org/10.3390/insects11030163
APA StyleLiang, C., Ding, G., Huang, J., Zhang, X., Miao, C., & An, J. (2020). Characteristics of the Two Asian Bumblebee Species Bombus friseanus and Bombus breviceps (Hymenoptera: Apidae). Insects, 11(3), 163. https://doi.org/10.3390/insects11030163