Morphological and Molecular Characterization of Lema bilineata (Germar), a New Alien Invasive Leaf Beetle for Europe, with Notes on the Related Species Lema daturaphila Kogan & Goeden



Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Monitoring in Campania
2.2. Color and Anatomical Observations
2.3. Molecular Data
2.4. Host Plant Range
3. Results
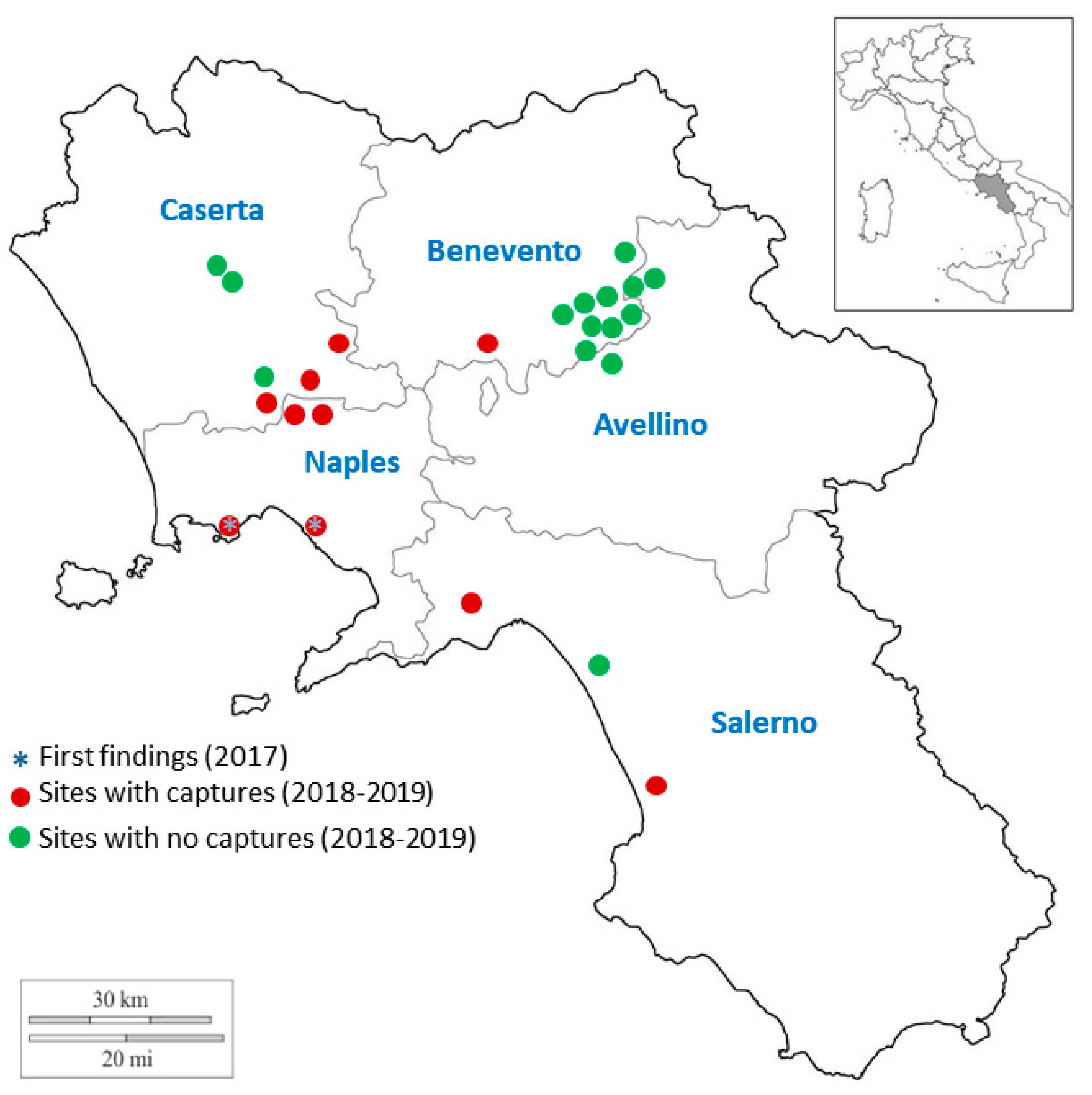
3.1. Field Monitoring in Campania
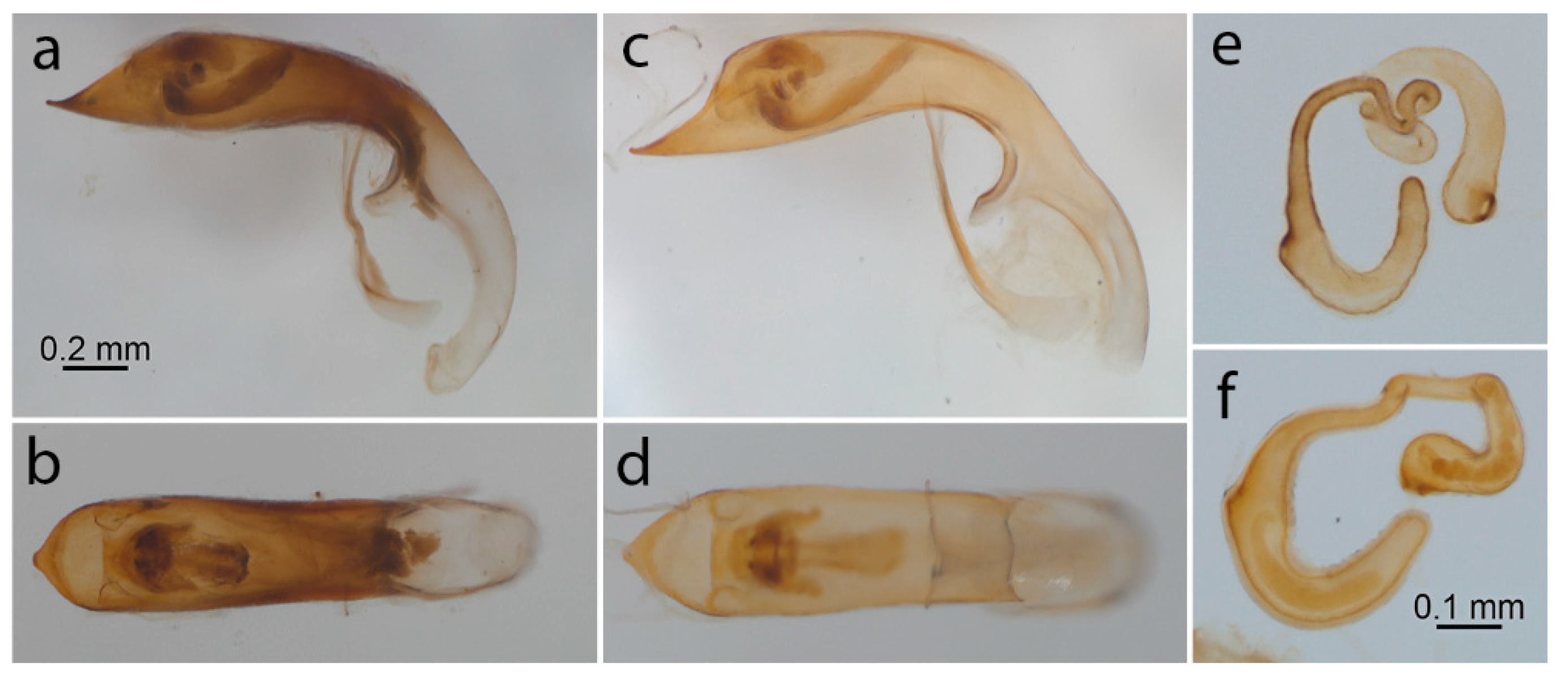
3.2. Color and Anatomical Observations
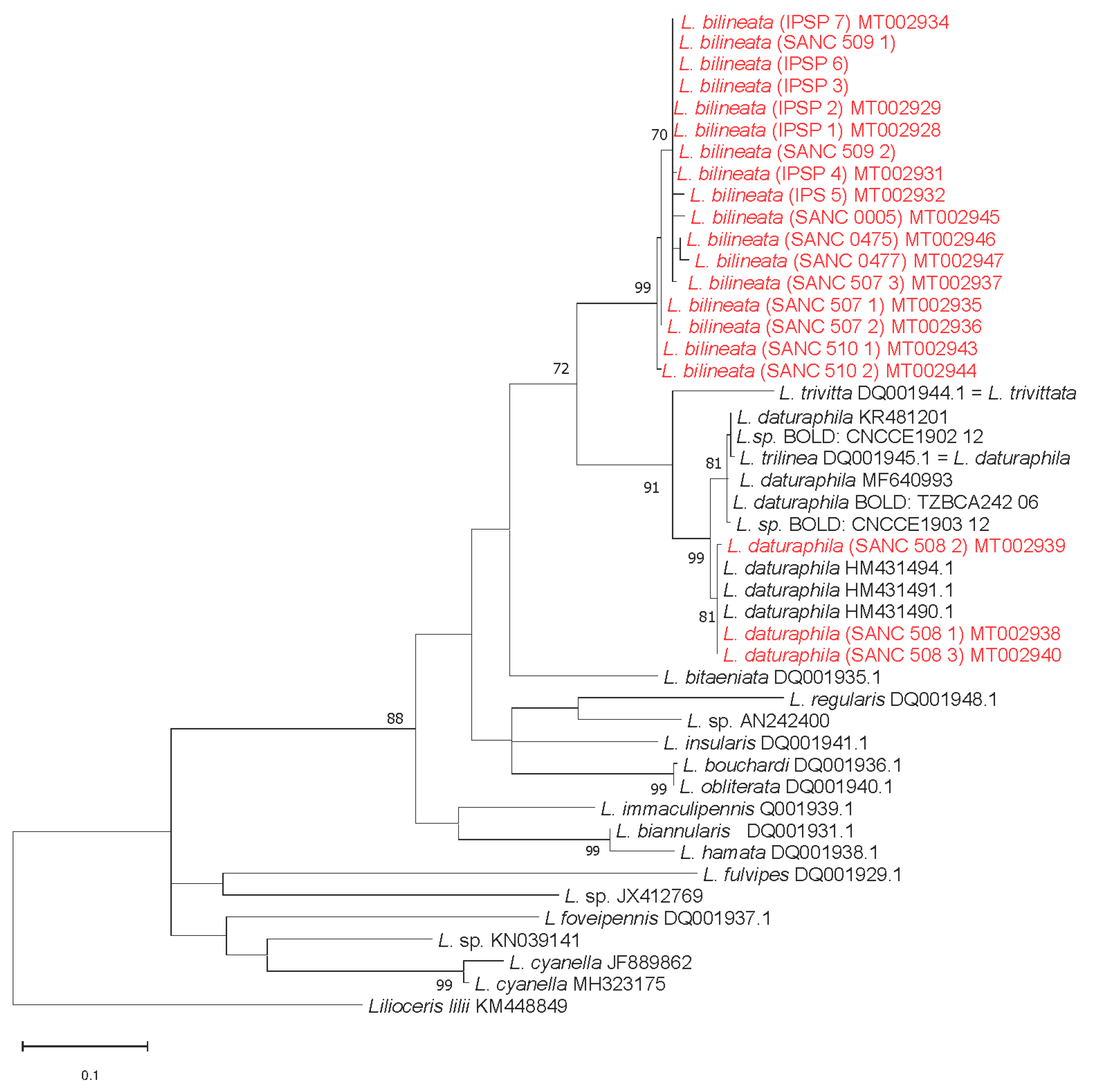
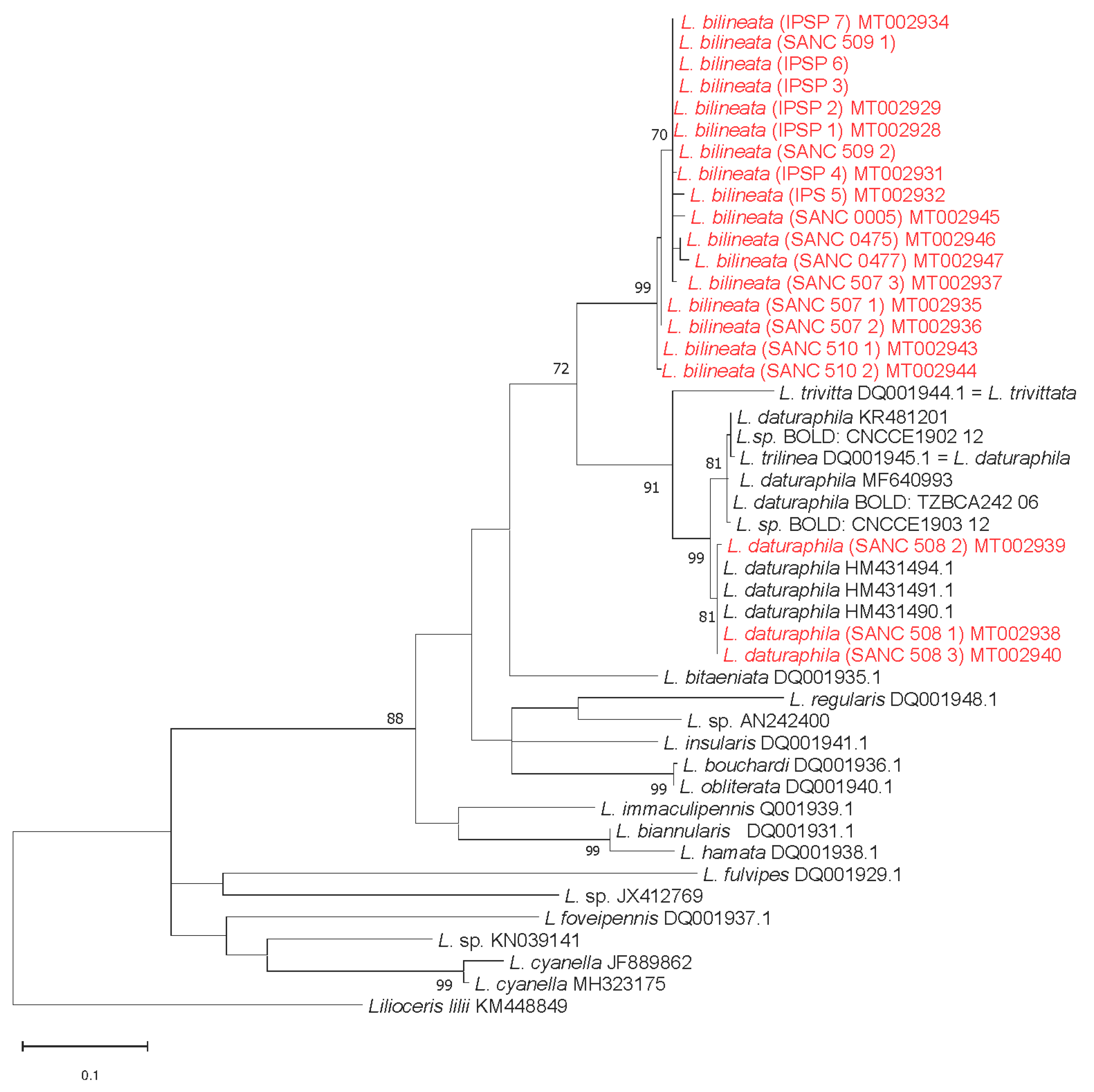
3.3. Molecular Analyses
3.4. Host Plant Range
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Servizio Fitosanitario della Regione Campania; Assssorato Agricoltura: Regione Campania, Italy; Available online: http://agricoltura.regione.campania.it/difesa/lema_bilineata.html (accessed on 15 November 2019).
- Marelli, C.A. Importancia de investigar en la Argentina los parásitos de Lema bilineata Germar. Convertida en peste de las plantaciones de tabaco en Sud Africa y su posible aclimatación en aquellos estados. Rev. Soc. Entomol. Argent. 1927, 2, 47–54. [Google Scholar]
- González, R.H. Insectos y Ácaros de Importancia Agrícola y Cuarentenaria en Chile; Universidad de Chile, BASF: Santiago, Chile, 1989; pp. 21, 184. [Google Scholar] [CrossRef]
- Van der Merwe, C.P. The tobacco slug (Lema bilineata Germar). J. Dep. Agric. Union S. Afr. 1921, 1, 28–38. [Google Scholar]
- Blair, B.W. Insect and Mite Pests of Tobacco in Zimbabwe: Description, Biology and Damage; Tobacco Research Board: Harare, Zimbabwe, 1990; p. 15. [Google Scholar]
- Stevens, M.M.; Stanton, R.A.; Wu, H.; Sampson, B.; Weir, T.A.; Reid, C.A.M.; Mo, J. Detection of Lema bilineata Germar (Coleoptera: Chrysomelidae) in Australia. Gen. Appl. Ent. 2010, 39, 1–3. Available online: https://researchoutput.csu.edu.au/ws/portalfiles/portal/8779894/Stevens+et+al+2010.pdf (accessed on 3 September 2017).
- Orrego Aravena, R. Insectos de la Pampa (Coleópteros); Consejo Provincial de Difusión; Gobierno del Pueblo, Provincia de La Pampa: Santa Rosa, Argentina, 1974; p. 126. [Google Scholar]
- Corradini, E.; Zilocchi, H.; Cuesta, R.; Segesso, R.; Jiménez, M.L.; Musco, J.M. Caracterización del Sector Productor Tabacalero en la República Argentina, 3rd ed.; Serie “Documentos de Investigación”; Universidad Catolica Argentina Santa Maria de Los Buenos Aires, Faculdad de Ciencias Agrarias Centro de Altos Estudios “Jorge Gándara”: Buenos Aires, Argentina, 2005; p. 94. Available online: https://www.magyp.gob.ar/sitio/areas/tabaco/informes/publicaciones/_archivos/000002-Estudios/000002-Caracterizaci%C3%B3n%20del%20Sector%20Tabacalero%20Argentino/000001-Informe%203%C2%BA%20Versi%C3%B3n%20-%20Junio%202005.pdf (accessed on 18 September 2019).
- Bennett, A.; du Toit, C.L.N.; Bennett, A.L. A new record of Lema trilinea White (Coleoptera: Chrysomelidae) on tobacco in South Africa, with reference to the common pest species, Lema bilineata (Germar) (Coleoptera: Chrysomelidae, Criocerinae). Afr. Entomol. 1999, 7, 316–319. Available online: https://journals.co.za/content/ento/7/2/AJA10213589_377 (accessed on 30 November 2019).
- Araya, J.E.; Ormeño, J.; Díaz, C.A. Calidad hospedera de Datura spp. y otras solanáceas para Lema bilineata Germar. Bol. San. Veg. Plagas 2000, 26, 65–71. Available online: https://www.miteco.gob.es/ministerio/pags/Biblioteca/Revistas/pdf_plagas/BSVP-26-01-065-071.pdf (accessed on 28 July 2019).
- Bado, S.G.; Mareggiani, G.; Vilella, F. Preferencia alimentaria de Lema bilineata G. (Coleoptera: Chrysomelidae) hacia tres Solanaceae (Solanoideae): Posibles defensas químicas. Bol. San. Veg. Plagas 2001, 27, 169–175. Available online: https://www.miteco.gob.es/ministerio/pags/Biblioteca/Revistas/pdf_plagas/BSVP-27-02-169-175.pdf (accessed on 28 July 2019).
- Díaz, C.A.; Ormeño, J.; Araya, J.E. Especificidad de Lema bifineata G. hacia Datura spp. mediante centrifugación filogenética. Bol. San. Veg. Plagas 1999, 25, 259–264. Available online: https://www.miteco.gob.es/ministerio/pags/Biblioteca/Revistas/pdf_plagas%2FBSVP-25-03-259-264.pdf (accessed on 23 July 2019).
- Ormeño, J.; Araya, J.E.; Díaz, C.A. Supervivencia y alimentación comparativa de larvas de Lema bilineata Germar en Datura spp., tabaco y otras solanáceas. Bol. San. Veg. Plagas 2002, 28, 21–31. Available online: https://www.miteco.gob.es/ministerio/pags/biblioteca/plagas/BSVP-28-01-021-031.pdf (accessed on 28 July 2019).
- Heinze, E.; Pinsdorf, W. Die Criocerinen Afrikas (Col. Chrysomelidae). Entomol. Arb. Mus. G. Frey 1964, 15, 334–569. [Google Scholar]
- Bado, S.G.; Cerri, A.M.; Vilella, F.; Molina, A. Aspectos morfológicos, biológicos y de ingesta de Lema bilineata (Germ) (Coleoptera: Chrysomelidae) sobre Physalis peruviana L. Bol. San. Veg. Plagas 2000, 26, 5–10. Available online: https://www.miteco.gob.es/ministerio/pags/biblioteca/plagas/BSVP-26-01-005-010.pdf (accessed on 28 July 2019).
- Omer-Cooper, J.; Miles, P. On Lema trilineata: A beetle closely resembling the tobacco slug, attacking the Cape gooseberry. S. Afr. J. Sci. 1951, 47, 330–333. Available online: https://journals.co.za/content/sajsci/47/12/AJA00382353_3382 (accessed on 24 November 2019).
- Franzmann, B.A. Discovery of Lema trilineata (Olivier) in Queensland. Entomol. Soc. Qld. News Bull. 1978, 6, 119–120. [Google Scholar]
- Vencl, F.V.; Levy, A.; Geeta, R.; Keller, G.; Windsor, D.M. Observations on the natural history, systematics and phylogeny of the Criocerinae of Costa Rica and Panama. In New Developments in the Biology of Chrysomelidae; Jolivet, P., Santiago-Blay, J.A., Schmitt, M., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 2004; pp. 423–454. [Google Scholar]
- Warchałowski, A. An introductory review of Lema Fabr. species from Eastern and Southeastern Asia (Coleoptera: Chrysomelidae: Criocerinae). Genus 2011, 22, 29–93. [Google Scholar]
- White, R.E. A revision of the subfamily Criocerinae (Chrysomelidae) of North America north of Mexico. U.S. Dep. Agric. Tech. Bull. 1993, 1805, 1–158. [Google Scholar]
- Matsumura, Y.; Yoshizawa, K. Homology of the internal sac components in the leaf beetle subfamily Criocerinae and evolutionary novelties related to the extremely elongated flagellum. J. Morphol. 2012, 273, 507–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, Y.; Yao, I.; Beutel, R.G.; Yoshizawa, K. Molecular phylogeny of the leaf beetle subfamily Criocerinae (Coleoptera: Chrysomelidae) and the correlated evolution of reproductive organs. Arthropod Syst. Phylo. 2014, 72, 95–110. Available online: https://www.senckenberg.de/wp-content/uploads/2019/08/02_asp_72_2_matsumura_et_al_95-110.pdf (accessed on 24 November 2019).
- Gebiola, M.; Monti, M.M.; Johnson, R.C.; Woolley, J.B.; Hunter, M.S.; Giorgini, M.; Pedata, P.A. A revision of the Encarsia pergandiella species complex (Hymenoptera: Aphelinidae) shows cryptic diversity in parasitoids of whitefly pests. Syst. Entomol. 2017, 42, 31–59. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- Hajibabaei, M.; Janzen, D.H.; Burns, J.M.; Hallwachs, W.; Hebert, P.D. DNA barcodes distinguish species of tropical Lepidoptera. Proc. Natl. Acad. Sci. USA 2006, 103, 968–971. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acid. S. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.-X.; Hewitt, G.M. Nuclear integrations: Challenges for mitochondrial DNA markers. Trends Ecol. Evol. 1996, 11, 247–251. [Google Scholar] [CrossRef]
- Bensasson, D.; Zhang, D.X.; Hartl, D.L.; Hewitt, G.M. Mitochondrial pseudogenes: Evolution’s misplaced witnesses. Trends Ecol. Evol. 2001, 16, 314–321. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Lanave, C.; Preparata, G.; Saccone, C.; Serio, G. A new method for calculating evolutionary substitution rates. J. Mol. Evol. 1984, 20, 86–93. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Magoga, G.; Sahin, D.C.; Fontaneto, D.; Montagna, M. Barcoding of Chrysomelidae of Euro-Mediterranean area: Efficiency and problematic species. Sci. Rep. 2018, 8, 13398. [Google Scholar] [CrossRef]
- White, R.E. Homonymy in world species-group names of Criocerinae (Coleoptera: Chrysomelidae). U.S. Dep. Agric. Tech. Bull. 1981, 1629, 1–69. [Google Scholar]
- Riley, E.G.; Clark, S.M.; Seeno, T.N. Catalog of the Leaf Beetles of America North of Mexico (Coleoptera: Megalopodidae, Orsodacnidae and Chrysomelidae, Excluding Bruchinae); Special Publication no. 1; Coleopterists Society: Sacramento, CA, USA, 2003; p. 187. [Google Scholar]
- Clark, S.M.; LeDoux, D.G.; Seeno, T.N.; Riley, E.G.; Gilbert, A.J.; Sullivan, J.M. Host plants of leaf beetle species occurring in the United States and Canada (Coleoptera: Megalopodidae, Orsodacnidae, Chrysomelidae exclusive of Bruchinae); Special Publication No. 2; Coleopterists Society: Sacramento, CA, USA, 2004; p. 146. Available online: https://www.zin.ru/animalia/coleoptera/pdf/clark_ledoux_et_al_2004.pdf (accessed on 18 September 2019).
- Aoyagi, M. Lema trivittata taken from Miyakojima, I., a new record from Japan. Gekkan-Mushi 2012, 497, 27–28. [Google Scholar]
- Kawaji, K. Lema trivittata was collected from Iriomotejima. Gekkan-Mushi 2012, 502, 13–14. [Google Scholar]
- Lee, C.-F.; Matsumura, Y. On newly and recently recorded species of the genus Lema Fabricius (Coleoptera, Chrysomelidae, Criocerinae) from Taiwan. ZooKeys 2013, 262, 17–38. [Google Scholar] [CrossRef] [Green Version]
- Flowers, R.W.; Eberhard, W.G. Fitting together: Copulatory linking in some Neotropical Chrysomeloidea. Rev. Biol. Trop. 2006, 54, 829–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Düngelhoef, S.; Schmitt, M. Functional morphology of copulation in Chrysomelidae-Criocerinae and Bruchidae (Insecta: Coleoptera). Bonn. Zool. Beitr. 2005, 54, 201–208. [Google Scholar]
- Suzuki, K. Comparative morphology of the internal reproductive system of the Chrysomelidae (Coleoptera). In Biology of Chrysomelidae; Series Entomologica; Jolivet, P., Petitpierre, E., Hsiao, T.H., Eds.; Springer: Dordrecht, The Netherlands, 1988; Volume 42, pp. 317–355. [Google Scholar]
- Matsumura, Y.; Suzuki, K. Comparative morphology of internal reproductive systems in leaf beetles of the Donaciinae and Criocerinae (Coleoptera: Chrysomelidae) and its implication for the phylogeny. Zootaxa 2008, 1845, 1–32. [Google Scholar] [CrossRef]
- Silveira-Guido, A.; Carbonell Bruhn, J. Los insectos enemigos del girasol en el Uruguay. Boletín Fac. Agron. Univ. Montev. 1965, 81, 5–64. [Google Scholar]
- Quintana, F.J.; Abot, A.R. Girasol: Lista Comentada de Los Organismos Animales Que Atacan al Cultivo en la República Argentina; INTA, Facultad de Ciencias Agrarias de la Universidad Nacional de Mar del Plata: Balcarce, Argentina, 1987; p. 30. [Google Scholar]
- Saini, E.D. Insectos y Ácaros Perjudiciales al Cultivo del Girasol y Sus Enemigos Naturales; Publicación del Instituto de Microbiología y Zoología Agrícola N° 8. INTA: Buenos Aires, Argentina, 2004; pp. 20–21. [Google Scholar]
- Speyer, W. Chrysomelidae. In Handbuch der Pflanzenkrankheiten; Blunck, H., Ed.; Paul Parey: Berlin/Heidelberg, Germany, 1954; pp. 270–299. [Google Scholar]
- Beenen, R.; Roques, A. Leaf and seed beetles (Coleoptera, Chrysomelidae). BioRisk 2010, 4, 267–292. [Google Scholar] [CrossRef]
- Boriani, M.; Calvi, M.; Taddei, A.; Tantardini, A.; Cavagna, B.; Spadoni Andreani, F.; Montagna, M.; Bonini, M.; Lommen, S.; Müller-Schäre, H. Ophraella communa segnalata in Italia su Ambrosia. L’Informatore Agrario 2013, 69, 61. Available online: http://www.ilpolline.it/wp-content/uploads/2013/09/Ophraella-communa.pdf (accessed on 30 April 2020).
- Clark, S.M.; Rattu, A.; Cillo, D. Monoxia obesula Blake, 1939, a species native to the U.S.A. and adventive to Sardinia, Italy (Coleoptera: Chrysomelidae: Galerucinae: Galerucini). Zootaxa 2014, 3774, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagna, M.; Zoia, S.; Leonardi, C.; Di Taddeo, V.; Caldara, R.; Sassi, D. Colasposoma dauricum Mannerheim, 1849 an Asian species adventive to Piedmont, Italy (Coleoptera: Chrysomelidae: Eumolpinae). Zootaxa 2016, 4097, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Malumphy, C. Arthropods intercepted on fresh pineapple fruit, Ananas comosus (L.) Merr. imported into England and Wales. Entomol. Gaz. 2015, 66, 65–75. [Google Scholar]
- Schmitt, M. The Criocerinae: Biology, Phylogeny and Evolution. In Biology of Chrysomelidae; Series Entomologica; Jolivet, P., Petitpierre, E., Hsiao, T.H., Eds.; Springer: Dordrecht, The Netherlands, 1988; Volume 42, pp. 475–495. [Google Scholar]
- EASIN. European Alien Species Information Network; European Commission–Joint Research Centre: Ispra, Italy, 2019; Available online: https://easin.jrc.ec.europa.eu/ (accessed on 15 November 2019).


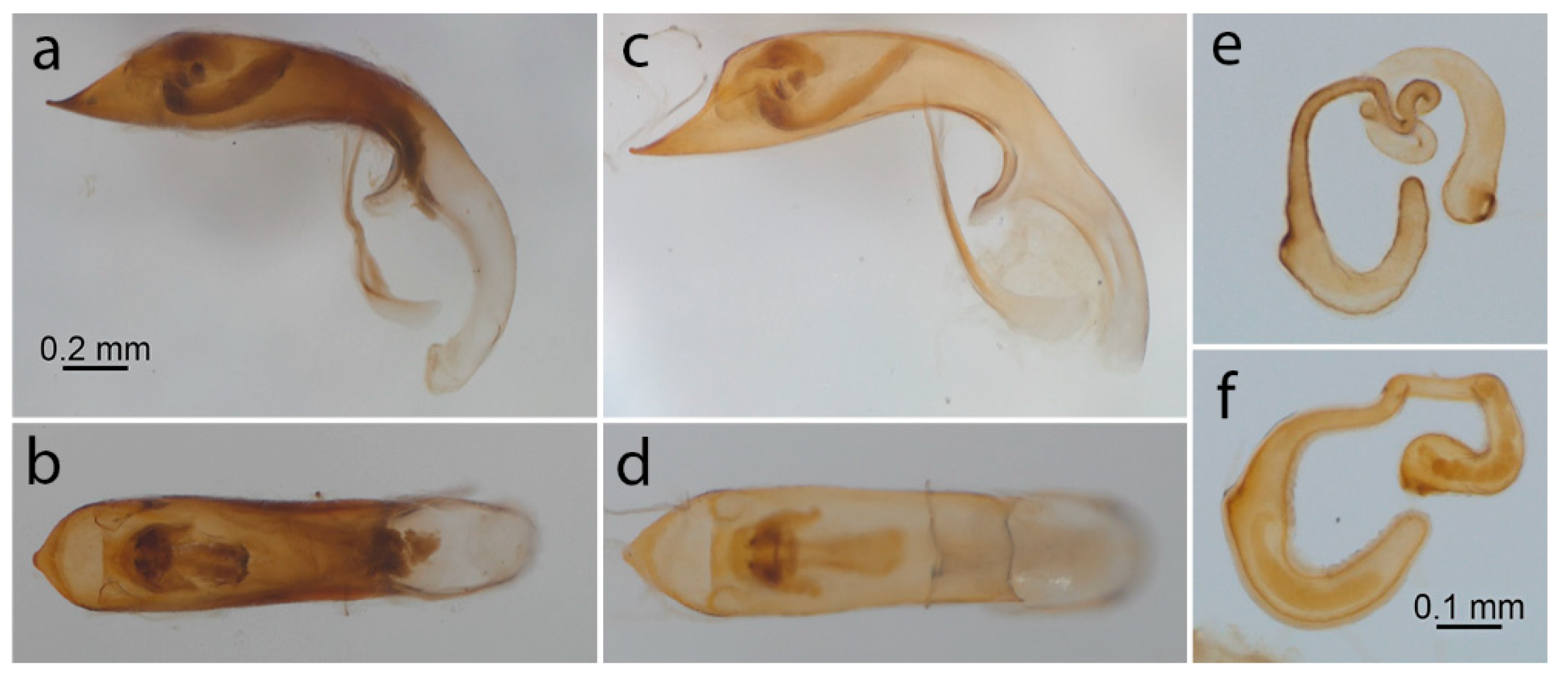


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Voucher | Lema Species | Collection Site | Coordinates | Date | Storage | Host |
|---|---|---|---|---|---|---|
| SANC MCOL 0005 | L. bilineata | SA: (Western Cape) Wilderness Ntl Park 10 km ESE George | 34°00′ S 22°34′ E | 20.xi.2000 | frozen | not available |
| SANC MCOL 0475 | L. bilineata | SA: (North West) Rustenburg | 25°40′ S 27°15′ E | i.1996 | dry | Datura sp. |
| SANC MCOL 0477 | L. bilineata | SA: (Gauteng) Montana, Pretoria | 25°40′ S 28°15′ E | 18.i.1994 | dry | Physalis peruviana |
| SANC MCOL 507-1 | L. bilineata | SA: (Gauteng) Roodeplaat | 25°36′ S 28°22′ E | 09.xi.2018 | EtOH | Datura stramonium |
| SANC MCOL 507-2 | L. bilineata | SA: (Gauteng) Roodeplaat | 25°36′ S 28°22′ E | 09. xi.2018 | EtOH | Datura stramonium |
| SANC MCOL 507-3 | L. bilineata | SA: (Gauteng) Roodeplaat | 25°36′ S 28°22′ E | 09. xi.2018 | EtOH | Datura stramonium |
| SANC MCOL 508-1 | L. daturaphila | SA: (Gauteng) Roodeplaat | 25°35′ S 28°20′ E | 29.x.2018 | EtOH | Datura stramonium |
| SANC MCOL 508-2 | L. daturaphila | SA: (Gauteng) Roodeplaat | 25°35′ S 28°20′ E | 29.x.2018 | EtOH | Datura stramonium |
| SANC MCOL 508-3 | L. daturaphila | SA: (Gauteng) Roodeplaat | 25°35′ S 28°20′ E | 29.x.2018 | EtOH | Datura stramonium |
| SANC MCOL 509-1 | L. bilineata | SA: (Gauteng) Roodeplaat | 25°36′ S 28°22′ E | 05. xi.2018 | EtOH | Datura stramonium |
| SANC MCOL 509-2 | L. bilineata | SA: (Gauteng) Roodeplaat | 25°36′ S 28°22′ E | 05. xi.2018 | EtOH | Datura stramonium |
| SANC MCOL 510-1 | L. bilineata | SA: (Gauteng) Roodeplaat | 25°36′ S 28°22′ E | 05. xi.2018 | EtOH | Datura stramonium |
| SANC MCOL 510-2 | L. bilineata | SA: (Gauteng) Roodeplaat | 25°36′ S 28°22′ E | 05. xi.2018 | EtOH | Datura stramonium |
| IPSP 1 | L. bilineata | I: (Campania) Capaccio | 40°27′ N 15°03′ E | 27.vii.2018 | fresh | Nicotiana tabacum |
| IPSP 2 | L. bilineata | I: (Campania) Caserta | 41°04′ N 14°20′ E | 23.vi.2018 | fresh | Salpichroa origanifolia |
| IPSP 3 | L. bilineata | I: (Campania) Acerra | 40°57′ N 14°20′ E | 26.vi.2018 | fresh | Nicotiana tabacum |
| IPSP 4 | L. bilineata | I: (Campania) Cava de’ Tirreni | 40°44′ N 14°42′ E | 18.vii.2018 | fresh | Nicotiana tabacum |
| IPSP 5 | L. bilineata | I: (Campania) Montesarchio | 41°02′ N 14°38′ E | 01.viii.2018 | fresh | Nicotiana tabacum |
| IPSP 6 | L. bilineata | I: (Campania) Napoli | 40°50′ N 14°14′ E | 25.vii.2017 | fresh | Physalis peruviana |
| IPSP 7 | L. bilineata | I: (Campania) Portici | 40°48′ N 14°21′ E | 02.viii.2017 | fresh | Salpichroa origanifolia |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monti, M.M.; Ruocco, M.; Grobbelaar, E.; Pedata, P.A. Morphological and Molecular Characterization of Lema bilineata (Germar), a New Alien Invasive Leaf Beetle for Europe, with Notes on the Related Species Lema daturaphila Kogan & Goeden. Insects 2020, 11, 295. https://doi.org/10.3390/insects11050295
Monti MM, Ruocco M, Grobbelaar E, Pedata PA. Morphological and Molecular Characterization of Lema bilineata (Germar), a New Alien Invasive Leaf Beetle for Europe, with Notes on the Related Species Lema daturaphila Kogan & Goeden. Insects. 2020; 11(5):295. https://doi.org/10.3390/insects11050295
Chicago/Turabian StyleMonti, Maurilia M., Michelina Ruocco, Elizabeth Grobbelaar, and Paolo A. Pedata. 2020. "Morphological and Molecular Characterization of Lema bilineata (Germar), a New Alien Invasive Leaf Beetle for Europe, with Notes on the Related Species Lema daturaphila Kogan & Goeden" Insects 11, no. 5: 295. https://doi.org/10.3390/insects11050295
APA StyleMonti, M. M., Ruocco, M., Grobbelaar, E., & Pedata, P. A. (2020). Morphological and Molecular Characterization of Lema bilineata (Germar), a New Alien Invasive Leaf Beetle for Europe, with Notes on the Related Species Lema daturaphila Kogan & Goeden. Insects, 11(5), 295. https://doi.org/10.3390/insects11050295