Potential Distribution and the Risks of Bactericera cockerelli and Its Associated Plant Pathogen Candidatus Liberibacter Solanacearum for Global Potato Production


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
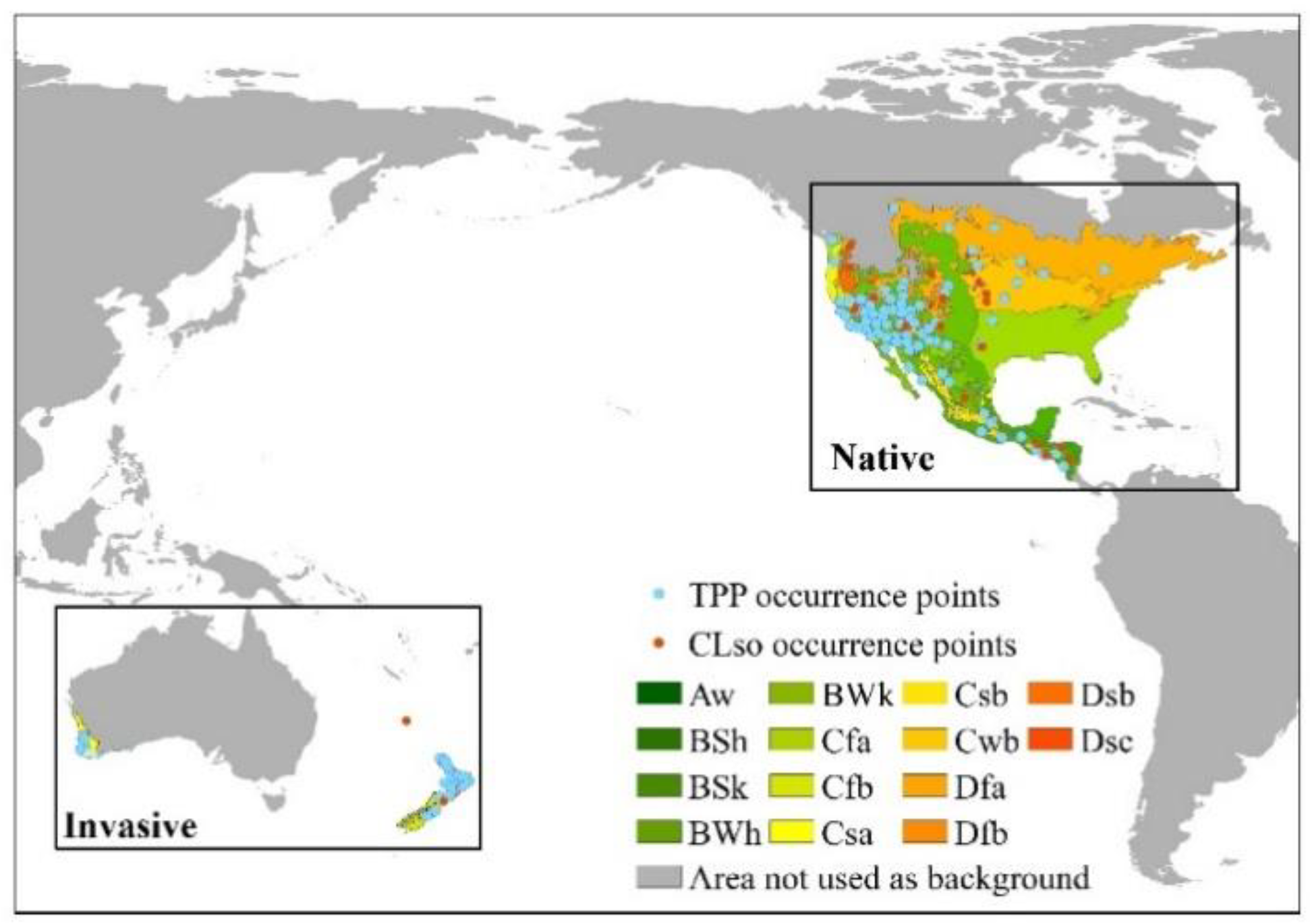
2.1. Occurrence Data Collection and Spatial Filtering
2.2. Background Study Area Delimitation
2.3. Occupied Climate Space Comparison between Native and Invasive Populations
2.4. Climate Variable Selection
2.5. MaxEnt Parameter Configuration and Best Model Selection
2.6. Model Projection to Predict the Potential Distribution of TPP and CLso
2.7. Spatial Analyses for Quantifying the Area at Risk of Attack
3. Results
3.1. Occupied Climate Space Comparison between Native and Introduced Populations
3.2. Model Calibration and Evaluation
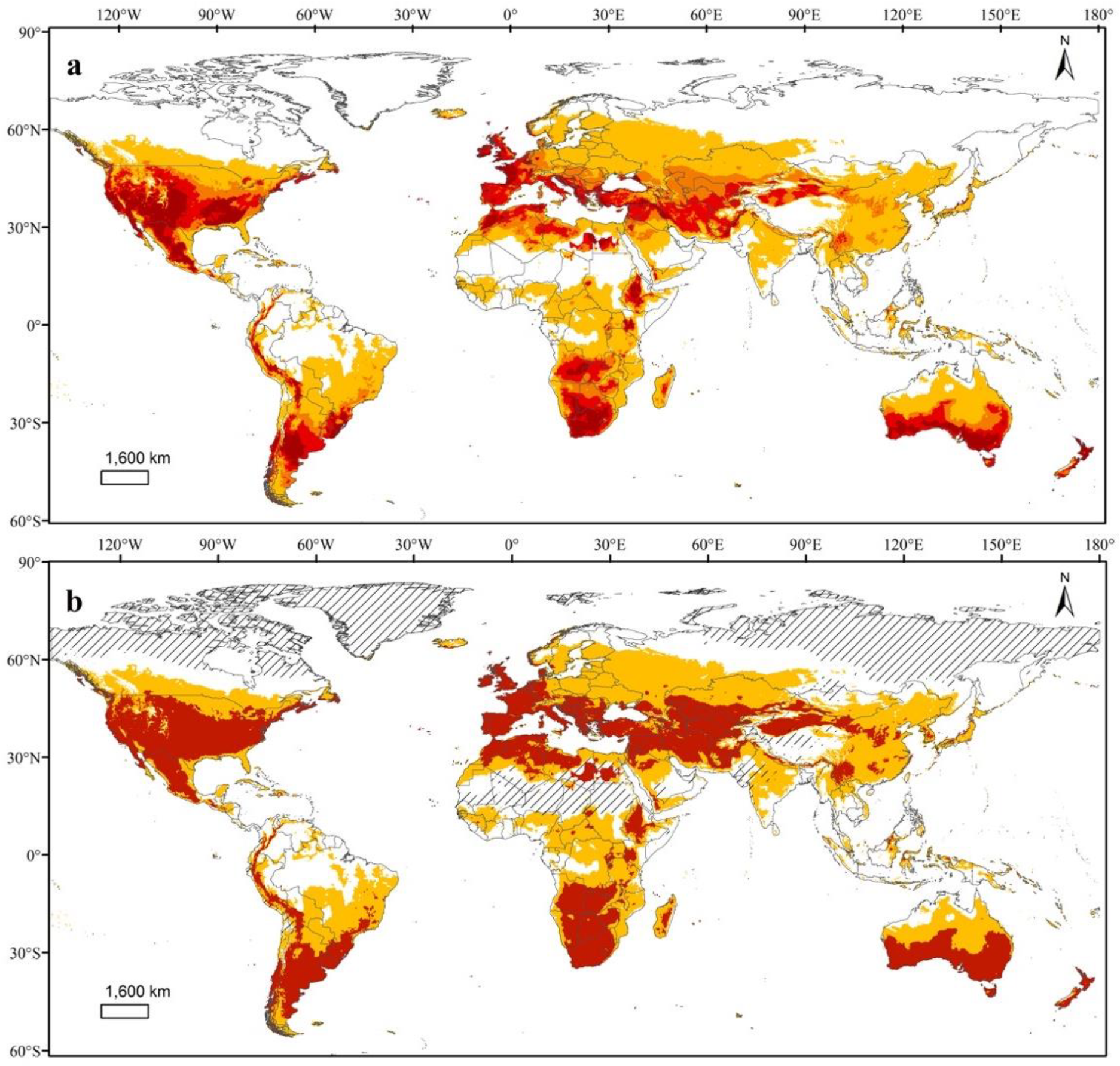
3.3. Potential Global Distributions of TPP and the Bacterial Pathogen It Transmits
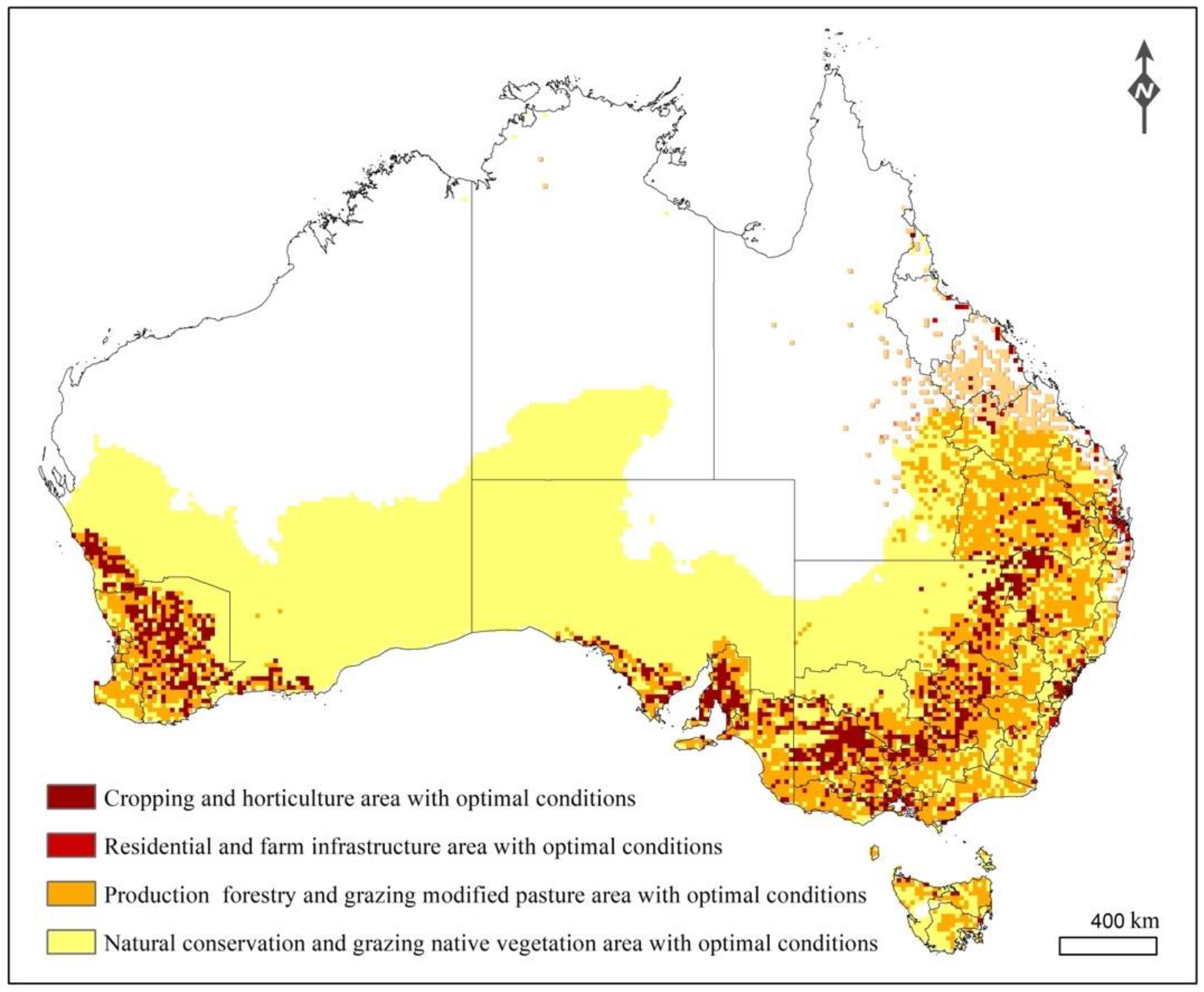
3.4. Risks to Global Potato Production and Australian Crop Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hulme, P.E.; Pyšek, P.; Nentwig, W.; Vilà, M. Will threat of biological invasions unite the European Union? Science 2009, 324, 40–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Wajnberg, E.; Wyckluys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Ruescas, D.C.; Tabone, E.; Frandon, J.; et al. Biological invasion of European tomato crops by Tuta absoluta ecology, geography expansion and prospects for biological control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. PNAS 2016, 113, 7575–7579. [Google Scholar] [CrossRef] [Green Version]
- Bacon, S.J.; Aebi, A.; Calanca, P.; Bacher, S. Quarantine arthropod invasions in Europe: The role of climate, hosts and propagule pressure. Divers Distrib. 2014, 20, 84–94. [Google Scholar] [CrossRef]
- Teulon, D.A.J.; Workman, P.J.; Thomas, K.L.; Nielsen, M.C. Bactericera cockerelli: Incursion, dispersal and current distribution on vegetable crops in New Zealand. NZPP 2009, 62, 136–144. [Google Scholar] [CrossRef] [Green Version]
- McNeill, M.R.; Phillips, C.B.; Bell, N.L.; Proffitt, J.R. Potential spread of pests in New Zealand through commercial transport of nursery plants. NZPP 2006, 59, 75–79. [Google Scholar] [CrossRef]
- Munyaneza, J.E. Zebra chip disease of potato: Biology, epidemiology, and management. Am. J. Potato Res. 2012, 89, 329–350. [Google Scholar] [CrossRef] [Green Version]
- Syfert, M.M.; Serbina, L.; Burckhardt, D.; Knapp, S.; Percy, D.M. Emerging New Crop Pests: Ecological Modelling and Analysis of the South American Potato Psyllid Russelliana solanicola (Hemiptera: Psylloidea) and Its Wild Relatives. PLoS ONE 2017, 12, e0167764. [Google Scholar] [CrossRef] [Green Version]
- EPPO (European and Mediterranean Plant Protection Organization). Bactericera cockerelli. EPPO Bull. 2013, 43, 202–208. [Google Scholar] [CrossRef]
- Salazar, L.F. Emerging and re-emerging potato diseases in the Andes. Potato Res. 2006, 49, 43–47. [Google Scholar] [CrossRef]
- Weintraub, P.G.; Beanland, L. Insect vectors of phytoplasmas. Annu. Rev. Entomol. 2006, 51, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, X.; Hoffmann, M.; Coy, M.R.; Stelinski, L.L.; Pelz-Stelinski, K.S. Infection of an insect vector with a bacterial plant pathogen increases its propensity for dispersal. PLoS ONE 2015, 10, e0129373. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.L.; Jones, D.C.; Kumarasinghe, L.B.; Richmond, J.E.; Gill, G.S.C.; Bullians, M.S. Investigation into the entry pathway for the tomato potato psyllid Bactericera cockerelli. NZPP 2011, 64, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Vereijssen, J.; Smith, G.R.; Weintraub, P.G. Bactericera cockerelli (Hemiptera: Triozidae) and Candidatus Liberibacter solanacearum in Potatoes in New Zealand: Biology, Transmission, and Implications for Management. J. Integr. Pest Manag. 2018, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Vereijssen, J. Ecology and management of Bactericera cockerelli and Candidatus Liberibacter solanacearum in New Zealand. J. Integr. Agr. 2020, 19, 333–337. [Google Scholar] [CrossRef]
- Gill, G. Tomato psyllid detected in New Zealand. Biosecurity 2006, 69, 10–11. [Google Scholar]
- Venette, R.C.; Kriticos, D.J.; Magarey, R.D.; Koch, F.H.; Baker, R.H.A.; Worner, S.P.; Raboteaux, N.N.G.; McKenney, D.W.; Dobesberger, E.J.; Yemshanov, D.; et al. Pest risk maps for invasive alien species: A roadmap for improvement. BioScie 2010, 60, 349–362. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberon, J.; Pearson, R.G.; Anderson, R.P.; Martinez-Meyer, E.; Nakamura, M.; Araujo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Cambridge, UK, 2011; p. 314. [Google Scholar]
- Gutierrez Illan, J.; Bloom, E.H.; Wohleb, C.H.; Wenninger, E.J.; Rondon, S.I.; Jensen, A.S.; Snyder, W.E.; Crowder, D.W. Landscape structure and climate drive population dynamics of an insect vector within intensely managed agroecosystems. Ecol. Appl. 2020. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Marchioro, C.A. Global Potential Distribution of Bactrocera carambolae and the risks for fruit production in Brazil. PLoS ONE 2016, 11, e0166142. [Google Scholar] [CrossRef] [PubMed]
- Estay, S.A.; Labra, F.A.; Sepulveda, R.D.; Bacigalupe, L.D. Evaluating habitat suitability for the establishment of Monochamus spp. through climate-based niche modeling. PLoS ONE 2014, 9, e102592. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modelling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1–12. [Google Scholar] [CrossRef]
- DAFWA. Tomato potato psyllid. Department of Western Australia. 2017. [Google Scholar]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Method Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudik, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M. Modelling of species distribution with Maxent: New extensions and a com- prehensive evaluation. Ecography 2008, 21, 161–175. [Google Scholar] [CrossRef]
- Rutherford, M.C. Vegetation of Southern Africa. Categorization of Biomes; Cowling, R.M., Richardson, D.M., Pierce, S.M., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 91–98. [Google Scholar]
- Webber, B.; Yates, C.; Le Maitre, D.C.; Scott, J.K.; Kriticos, D.J.; Ota, N.; McNeill, A.; Le Roux, J.J.; Midgley, G.F. Modelling horses for novel climate courses: Insights from projecting potential distributions of native and alien Australian acacias with correlative and mechanistic models. Divers Distrib. 2011, 17, 978–1000. [Google Scholar] [CrossRef]
- Hill, M.P.; Terblanche, J.S. Niche overlap of congeneric invaders supports a single-species hypothesis and provides insight into future invasion risk: Implications for global management of the Bactrocera dorsalis complex. PLoS ONE 2014, 9, e90121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broennimann, O.; Treier, U.A.; Müller-Schärer, H.; Thuiller, W.; Peterson, A.T.; Guisan, A. Evidence of climatic niche shift during biological invasion. Biol. Lett. 2007, 10, 701–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broennimann, O.; Guisan, A. Predicting current and future biological invasions: Both native and invaded ranges matter. Biol. Lett. 2008, 4, 585–589. [Google Scholar] [CrossRef]
- Tingley, R.; Vallinoto, M.; Sequeira, F.; Kearney, M.R. Realized niche shift during a global biological invasion. Proc. Natl. Acad. Sci. USA 2014, 111, 10233–10238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models. With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Slater, H.; Michael, E. Predicting the current and future potential distributions of lymphatic filariasis in Africa using maximum entropy ecological niche modelling. PLoS ONE 2012, 7, e32202. [Google Scholar] [CrossRef]
- Sheppard, C.S. How does selection of climate variables affect predictions of species distributions? A case study of three new weeds in New Zealand. Weed Res. 2013, 53, 259–268. [Google Scholar] [CrossRef]
- Barve, N.; Barve, V.; Jimenéz-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Kumar, S.; Neven, L.G.; Zhu, H.; Zhang, R. Assessing the global risk of establishment of Cydia pomonella (Lepidoptera: Tortricidae) using CLIMEX and MaxEnt niche models. J. Econ. Entomol. 2015, 108, 1708–1719. [Google Scholar] [CrossRef] [PubMed]
- Morales, N.S.; Fernández, I.C.; Baca⁃González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. PeerJ 2017, 5, e3093. [Google Scholar] [CrossRef] [PubMed]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distribution: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Method Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Owens, H.L.; Campbell, L.P.; Dornak, L.L.; Saupe, E.E.; Barve, N.; Soberon, J.; Ingenloff, K.; Lira-Noriega, A.; Hensz, C.M.; Myers, C.E. Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecol. Model. 2013, 263, 10–18. [Google Scholar] [CrossRef]
- Fitzpatrick, M.C.; Hargrove, W.H. The projection of species distribution models and the problem of non-analog climate. Biodivers Conserv. 2009, 18, 2255–2261. [Google Scholar] [CrossRef]
- Schafleitner, R.; Ramirez, J.; Jarvis, A.; Evers, D.; Gutierrez, R.; Scurrah, M. Adaptation of the potato crop to changing climates. In Crop Adaptation to Climate change; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; Wiley-Blackwell: Chichester, UK, 2011; pp. 287–297. [Google Scholar]
- Hijmans, R.J. Global distribution of the potato crop. Am. J. Potato Res. 2001, 78, 403–412. [Google Scholar] [CrossRef]
- Warren, D.L.; Wright, A.N.; Seifert, S.N.; Shaffer, H.B. Incorporating model complexity and spatial sampling bias into ecological niche models of climate change risks faced by 90 California vertebrate species of concern. Divers Distrib. 2014, 20, 334–343. [Google Scholar] [CrossRef]
- Moreno-Amat, E.; Mateo, R.G.; Nieto-Lugilde, D.; Marueta-Holme, N.; Svenning, J.C.; García-Amorena, I. Impact of model complexity on cross-temporal transferability in MaxEnt species distribution models: An assessment using paleobotanical data. Ecol. Model. 2015, 312, 308–317. [Google Scholar] [CrossRef]
- Lodge, D.M.; Williams, S.; MacIsaac, H.J.; Hayes, K.R.; Leung, B.; Reichard, S.; Mack, R.N.; Moyle, P.B.; Smith, M.; Andow, D.A.; et al. Biological invasions: Recommendations for U.S. policy and management. Ecol. Appl. 2006, 16, 2035–2054. [Google Scholar] [CrossRef] [Green Version]
- Wan, F.H.; Yang, N.W. Invasion and management of agricultural alien insects in China. Annu. Rev. Entomoly 2016, 61, 77–98. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, J.; Wang, R.; Ren, Y.; McKirdy, S. Potential Distribution and the Risks of Bactericera cockerelli and Its Associated Plant Pathogen Candidatus Liberibacter Solanacearum for Global Potato Production. Insects 2020, 11, 298. https://doi.org/10.3390/insects11050298
Wan J, Wang R, Ren Y, McKirdy S. Potential Distribution and the Risks of Bactericera cockerelli and Its Associated Plant Pathogen Candidatus Liberibacter Solanacearum for Global Potato Production. Insects. 2020; 11(5):298. https://doi.org/10.3390/insects11050298
Chicago/Turabian StyleWan, Jing, Rui Wang, Yonglin Ren, and Simon McKirdy. 2020. "Potential Distribution and the Risks of Bactericera cockerelli and Its Associated Plant Pathogen Candidatus Liberibacter Solanacearum for Global Potato Production" Insects 11, no. 5: 298. https://doi.org/10.3390/insects11050298
APA StyleWan, J., Wang, R., Ren, Y., & McKirdy, S. (2020). Potential Distribution and the Risks of Bactericera cockerelli and Its Associated Plant Pathogen Candidatus Liberibacter Solanacearum for Global Potato Production. Insects, 11(5), 298. https://doi.org/10.3390/insects11050298