Phylogeny of Anopheles (Kerteszia) (Diptera: Culicidae) Using Mitochondrial Genes

 ,
,  ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
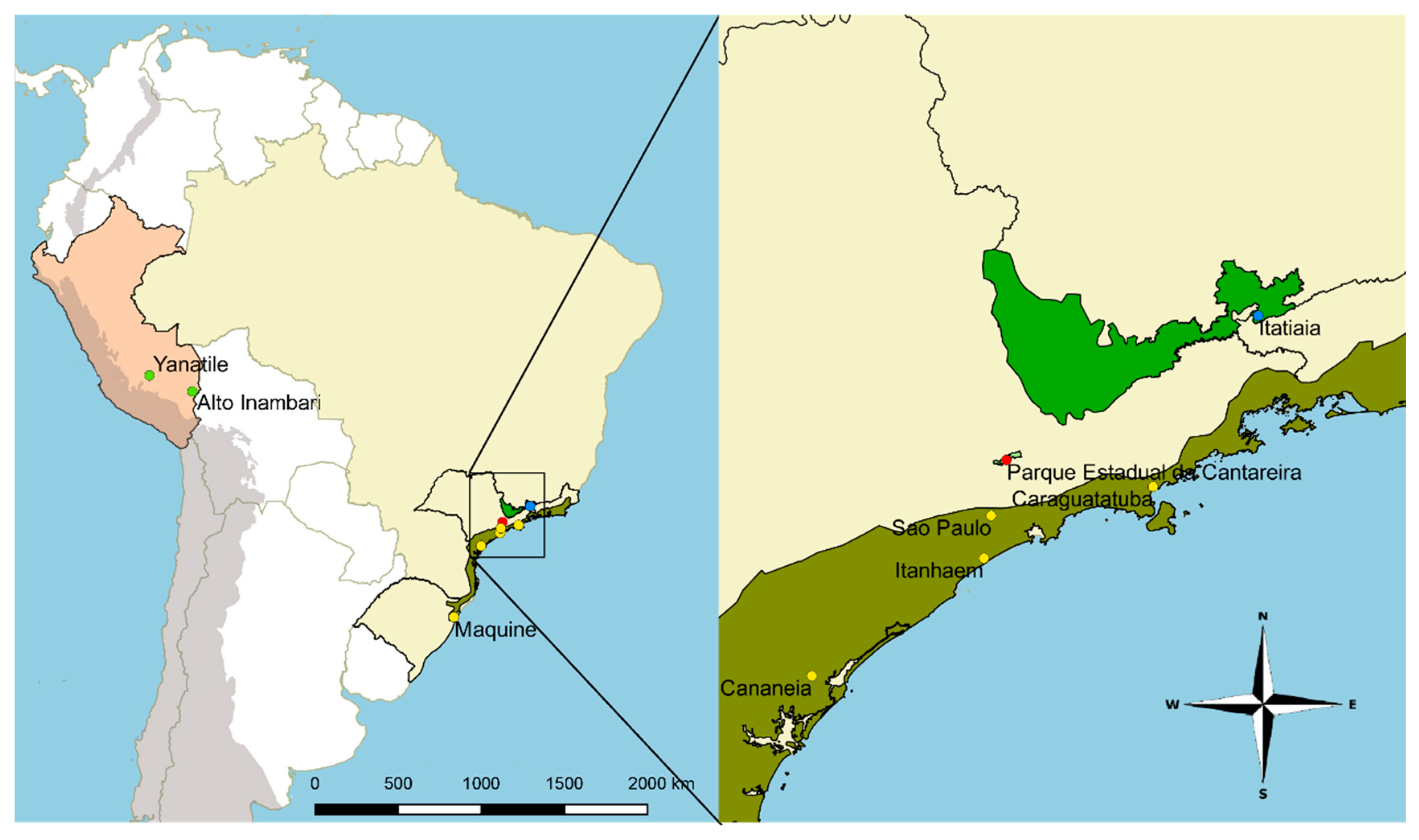
2.1. Mosquito Sampling and Handling
2.2. Genomic DNA Extraction and PCR Amplification of Mitochondrial Gene Fragments
2.3. Sequencing, Alignment, and Phylogenetic Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Centro Nacional de Epidemiología, Prevención y Control de Enfermedades—MINSA, Perú. Available online: http://www.dge.gob.pe/portal/docs/vigilancia/sala/2017/SE51/malaria.pdf (accessed on 22 May 2019).
- Pan American Health Organization; World Health Organization. Epidemiological Update: Malaria in the Americas; PAHO: Washington, DC, USA; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Recht, J.; Siqueira, A.M.; Monteiro, W.M.; Herrera, S.M.; Herrera, S.; Lacerda, M.V.G. Malaria in Brazil, Colombia, Peru and Venezuela: Current challenges in malaria control and elimination. Malar. J. 2017, 16, 273. [Google Scholar] [CrossRef]
- Sulzer, A.J.; Cantella, R.; Colichon, A.; Gleason, N.N.; Walls, K.W. A focus of hyperendemic Plasmodium malariae–P. vivax with no P. falciparum in a primitive population in the Peruvian Amazon jungle. Bull. World Health Organ. 1975, 52, 273–278. [Google Scholar]
- Aramburu, G.J.; Asayag, C.R.; Witzig, R. Malaria reemergence in the peruvian amazon region. Emerg. Infect. Dis. 1999, 5, 209–215. [Google Scholar]
- Manguin, S.R.; Wilkerson, R.C.; Conn, J.E.; Rubio-Palis, Y.; Danoff-Burg, J.A.; Roberts, D.R. Population structure of the primary malaria vector in South America, Anopheles darlingi, using isoenzyme, ITS2, RAPD, and morphological markers. Am. J. Trop. Med. Hyg. 1999, 60, 364–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, C.; Fernández, R.; Escobedo, K.S.; Vela, Q.; Schoeler, G.B. Natural Plasmodium infections in Anopheles darlingi and Anopheles benarrochi (Diptera: Culicidae) from Eastern Peru. J. Med. Entomol. 2004, 41, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Reinbold-Wasson, D.D.; Sardelis, M.R.; Jones, J.W.; Watts, D.M.; Fernandez, R.; Carbajal, F.; Pecor, J.E.; Calampa, C.; Klein, T.A.; Turell, M.J. Determinants of Anopheles seasonal distribution patterns across a forest to periurban gradient near Iquitos, Peru. Am. J. Trop. Med. Hyg. 2012, 86, 459–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, M.; Saavedra, M.P.; Bickersmith, S.A.; Prussing, C.; Michalski, A.; Tong Rios, C.; Vinetz, J.M.; Conn, J.E. Intensive trapping of blood-fed Anopheles darlingi in Amazonian Peru reveals unexpectedly high proportions of avian blood-meals. PLoS Negl. Trop. Dis. 2017, 11, e0005337. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Ferreira, J.; Lacerda, M.V.; Brasil, P.; Ladislau, J.L.; Tauil, P.L.; Daniel-Ribeiro, C.T. Malaria in Brazil: An overview. Malar. J. 2010, 9, 115. [Google Scholar] [CrossRef] [Green Version]
- Forattini, O. P Culicidologia Médica, vol. 2: Identificação, Biologia, Epidemiologia; Edusp: São Paulo, Brazil, 2002. [Google Scholar]
- Carlos, B.C.; Rona, L.D.P.; Christophides, G.K.; Souza-Neto, J.A. A comprehensive analysis of malaria transmission in Brazil. Pathog. Glob. Health 2019, 113, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Deane, L.M.; Ferreira-Neto, J.A.; Deane, S.P.; Silveira, I.P. Anopheles (Kerteszia) cruzii, a natural vector of the monkey malaria parasites, Plasmodium simium and Plasmodium Brasilianum. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 647. [Google Scholar] [CrossRef]
- Deane, L.M.; Ferreira-Neto, J.A.; Lima, M.M. The vertical dispersion of Anopheles (Kerteszia) cruzii in a forest in southern Brazil suggests that human cases of malaria of simian origin might be expected. Mem. Inst. Oswaldo Cruz 1984, 79, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Demari-Silva, B.; Laporta, G.Z.; Oliveira, T.; Sallum, M. Plasmodium infection in Kerteszia cruzii (Diptera: Culicidae) in the Atlantic tropical rain forest, southeastern Brazil. Infect. Genet. Evol. 2020, 78, 104061. [Google Scholar] [CrossRef] [PubMed]
- Multini, L.C.; Wilke, A.B.B.; Marrelli, M.T. Neotropical Anopheles (Kerteszia) mosquitoes associated with bromeliad-malaria transmission in a changing world. Acta Trop. 2020, 205, 105413. [Google Scholar] [CrossRef] [PubMed]
- Zavortink, T.J. Mosquito studies XXIX. A review of the subgenus Kerteszia of Anopheles. Contr. Amer. Entomol. Inst. 1973, 9, 1–54. [Google Scholar]
- Foster, P.G.; de Oliveira, T.M.P.; Bergo, E.S.; Conn, J.E.; Sant’Ana, D.C.; Nagaki, S.S.; Nihei, S.; Lamas, C.E.; González, C.; Moreira, C.C.; et al. Phylogeny of Anophelinae using mitochondrial protein coding genes. R. Soc. Open Sci. 2017, 4, 170758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collucci, E.; Sallum, M.A.M. Phylogenetic analysis of the subgenus Kerteszia of Anopheles (Diptera: Culicidae: Anophelinae) based on morphological characters. Insect Syst. Evol. 2003, 34, 361–372. [Google Scholar] [CrossRef]
- Marrelli, M.T.; Malafronte, R.S.; Sallum, M.A.; Natal, D. Kerteszia subgenus of Anopheles associated with the Brazilian Atlantic rainforest: Current knowledge and future challenges. Malar. J. 2007, 6, 127. [Google Scholar] [CrossRef]
- Walter Reed Biosystematics Unit (WRBU). Systematic Catalog of Culicidae. 2019. Available online: http://www.mosquitocatalog.org/taxon_descr.aspx?ID=16082 (accessed on 31 August 2019).
- Guimarães, J.H. Systematic Database of Diptera of the Americas South of the United States (Family Culicidae); Plêiade/FAPESP: São Paulo, Brazil, 1997. [Google Scholar]
- Sallum, M.A.M.; Dos Santos, C.L.S.; Wilkerson, R.C. Studies on Anopheles (Kerteszia) homunculus Komp (Diptera: Culicidae). Zootaxa 2009, 2299, 1–18. [Google Scholar] [CrossRef]
- Harrison, B.A.; Ruiz-Lopez, F.; Falero, G.C.; Savage, H.M.; Pecor, J.E.; Wilkerson, R.C. Anopheles (Kerteszia) lepidotus (Diptera: Culicidae), not the malaria vector we thought it was: Revised male and female morphology; larva, pupa, and male genitalia characters; and molecular verification. Zootaxa 2012, 3218, 1–17. [Google Scholar] [CrossRef]
- Ramirez, C.C.; Dessen, E.M. Chromosome differentiated populations of Anopheles cruzii: Evidence for a third sibling species. Genetica 2000, 108, 73–80. [Google Scholar] [CrossRef]
- Oliveira, T.M.; Foster, P.G.; Bergo, E.S.; Nagaki, S.S.; Sanabani, S.S.; Marinotti, O.; Marinotti, P.N.; Sallum, M.A. Mitochondrial Genomes of Anopheles (Kerteszia) (Diptera: Culicidae) From the Atlantic Forest, Brazil. J. Med. Entomol. 2016, 53, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Multini, L.C.; Marrelli, M.T.; Beier, J.C.; Wilke, A.B.B. Increasing Complexity Threatens the Elimination of Extra-Amazonian Malaria in Brazil. Trends Parasitol. 2019, 35, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lopez, F.; Wilkerson, R.C.; Conn, J.E.; McKeon, S.N.; Levin, D.M.; Quiñones, M.L.; Póvoa, M.M.; Linton, Y.M. DNA barcoding reveals both known and novel taxa in the albitarsis group (Anopheles: Nyssorhynchus) of Neotropical malaria vectors. Parasit. Vectors 2012, 5, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourke, B.P.; Oliveira, T.P.; Suesdek, L.; Bergo, E.S.; Sallum, M.A. A multi-locus approach to barcoding in the Anopheles strodei subgroup (Diptera: Culicidae). Parasit. Vectors 2013, 6, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; deWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [Green Version]
- Cywinska, A.; Hunter, F.F.; Hebert, P.D.N. Identifying Canadian mosquito species through DNA barcodes. Med. Vet. Entomol. 2006, 20, 413–424. [Google Scholar] [CrossRef]
- Kumar, N.P.; Rajavel, A.R.; Natarajan, R.; Jambulingam, P. DNA barcodes can distinguish species of Indian mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 1–7. [Google Scholar] [CrossRef]
- Mirabello, L.; Conn, J.E. Molecular population genetics of the malaria vector Anopheles darlingi in Central and South America. Heredity 2006, 96, 311–321. [Google Scholar] [CrossRef]
- Gutiérrez, L.; Orrego, L.M.; Gómez, G.F.; López, A.; Luckhart, S.; Conn, J.E.; Correa, M.M. A new mtDNA COI gene lineage closely related to Anopheles janconnae of the Albitarsis complex in the Caribbean region of Colombia. Mem. Inst. Oswaldo Cruz 2010, 105, 1019–1025. [Google Scholar] [CrossRef]
- Jaramillo, L.M.; Gutiérrez, L.A.; Luckhart, S.; Conn, J.E.; Correa, M.M. Molecular evidence for a single taxon, Anopheles nuneztovari s.l., from two endemic malaria regions in Colombia. Mem. Inst. Oswaldo Cruz 2011, 106, 1017–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consoli, R.A.G.B.; Lourenço-de-Oliveira, R. Principais Mosquitos de Importância Sanitária No Brasil; Editora Fiocruz: Rio de Janeiro, Brazil, 1994. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Ruiz, F.; Linton, Y.M.; Ponsonby, D.J.; Conn, J.E.; Herrera, M.; Quiñones, M.L.; Vélez, I.D.; Wilkerson, R.C. Molecular comparison of topotypic specimens confirms Anopheles (Nyssorhynchus) dunhami Causey (Diptera: Culicidae) in the Colombian Amazon. Mem. Inst. Oswaldo Cruz 2010, 105, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Sallum, M.A.M.; Schultz, T.R.; Foster, P.G.; Aronstein, K.; Wirtz, R.A.; Wilkerson, R.C. Phylogeny of Anophelinae (Diptera: Culicidae) based on nuclear ribosomal and mitochondrial DNA sequences. Syst. Entomol. 2002, 27, 361–382. [Google Scholar] [CrossRef] [Green Version]
- Gorrochotegui-Escalante, N.; Munoz, M.L.; Fernandez-Salas, I.; Beaty, B.J.; Black, W.C. 4th. Genetic isolation by distance among Aedes aegypti populations along the northeastern coast of Mexico. Am. J. Trop. Med. Hyg. 2000, 62, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree: Tree Figure Drawing Tool Version 1.4.0, Institute of Evolutionary Biology, University of Edinburgh. 2006–2012. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 22 May 2019).
- Escovar, J.E.; González, R.; Quiñones, M.L.; Wilkerson, R.C.; Ruiz, F.; Harrison, B.A. Morphology of the larvae, male genitalia and DNA sequences of Anopheles (Kerteszia) pholidotus (Diptera: Culicidae) from Colombia. Mem. Inst. Oswaldo Cruz 2014, 109, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Freitas, L.A.; Russo, C.A.; Voloch, C.M.; Mutaquiha, O.C.; Marques, L.P.; Schrago, C.G. Diversification of the Genus Anopheles and a Neotropical Clade from the Late Cretaceous. PLoS ONE 2015, 10, e0134462. [Google Scholar] [CrossRef] [Green Version]
- Hua, Y.Q.; Ding, Y.R.; Yan, Z.T.; Si, F.L.; Luo, Q.C.; Chen, B. The complete mitochondrial genome of Anopheles minimus (Diptera: Culicidae) and the phylogenetics of known Anopheles mitogenomes. Insect Sci. 2016, 23, 353–365. [Google Scholar] [CrossRef] [PubMed]
- López-Rubio, A.; Suaza-Vasco, J.; Marcet, P.L.; Ruíz-Molina, N.; Cáceres, L.; Porter, C.; Uribe, S. Use of DNA barcoding to distinguish the malaria vector Anopheles neivai in Colombia. Zootaxa 2016, 4175, 377–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Li, C.; Zheng, W.; Song, F.; Guo, X.; Wu, Z.; Luo, P.; Yang, Y.; He, L.; Zhao, T. An evaluation of the suitability of COI and COII gene variation for reconstructing the phylogeny of, and identifying cryptic species in, anopheline mosquitoes (Diptera Culicidae). Mitochondrial Dna A Dna Mapp. Seq. Anal. 2017, 28, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, Y.; Li, X.Y.; Peng, H.; Ma, Y.J. Sequencing and analysis of the complete mitochondrial genome in Anopheles sinensis (Diptera: Culicidae). Infect. Dis. Poverty 2017, 6, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Rubio, A.; Suaza, J.D.; Porter, C.; Uribe, S.; Bedoya, G.; Vélez, I.D. Phylogenetic signal at the Cytb-SertRNA-IG1-ND1 mitochondrial region in Anopheles (Kerteszia) neivai Howard, Dyar & Knab, 1913. Biomedica 2017, 37, 143–154. [Google Scholar] [PubMed] [Green Version]
- Lorenz, C.; Patané, J.S.; Suesdek, L. Morphogenetic characterisation, date of divergence, and evolutionary relationships of malaria vectors Anopheles cruzii and Anopheles homunculus. Infect. Genet. Evol. 2015, 35, 144–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, C.C.L.; Dessen, E.M.B. The polytene chromosomes of the mosquito Anopheles bellator compared with those of Anopheles cruzii. Braz. J. Genet. 1996, 19, 555–558. [Google Scholar] [CrossRef] [Green Version]
- Rona, L.D.; Carvalho-Pinto, C.J.; Gentile, C.; Grisard, E.C.; Peixoto, A.A. Assessing the molecular divergence between Anopheles (Kerteszia) cruzii populations from Brazil using the timeless gene: Further evidence of a species complex. Malar. J. 2009, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Rona, L.D.; Carvalho-Pinto, C.J.; Mazzoni, C.J.; Peixoto, A.A. Estimation of divergence time between two sibling species of the Anopheles (Kerteszia) cruzii complex using a multilocus approach. BMC Evol. Biol. 2010, 10, 91. [Google Scholar] [CrossRef] [Green Version]
- Rona, L.D.; Carvalho-Pinto, C.J.; Peixoto, A.A. Evidence for the occurrence of two sympatric sibling species within the Anopheles (Kerteszia) cruzii complex in southeast Brazil and the detection of asymmetric introgression between them using a multilocus analysis. BMC Evol. Biol. 2013, 13, 207. [Google Scholar] [CrossRef] [Green Version]
- Rona, L.D.; Carvalho-Pinto, C.J.; Peixoto, A.A. Molecular evidence for the occurrence of a new sibling species within the Anopheles (Kerteszia) cruzii complex in south-east Brazil. Malar. J. 2010, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Rezende Dias, G.; Fujii, T.T.S.; Fogel, B.F.; Lourenço-de-Oliveira, R.; Silva-do-Nascimento, T.F.; Pitaluga, A.N.; Carvalho-Pinto, C.J.; Carvalho, A.B.; Peixoto, A.A.; Rona, L.D.P. Cryptic diversity in an Atlantic Forest malaria vector from the mountains of South-East Brazil. Parasit. Vectors 2018, 11, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Mosquito ID | Origin | Collection Year | Species (According to Morphological Identification) | Species (According to Phylogeny Analysis) | COI * (COIF/COIR Primers) | ND4 * | COI * (LCO/HCO Primers) |
|---|---|---|---|---|---|---|---|
| 1A | Miraflores, Yanatile, Calca, Cusco, Peru | 2007 | Anopheles (Kerteszia) boliviensis | Anopheles (Kerteszia) boliviensis | MH589387 | MH560305 | -- |
| 1B | Miraflores, Yanatile, Calca, Cusco, Peru | 2007 | Anopheles (Kerteszia) boliviensis | Anopheles (Kerteszia) boliviensis | MH589388 | MH560306 | MH589377 |
| 2A | Corimayo, Yanatile, Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) boliviensis | Anopheles (Kerteszia) boliviensis | MH589389 | MH560307 | -- |
| 2B | Corimayo, Yanatile, Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) boliviensis | Anopheles (Kerteszia) boliviensis | MH589390 | MH560308 | MH589378 |
| 3A | Colca, Yanatile, Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) boliviensis | Anopheles (Kerteszia) boliviensis | MH589391 | MH560309 | MH589379 |
| 3B | Colca, Yanatile Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) boliviensis | Anopheles (Kerteszia) boliviensis | MH589392 | MH560310 | MH589380 |
| 4A | Miraflores, Yanatile, Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) boliviensis | MH589393 | MH560311 | MH589381 |
| 4B | Miraflores, Yanatile, Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) boliviensis | MH589394 | MH563012 | MH589382 |
| 5A | Colca, Yanatile, Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) lepidotus | Anopheles (Kerteszia) pholidotus | MH589395 | MH563013 | -- |
| 5B | Colca, Yanatile, Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) lepidotus | Anopheles (Kerteszia) pholidotus | MH589396 | MH560314 | -- |
| 6A | Miraflores, Yanatile, Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) lepidotus | Anopheles (Kerteszia) boliviensis | MH589397 | MH560315 | MH589383 |
| 6B | Miraflores, Yanatile, Calca, Cusco, Peru | 2008 | Anopheles (Kerteszia) lepidotus | Anopheles (Kerteszia) pholidotus | MH589398 | MH563016 | MH589384 |
| LAN | Selva Alegre, Alto Inambari, Sandia, Puno, Peru | 2000 | Anopheles (Kerteszia) laneanus | Anopheles (Kerteszia) boliviensis | MH589409 | --- | MH589367 |
| Mosquito ID | Origin | Collection Year | Species (According Morphological Identification) | Species (According Phylogeny Analysis) | COI * (COIF/COIR Primers) | ND4 * | COI * (LCO/HCO Primers) |
|---|---|---|---|---|---|---|---|
| CA1 | Caraguatatuba, SP, Brazil | 1946 | Anopheles (Kerteszia) bellator | Anopheles (Kerteszia) cruzii | MH589406 | MH560328 | MH589361 |
| B1 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) bellator | Anopheles (Kerteszia) cruzii | -- | MH560317 | MH589362 |
| B2 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) bellator | Anopheles (Kerteszia) cruzii | MH589399 | MH560318 | MH589363 |
| B3 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) bellator | Anopheles (Kerteszia) cruzii | -- | MH560319 | MH589364 |
| PAC1 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589412 | MH560333 | MH589352 |
| PAC2 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589414 | MH560335 | MH589353 |
| PAC3 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589415 | MH560336 | MH589354 |
| PAC4 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589416 | MH560337 | MH589355 |
| PAC5 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589417 | MH560338 | MH589356 |
| PAC6 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589418 | -- | MH589357 |
| PAC7 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589419 | MH560339 | MH589358 |
| PAC8 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589420 | MH560340 | MH589359 |
| PAC9 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589421 | MH560341 | MH589360 |
| PAC10X | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589413 | MH560334 | -- |
| C1 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | -- | MH560320 | MH589369 |
| C2 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589400 | MH560321 | MH589370 |
| C3 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | -- | MH560322 | MH589371 |
| C4 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589401 | MH560323 | MH589372 |
| C5 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589402 | MH560324 | MH589373 |
| C6 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589403 | MH560325 | MH589374 |
| C7 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589404 | MH560326 | MH589375 |
| C8 | Marsilac, São Paulo, SP, Brazil | 2009 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589405 | MH560327 | MH589376 |
| L1 | Campos do Jordão, SP, Brazil | 1997 | Anopheles (Kerteszia) laneanus | Anopheles (Kerteszia) laneanus | MH589407 | MH560329 | MH589365 |
| L3 | Campos do Jordão, SP, Brazil | 1997 | Anopheles (Kerteszia) laneanus | Anopheles (Kerteszia) laneanus | MH589408 | MH560330 | MH589366 |
| M95 | Itanhaém, SP, Brazil | 2010 | Anopheles (Kerteszia) cruzii | Anopheles (Kerteszia) cruzii | MH589411 | MH560332 | MH589386 |
| M83 | Itanhaém, SP, Brazil | 2010 | Anopheles (Anopheles) maculipes/pseudomaculipes | Anopheles (Anopheles) intermedius | MH589410 | MH560331 | MH589385 |
| PAN1 | Parelheiros, São Paulo, SP, Brazil | 2002 | Anopheles (Nyssorhynchus) sp. | Anopheles (Nysssorhynchus) strodei | -- | MH560342 | MH589368 |
| Species | Mosquito ID | Origin | # GenBank |
|---|---|---|---|
| An. lepidotus | 51917879 | Napo/Ecuador | JQ041282 * |
| An. lepidotus | 51917894 | Napo/Ecuador | JQ041284 * |
| An. pholidotus | 51917595 | Tachira/Venezuela | JQ041287 * |
| An. pholidotus | 51917607 | Tachira/Venezuela | JQ041288 * |
| An. homunculus | BA22_33 | Camacan, BA/Brazil | JQ291234 * |
| An. homunculus | ES10_2 | Santa Teresa, ES/Brazil | JQ291235 * |
| An. homunculus | RS20_21 | Maquiné, RS/Brazil | JQ291236 * |
| An. homunculus | RS20_33 | Maquiné, RS/Brazil | JQ291237 * |
| An. homunculus | RS30_56 | Maquiné, RS/Brazil | JQ291238 * |
| An. homunculus | SP23_1 | Cananéia, SP/Brazil | JQ291240 * |
| An. homunculus | ST26 | Cananéia, SP/Brazil | JQ291242 * |
| An. lepidotus | CCDB-10764-A9 | Orellana/Ecuador | KF670991 * |
| An. cruzii | ??? | Cananéia/SP | KJ701506 |
| An. cruzii | 104 | Tapiraí, SP/Brazil | KM507293 * |
| An. cruzii | 110 | Tapiraí, SP/Brazil | KM507294 * |
| An. cruzii | 112 | Tapiraí, SP/Brazil | KM507295 * |
| An. cruzii | 127 | Tapiraí, SP/Brazil | KM507296 * |
| An. cruzii | 192 | Tapiraí, SP/Brazil | KM507297 * |
| An. cruzii | 198 | Tapiraí, SP/Brazil | KM507298 * |
| An. cruzii | 199 | Tapiraí, SP/Brazil | KM507299 * |
| An. cruzii | 200 | Tapiraí, SP/Brazil | KM507300 * |
| An. cruzii | 217 | Tapiraí, SP/Brazil | KM507301 * |
| An. cruzii | 240 | Tapiraí, SP/Brazil | KM507302 * |
| An. cruzii | 241 | Tapiraí, SP/Brazil | KM507303 * |
| An. cruzii | 242 | Tapiraí, SP/Brazil | KM507304 * |
| An. cruzii | 255 | Tapiraí, SP/Brazil | KM507305 * |
| An. cruzii | 257 | Tapiraí, SP/Brazil | KM507306 * |
| An. cruzii | 258 | Tapiraí, SP/Brazil | KM507307 * |
| An. cruzii | 259 | Tapiraí, SP/Brazil | KM507308 * |
| An. cruzii | 260 | Tapiraí, SP/Brazil | KM507309 * |
| An. cruzii | 261 | Tapiraí, SP/Brazil | KM507310 * |
| An. cruzii | 262 | Tapiraí, SP/Brazil | KM507311 * |
| An. cruzii | 267 | Tapiraí, SP/Brazil | KM507312 * |
| An. cruzii | 269 | Tapiraí, SP/Brazil | KM507314 * |
| An. homunculus | BA22_32 | Camacan/BA | KU551283 |
| An. cruzii | PEC_2_7 | São Paulo/SP | KU551284 |
| An. cruzii | RJ03_2 | Itatiaia/RJ | KU551285 |
| An. cruzii | RS32_11_7 | Maquiné/RS | KU551286 |
| An. bellator | SP24_3_1 | Cananéia/SP | KU551287 |
| An. laneanus | SP52-103 | Pindamonhangaba/SP | KU551288 |
| An. cruzii | ST16 | Cananéia/SP | KU551289 |
| An. neivai | MB10165 | Petit-Saut/French Guyana | MF172273 * |
| An. neivai | MB10252 | Petit-Saut/French Guyana | MF172274 * |
| An. neivai | MB10253 | Petit-Saut/French Guyana | MF172275 * |
| An. neivai | MB10254 | Petit-Saut/French Guyana | MF172276 * |
| An. homunculus | BA22_31 | Camacan/BA | MF381605 |
| An. laneanus | SP52-17 | Pindamonhangaba/SP | MF381613 |
| An. homunculus | RS30_158 | Maquiné/RS | MF381697 |
| An. (An.) intermedius | SP02_8_1 | Pariquera-Açú/SP | MF381700 |
| An. (Nyr.) strodei | SP104_18_1 | Pindamonhangaba/SP | MF381724 * |
| An. homunculus | ST19 | Cananéia/SP | MF381729 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirchgatter, K.; de Oliveira Guimarães, L.; Hugo Yañez Trujillano, H.; Rafael Arias, F.; Cáceres, A.G.; de Castro Duarte, A.M.R.; dos Santos Malafronte, R.; Tubaki, R.M.; Mureb Sallum, M.A. Phylogeny of Anopheles (Kerteszia) (Diptera: Culicidae) Using Mitochondrial Genes. Insects 2020, 11, 324. https://doi.org/10.3390/insects11050324
Kirchgatter K, de Oliveira Guimarães L, Hugo Yañez Trujillano H, Rafael Arias F, Cáceres AG, de Castro Duarte AMR, dos Santos Malafronte R, Tubaki RM, Mureb Sallum MA. Phylogeny of Anopheles (Kerteszia) (Diptera: Culicidae) Using Mitochondrial Genes. Insects. 2020; 11(5):324. https://doi.org/10.3390/insects11050324
Chicago/Turabian StyleKirchgatter, Karin, Lilian de Oliveira Guimarães, Henrry Hugo Yañez Trujillano, Fernando Rafael Arias, Abraham Germán Cáceres, Ana Maria Ribeiro de Castro Duarte, Rosely dos Santos Malafronte, Rosa Maria Tubaki, and Maria Anice Mureb Sallum. 2020. "Phylogeny of Anopheles (Kerteszia) (Diptera: Culicidae) Using Mitochondrial Genes" Insects 11, no. 5: 324. https://doi.org/10.3390/insects11050324
APA StyleKirchgatter, K., de Oliveira Guimarães, L., Hugo Yañez Trujillano, H., Rafael Arias, F., Cáceres, A. G., de Castro Duarte, A. M. R., dos Santos Malafronte, R., Tubaki, R. M., & Mureb Sallum, M. A. (2020). Phylogeny of Anopheles (Kerteszia) (Diptera: Culicidae) Using Mitochondrial Genes. Insects, 11(5), 324. https://doi.org/10.3390/insects11050324