Adult Mosquitoes Infected with Bacteria Early in Life Have Stronger Antimicrobial Responses and More Hemocytes after Reinfection Later in Life



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing and Maintenance
2.2. General Experimental Design
2.3. Intravital Injections and Infections
2.4. Bacterial Infection Intensity
2.5. Quantification of the Number of Circulating Hemocytes Per Mosquito
2.6. Quantification of Phagocytosis by Circulating Hemocytes
2.7. Quantification of Gene Expression by Real-Time PCR (RT-PCR)
2.8. Mosquito Survival
3. Results
3.1. A Bacterial Infection in the Hemocoel Early in Life Increases the Ability of Mosquitoes to Kill Bacteria Acquired Later in Life
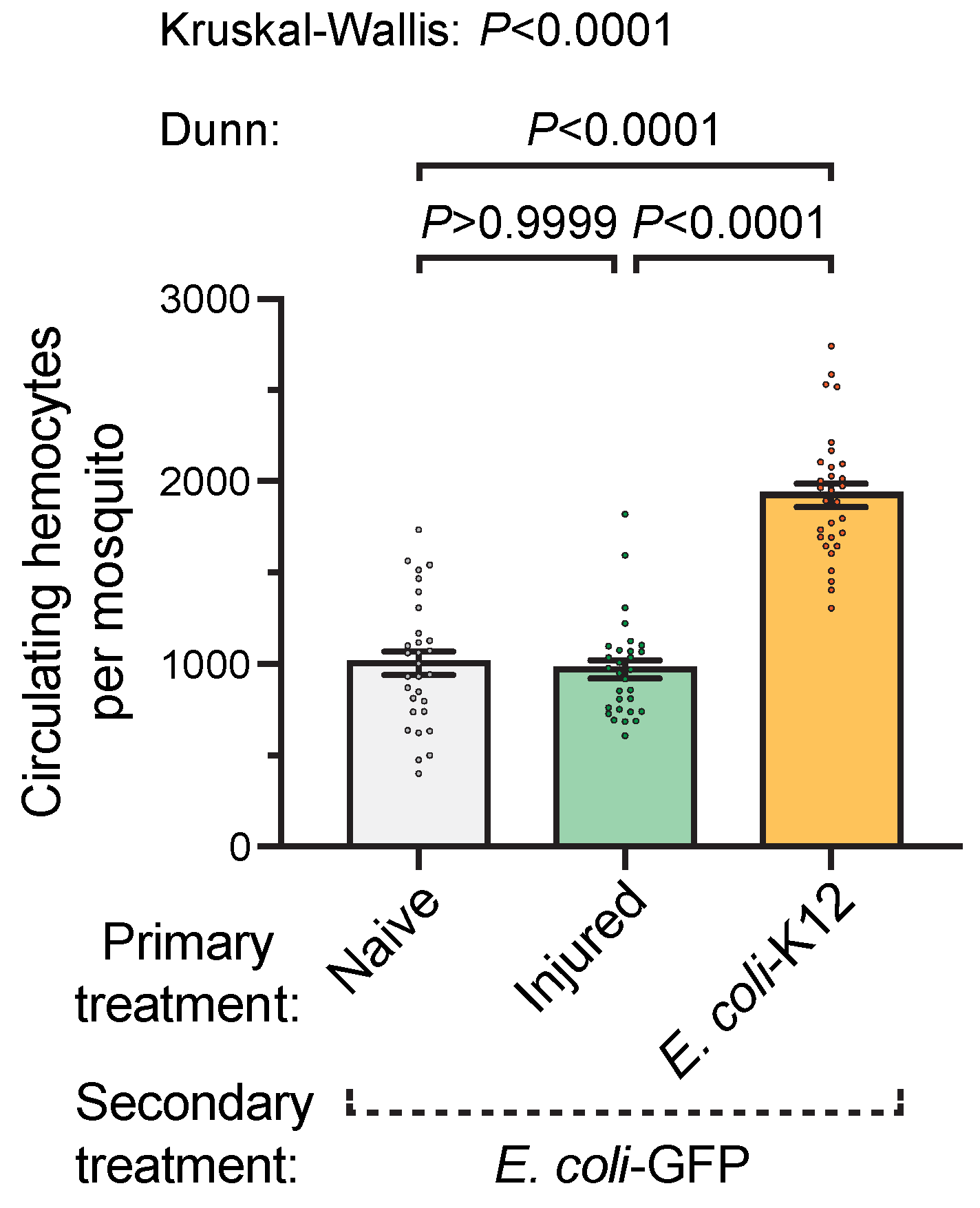
3.2. A Prior Infection Augments the Infection-Induced Increase in the Number of Circulating Hemocytes
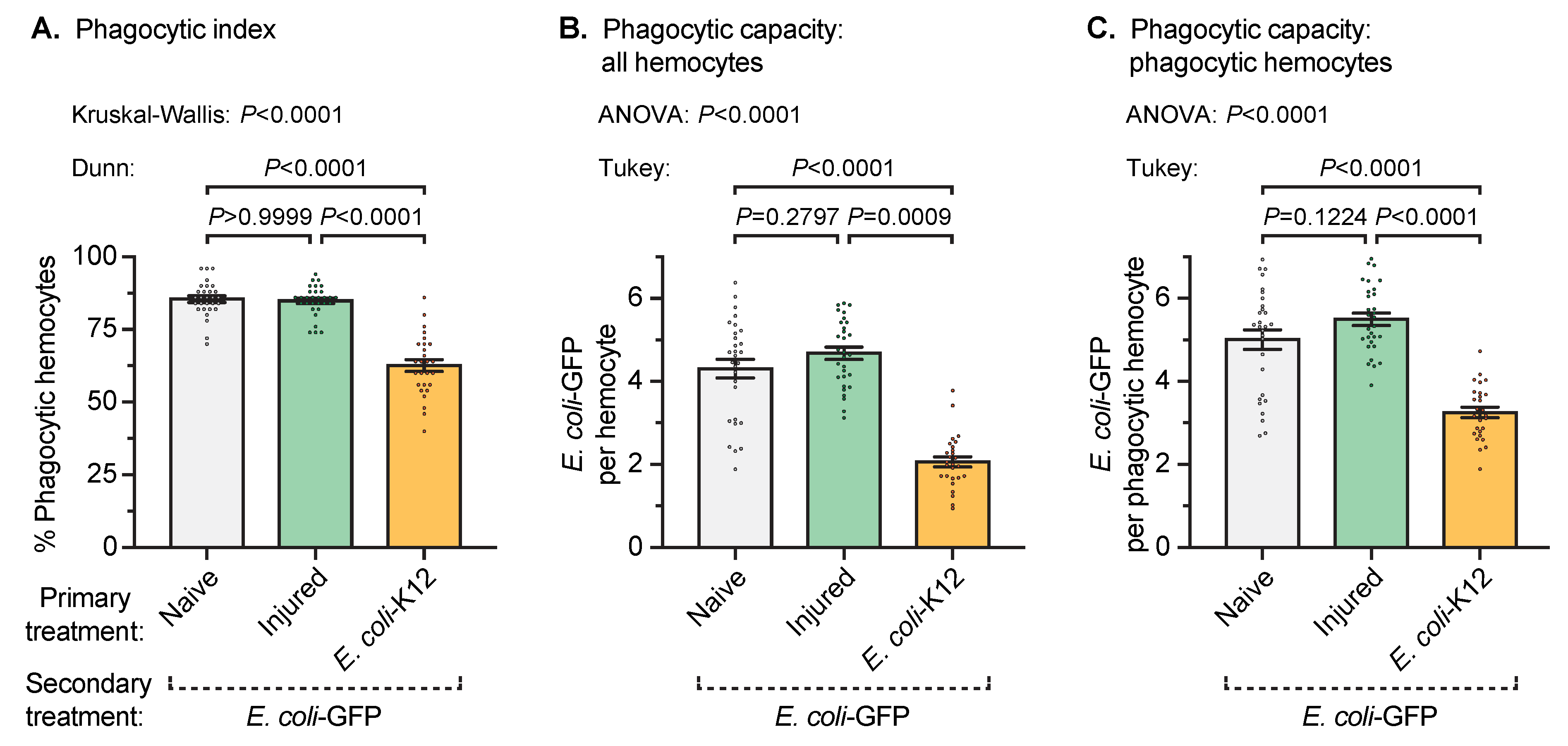
3.3. A Prior Infection Decreases the Phagocytic Activity of Individual Hemocytes Against an Infection Acquired Later in Life
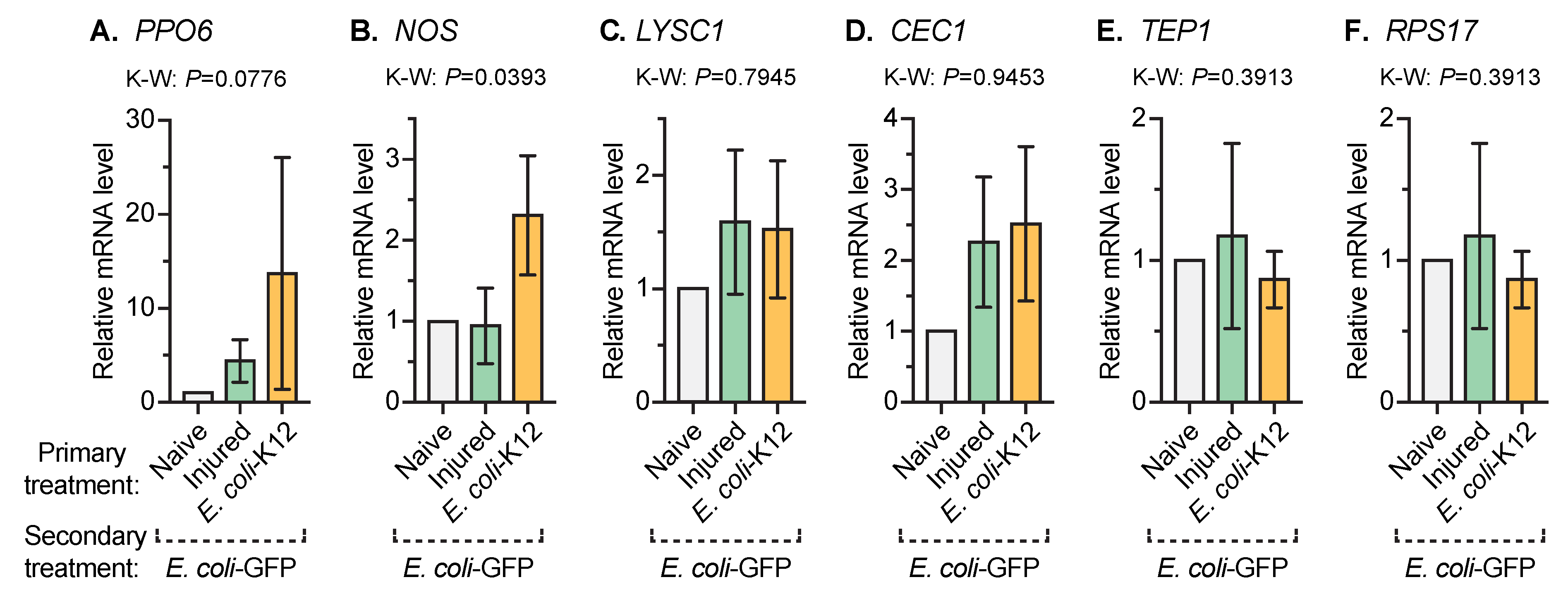
3.4. An Infection Early in Life Increases the Expression of Nitric Oxide Synthase When Mosquitoes Acquire Another Infection Later in Life
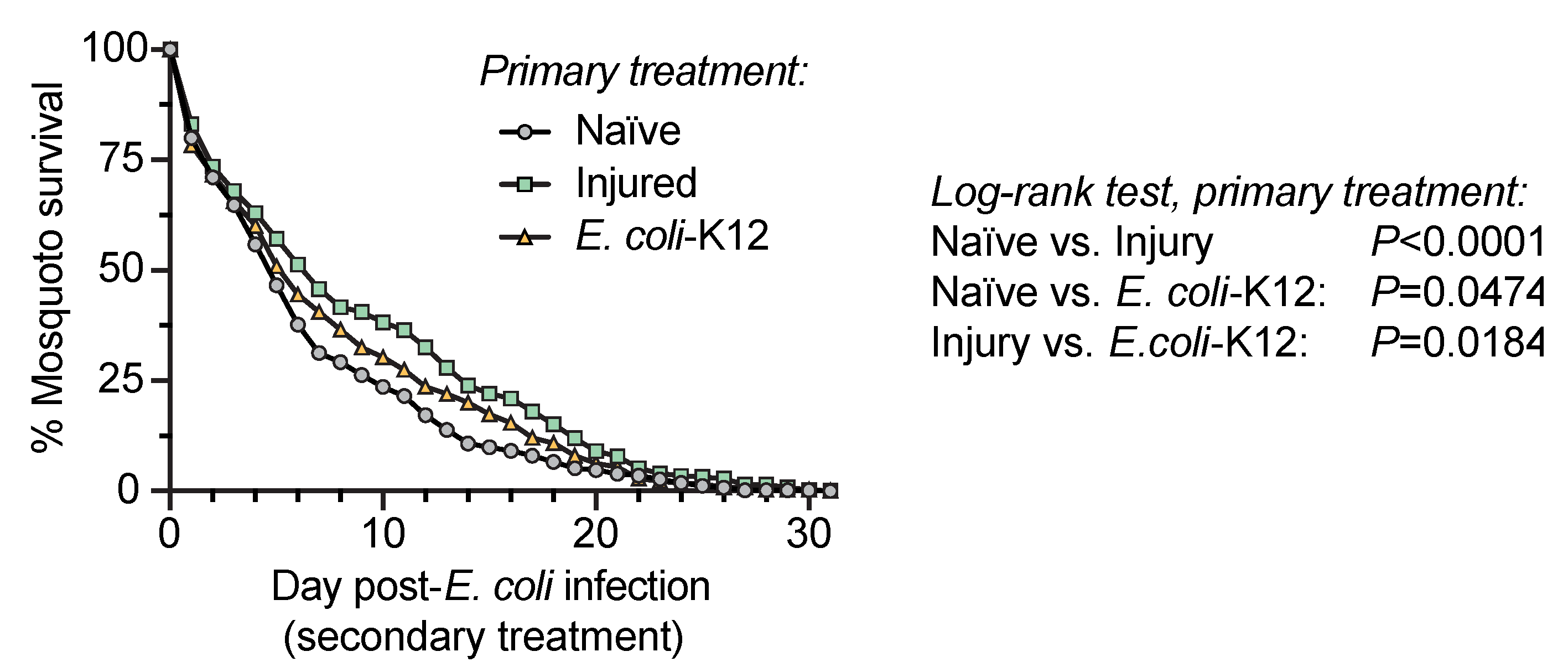
3.5. Injury and Infection Early in Life Improves the Survival of Mosquitoes that Become Infected with Bacteria Later in Life
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Delves, P.J.; Martin, S.J.; Burton, D.R.; Roitt, I.M. Roitt’s Essential Immunology, 12th ed.; Blackwell Publishing Ltd.: West Sussex, UK, 2011. [Google Scholar]
- Masri, L.; Cremer, S. Individual and social immunisation in insects. Trends Immunol. 2014, 35, 471–482. [Google Scholar] [CrossRef]
- Shaw, D.K.; Tate, A.T.; Schneider, D.S.; Levashina, E.A.; Kagan, J.C.; Pal, U.; Fikrig, E.; Pedra, J.H.F. Vector immunity and evolutionary ecology: The harmonious dissonance. Trends Immunol. 2018, 39, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, G.; Farrell, G.; Kavanagh, K. Immune priming: The secret weapon of the insect world. Virulence 2020, 11, 238–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, D.; Eleftherianos, I. Memory and specificity in the insect immune system: Current perspectives and future challenges. Front. Immunol. 2017, 8, 539. [Google Scholar] [CrossRef] [PubMed]
- Melillo, D.; Marino, R.; Italiani, P.; Boraschi, D. Innate immune memory in invertebrate metazoans: A critical appraisal. Front. Immunol. 2018, 9, 1915. [Google Scholar] [CrossRef] [PubMed]
- Dhinaut, J.; Chogne, M.; Moret, Y. Immune priming specificity within and across generations reveals the range of pathogens affecting evolution of immunity in an insect. J. Anim. Ecol. 2018, 87, 448–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferro, K.; Peuss, R.; Yang, W.; Rosenstiel, P.; Schulenburg, H.; Kurtz, J. Experimental evolution of immunological specificity. Proc. Natl. Acad. Sci. USA 2019, 116, 20598–20604. [Google Scholar] [CrossRef] [Green Version]
- Pham, L.N.; Dionne, M.S.; Shirasu-Hiza, M.; Schneider, D.S. A specific primed immune response in Drosophila is dependent on phagocytes. PLoS Pathog. 2007, 3, e26. [Google Scholar] [CrossRef]
- Moret, Y.; Siva-Jothy, M.T. Adaptive innate immunity? Responsive-mode prophylaxis in the mealworm beetle, Tenebrio molitor. Proc. Biol. Sci. 2003, 270, 2475–2480. [Google Scholar] [CrossRef] [Green Version]
- Sadd, B.M.; Schmid-Hempel, P. Insect immunity shows specificity in protection upon secondary pathogen exposure. Curr. Biol. 2006, 16, 1206–1210. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Tokman, D.M.; Gonzalez-Santoyo, I.; Lanz-Mendoza, H.; Aguilar, A.C. Territorial damselflies do not show immunological priming in the wild. Physiol. Entomol. 2010, 35, 364–372. [Google Scholar] [CrossRef]
- Reber, A.; Chapuisat, M. No evidence for immune priming in ants exposed to a fungal pathogen. PLoS ONE 2012, 7, e35372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrnogic, J.; Castillo, J.C.; Shokal, U.; Yadav, S.; Kenney, E.; Heryanto, C.; Ozakman, Y.; Eleftherianos, I. Pre-exposure to non-pathogenic bacteria does not protect Drosophila against the entomopathogenic bacterium Photorhabdus. PLoS ONE 2018, 13, e0205256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futo, M.; Sell, M.P.; Kutzer, M.A.M.; Kurtz, J. Specificity of oral immune priming in the red flour beetle Tribolium castaneum. Biol. Lett. 2017, 13, 20170632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargielowski, I.; Koella, J.C. A possible mechanism for the suppression of Plasmodium berghei development in the mosquito Anopheles gambiae by the microsporidian Vavraia culicis. PLoS ONE 2009, 4, e4676. [Google Scholar] [CrossRef]
- Contreras-Garduno, J.; Rodriguez, M.C.; Hernandez-Martinez, S.; Martinez-Barnetche, J.; Alvarado-Delgado, A.; Izquierdo, J.; Herrera-Ortiz, A.; Moreno-Garcia, M.; Velazquez-Meza, M.E.; Valverde, V.; et al. Plasmodium berghei induced priming in Anopheles albimanus independently of bacterial co-infection. Dev. Comp. Immunol. 2015, 52, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Garduno, J.; Rodriguez, M.C.; Rodriguez, M.H.; Alvarado-Delgado, A.; Lanz-Mendoza, H. Cost of immune priming within generations: Trade-off between infection and reproduction. Microbes Infect. 2014, 16, 261–267. [Google Scholar] [CrossRef]
- Lowenberger, C.A.; Kamal, S.; Chiles, J.; Paskewitz, S.; Bulet, P.; Hoffmann, J.A.; Christensen, B.M. Mosquito-Plasmodium interactions in response to immune activation of the vector. Exp. Parasitol. 1999, 91, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.; Brayner, F.A.; Alves, L.C.; Dixit, R.; Barillas-Mury, C. Hemocyte differentiation mediates innate immune memory in Anopheles gambiae mosquitoes. Science 2010, 329, 1353–1355. [Google Scholar] [CrossRef] [Green Version]
- Herren, J.K.; Mbaisi, L.; Mararo, E.; Makhulu, E.E.; Mobegi, V.A.; Butungi, H.; Mancini, M.V.; Oundo, J.W.; Teal, E.T.; Pinaud, S.; et al. A microsporidian impairs Plasmodium falciparum transmission in Anopheles arabiensis mosquitoes. Nat. Commun. 2020, 11, 2187. [Google Scholar] [CrossRef]
- Kambris, Z.; Cook, P.E.; Phuc, H.K.; Sinkins, S.P. Immune activation by life-shortening Wolbachia and reduced filarial competence in mosquitoes. Science 2009, 326, 134–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowenberger, C.A.; Ferdig, M.T.; Bulet, P.; Khalili, S.; Hoffmann, J.A.; Christensen, B.M. Aedes aegypti: Induced antibacterial proteins reduce the establishment and development of Brugia malayi. Exp. Parasitol. 1996, 83, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.H.; Woolfit, M.; Rances, E.; O’Neill, S.L.; McGraw, E.A. Wolbachia-associated bacterial protection in the mosquito Aedes aegypti. PLoS Negl. Trop. Dis. 2013, 7, e2362. [Google Scholar] [CrossRef] [Green Version]
- Aliota, M.T.; Peinado, S.A.; Velez, I.D.; Osorio, J.E. The wMel strain of Wolbachia reduces transmission of Zika virus by Aedes aegypti. Sci. Rep. 2016, 6, 28792. [Google Scholar] [CrossRef]
- Serrato-Salas, J.; Izquierdo-Sanchez, J.; Arguello, M.; Conde, R.; Alvarado-Delgado, A.; Lanz-Mendoza, H. Aedes aegypti antiviral adaptive response against DENV-2. Dev. Comp. Immunol. 2018, 84, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Cappelli, A.; Damiani, C.; Mancini, M.V.; Valzano, M.; Rossi, P.; Serrao, A.; Ricci, I.; Favia, G. Asaia activates immune genes in mosquito eliciting an anti-Plasmodium response: Implications in malaria control. Front. Genet. 2019, 10, 836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, V.; Moreno-Garcia, M.; Duarte-Elguea, E.; Lanz-Mendoza, H. Limited specificity in the injury and infection priming against bacteria in Aedes aegypti mosquitoes. Front. Microbiol. 2016, 7, 975. [Google Scholar] [CrossRef]
- Dodson, B.L.; Hughes, G.L.; Paul, O.; Matacchiero, A.C.; Kramer, L.D.; Rasgon, J.L. Wolbachia enhances West Nile virus (WNV) infection in the mosquito Culex tarsalis. PLoS Negl. Trop. Dis. 2014, 8, e2965. [Google Scholar] [CrossRef]
- Joshi, D.; Pan, X.; McFadden, M.J.; Bevins, D.; Liang, X.; Lu, P.; Thiem, S.; Xi, Z. The maternally inheritable Wolbachia wAlbB induces refractoriness to Plasmodium berghei in Anopheles stephensi. Front. Microbiol. 2017, 8, 366. [Google Scholar] [CrossRef] [Green Version]
- Hughes, G.L.; Koga, R.; Xue, P.; Fukatsu, T.; Rasgon, J.L. Wolbachia infections are virulent and inhibit the human malaria parasite Plasmodium falciparum in Anopheles gambiae. PLoS Pathog. 2011, 7, e1002043. [Google Scholar] [CrossRef] [Green Version]
- Murdock, C.C.; Blanford, S.; Hughes, G.L.; Rasgon, J.L.; Thomas, M.B. Temperature alters Plasmodium blocking by Wolbachia. Sci. Rep. 2014, 4, 3932. [Google Scholar] [CrossRef] [Green Version]
- Zele, F.; Nicot, A.; Berthomieu, A.; Weill, M.; Duron, O.; Rivero, A. Wolbachia increases susceptibility to Plasmodium infection in a natural system. Proc. Biol. Sci. 2014, 281, 20132837. [Google Scholar] [CrossRef]
- Estevez-Lao, T.Y.; Boyce, D.S.; Honegger, H.W.; Hillyer, J.F. Cardioacceleratory function of the neurohormone CCAP in the mosquito Anopheles gambiae. J. Exp. Biol. 2013, 216, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Coggins, S.A.; Estevez-Lao, T.Y.; Hillyer, J.F. Increased survivorship following bacterial infection by the mosquito Aedes aegypti as compared to Anopheles gambiae correlates with increased transcriptional induction of antimicrobial peptides. Dev. Comp. Immunol. 2012, 37, 390–401. [Google Scholar] [CrossRef]
- Hillyer, J.F.; Schmidt, S.L.; Fuchs, J.F.; Boyle, J.P.; Christensen, B.M. Age-associated mortality in immune challenged mosquitoes (Aedes aegypti) correlates with a decrease in haemocyte numbers. Cell. Microbiol. 2005, 7, 39–51. [Google Scholar] [CrossRef] [PubMed]
- League, G.P.; Estevez-Lao, T.Y.; Yan, Y.; Garcia-Lopez, V.A.; Hillyer, J.F. Anopheles gambiae larvae mount stronger immune responses against bacterial infection than adults: Evidence of adaptive decoupling in mosquitoes. Parasit Vectors 2017, 10, 367. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gorman, M.J.; Paskewitz, S.M. Persistence of infection in mosquitoes injected with bacteria. J. Invertebr. Pathol. 2000, 75, 296–297. [Google Scholar] [CrossRef] [PubMed]
- King, J.G.; Hillyer, J.F. Infection-induced interaction between the mosquito circulatory and immune systems. PLoS Pathog. 2012, 8, e1003058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, O.; Sadd, B.M.; Schmid-Hempel, P.; Kurtz, J. Strain-specific priming of resistance in the red flour beetle, Tribolium castaneum. Proc. Biol. Sci. 2009, 276, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Li, M.; Liu, Y.; Ding, Y.; Yi, Y. The specificity of immune priming in silkworm, Bombyx mori, is mediated by the phagocytic ability of granular cells. J. Insect Physiol. 2015, 81, 60–68. [Google Scholar] [CrossRef]
- Hillyer, J.F.; Strand, M.R. Mosquito hemocyte-mediated immune responses. Curr. Opin. Insect Sci. 2014, 3, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Baton, L.A.; Robertson, A.; Warr, E.; Strand, M.R.; Dimopoulos, G. Genome-wide transcriptomic profiling of Anopheles gambiae hemocytes reveals pathogen-specific signatures upon bacterial challenge and Plasmodium berghei infection. BMC Genom. 2009, 10, 257. [Google Scholar] [CrossRef] [Green Version]
- King, J.G.; Hillyer, J.F. Spatial and temporal in vivo analysis of circulating and sessile immune cells in mosquitoes: Hemocyte mitosis following infection. BMC Biol. 2013, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.D.; Shapiro, L.L.M.; Thompson, G.A.; Estevez-Lao, T.Y.; Hillyer, J.F. Transstadial immune activation in a mosquito: Adults that emerge from infected larvae have stronger antibacterial activity in their hemocoel yet increased susceptibility to malaria infection. Ecol. Evol. 2019, 9, 6082–6095. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; de Almeida Oliveira, G.; Calvo, E.; Dalli, J.; Colas, R.A.; Serhan, C.N.; Ribeiro, J.M.; Barillas-Mury, C. A mosquito lipoxin/lipocalin complex mediates innate immune priming in Anopheles gambiae. Nat. Commun. 2015, 6, 7403. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Garver, L.S.; Brayner, F.A.; Alves, L.C.; Rodrigues, J.; Molina-Cruz, A.; Barillas-Mury, C. The role of hemocytes in Anopheles gambiae antiplasmodial immunity. J Innate Immun. 2014, 6, 119–128. [Google Scholar] [CrossRef]
- Smith, R.C.; King, J.G.; Tao, D.; Zeleznik, O.A.; Brando, C.; Thallinger, G.G.; Dinglasan, R.R. Molecular profiling of phagocytic immune cells in Anopheles gambiae reveals integral roles for hemocytes in mosquito innate immunity. Mol. Cell. Proteom. 2016, 15, 3373–3387. [Google Scholar] [CrossRef] [Green Version]
- Bryant, W.B.; Michel, K. Blood feeding induces hemocyte proliferation and activation in the African malaria mosquito, Anopheles gambiae Giles. J. Exp. Biol. 2014, 217, 1238–1245. [Google Scholar] [CrossRef] [Green Version]
- Bryant, W.B.; Michel, K. Anopheles gambiae hemocytes exhibit transient states of activation. Dev. Comp. Immunol. 2016, 55, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.; Brown, M.R.; Strand, M.R. Blood feeding and insulin-like peptide 3 stimulate proliferation of hemocytes in the mosquito Aedes aegypti. PLoS Pathog. 2011, 7, e1002274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, R.A.; Kwon, H.; Smith, R.C. 20-Hydroxyecdysone primes innate immune responses that limit bacterial and malarial parasite survival in Anopheles gambiae. mSphere 2020, 5, e00983-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upton, L.M.; Povelones, M.; Christophides, G.K. Anopheles gambiae blood feeding initiates an anticipatory defense response to Plasmodium berghei. J. Innate Immun. 2015, 7, 74–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallon, J.P.; Troy, N.; Kavanagh, K. Pre-exposure of Galleria mellonella larvae to different doses of Aspergillus fumigatus conidia causes differential activation of cellular and humoral immune responses. Virulence 2011, 2, 413–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwood, J.M.; Milutinovic, B.; Peuss, R.; Behrens, S.; Esser, D.; Rosenstiel, P.; Schulenburg, H.; Kurtz, J. Oral immune priming with Bacillus thuringiensis induces a shift in the gene expression of Tribolium castaneum larvae. BMC Genom. 2017, 18, 329. [Google Scholar] [CrossRef]
- Bergin, D.; Murphy, L.; Keenan, J.; Clynes, M.; Kavanagh, K. Pre-exposure to yeast protects larvae of Galleria mellonella from a subsequent lethal infection by Candida albicans and is mediated by the increased expression of antimicrobial peptides. Microbes Infect. 2006, 8, 2105–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vertyporokh, L.; Wojda, I. Immune response of Galleria mellonella after injection with non-lethal and lethal dosages of Candida albicans. J. Invertebr. Pathol. 2020, 170, 107327. [Google Scholar] [CrossRef]
- Hillyer, J.F.; Estevez-Lao, T.Y. Nitric oxide is an essential component of the hemocyte-mediated mosquito immune response against bacteria. Dev. Comp. Immunol. 2010, 34, 141–149. [Google Scholar] [CrossRef]
- Estevez-Lao, T.Y.; Sigle, L.T.; Gomez, S.N.; Hillyer, J.F. Nitric oxide produced by periostial hemocytes modulates the bacterial infection induced reduction of the mosquito heart rate. J. Exp. Biol. 2020. In Revision. [Google Scholar]
- Broderick, K.E.; Feala, J.; McCulloch, A.; Paternostro, G.; Sharma, V.S.; Pilz, R.B.; Boss, G.R. The nitric oxide scavenger cobinamide profoundly improves survival in a Drosophila melanogaster model of bacterial sepsis. FASEB J. 2006, 20, 1865–1873. [Google Scholar] [CrossRef]
- Bullerjahn, A.; Mentel, T.; Pfluger, H.J.; Stevenson, P.A. Nitric oxide: A co-modulator of efferent peptidergic neurosecretory cells including a unique octopaminergic neurone innervating locust heart. Cell Tissue Res. 2006, 325, 345–360. [Google Scholar] [CrossRef] [PubMed]
- da Silva, R.; da Silva, S.R.; Lange, A.B. The regulation of cardiac activity by nitric oxide (NO) in the Vietnamese stick insect, Baculum extradentatum. Cell. Signal. 2012, 24, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Sigle, L.T.; Hillyer, J.F. Mosquito hemocytes preferentially aggregate and phagocytose pathogens in the periostial regions of the heart that experience the most hemolymph flow. Dev. Comp. Immunol. 2016, 55, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillyer, J.F. Integrated immune and cardiovascular function in Pancrustacea: Lessons from the insects. Integr. Comp. Biol. 2015, 55, 843–855. [Google Scholar] [CrossRef] [Green Version]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef] [Green Version]
- Whitten, M.M.A.; Coates, C.J. Re-evaluation of insect melanogenesis research: Views from the dark side. Pigment Cell Melanoma Res. 2017, 30, 386–401. [Google Scholar] [CrossRef]
- Brown, L.D.; Thompson, G.A.; Hillyer, J.F. Transstadial transmission of larval hemocoelic infection negatively affects development and adult female longevity in the mosquito Anopheles gambiae. J. Invertebr. Pathol. 2018, 151, 21–31. [Google Scholar] [CrossRef]
- Carlson, J.S.; Short, S.M.; Anglero-Rodriguez, Y.I.; Dimopoulos, G. Larval exposure to bacteria modulates arbovirus infection and immune gene expression in adult Aedes aegypti. Dev. Comp. Immunol. 2020, 104, 103540. [Google Scholar] [CrossRef]
- Vargas, V.; Cime-Castillo, J.; Lanz-Mendoza, H. Immune priming with inactive dengue virus during the larval stage of Aedes aegypti protects against the infection in adult mosquitoes. Sci. Rep. 2020, 10, 6723. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Garcia, M.; Vargas, V.; Ramirez-Bello, I.; Hernandez-Martinez, G.; Lanz-Mendoza, H. Bacterial exposure at the larval stage induced sexual immune dimorphism and priming in adult Aedes aegypti mosquitoes. PLoS ONE 2015, 10, e0133240. [Google Scholar] [CrossRef] [Green Version]
- Kala, M.K.; Gunasekaran, K. Effect of Bacillus thuringiensis ssp. israelensis on the development of Plasmodium gallinaceum in Aedes aegypti (Diptera: Culicidae). Ann. Trop. Med. Parasitol. 1999, 93, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, N.; Hazra, R.K.; Rup, S.; Acharya, A.S.; Dash, A.P. Bacillus sphaericus interferes with the development of Brugia malayi in Aedes aegypti. J. Helminthol. 1999, 73, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Paily, K.P.; Geetha, I.; Kumar, B.A.; Balaraman, K. Bacillus sphaericus in the adults of Culex quinquefasciatus mosquitoes emerged from treated larvae and its effect on development of the filarial parasite, Wuchereria bancrofti. Parasitol. Res. 2012, 110, 2229–2235. [Google Scholar] [CrossRef] [PubMed]
- League, G.P.; Hillyer, J.F. Functional integration of the circulatory, immune, and respiratory systems in mosquito larvae: Pathogen killing in the hemocyte-rich tracheal tufts. BMC Biol. 2016, 14, 78. [Google Scholar] [CrossRef] [Green Version]
- Babcock, D.T.; Brock, A.R.; Fish, G.S.; Wang, Y.; Perrin, L.; Krasnow, M.A.; Galko, M.J. Circulating blood cells function as a surveillance system for damaged tissue in Drosophila larvae. Proc. Natl. Acad. Sci. USA 2008, 105, 10017–10022. [Google Scholar] [CrossRef] [Green Version]
- Markus, R.; Laurinyecz, B.; Kurucz, E.; Honti, V.; Bajusz, I.; Sipos, B.; Somogyi, K.; Kronhamn, J.; Hultmark, D.; Ando, I. Sessile hemocytes as a hematopoietic compartment in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2009, 106, 4805–4809. [Google Scholar] [CrossRef] [Green Version]
- Sigle, L.T.; Hillyer, J.F. Mosquito hemocytes associate with circulatory structures that support intracardiac retrograde hemolymph flow. Front. Physiol. 2018, 9, 1187. [Google Scholar] [CrossRef]
- Williams, M.J.; Wiklund, M.L.; Wikman, S.; Hultmark, D. Rac1 signalling in the Drosophila larval cellular immune response. J. Cell Sci. 2006, 119, 2015–2024. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | VectorBase ID 1 | Primer sequences: 5’→3’ | Amplicon (bp) 2,3 | ||
|---|---|---|---|---|---|
| Forward | Reverse | Transcript | Genomic | ||
| RPS7 | AGAP010592 | GACGGATCCCAGCTGATAAA | GTTCTCTGGGAATTCGAACG | 132 | 281 |
| RPS17 | AGAP004887 | GACGAAACCACTGCGTAACA | TGCTCCAGTGCTGAAACATC | 153 | 264 |
| PPO6 | AGAP004977 | AGAGCCACTACCGGAAGGAT | TCGATGCTCTCAGCAATACG | 174 | 242 |
| NOS | AGAP029502 | CAAGAGTGGGACCACATCAA | ACCCTTCTGGACCATCTCCT | 129 | 210 |
| LYSC1 | AGAP007347 | ACGGCATCTTCCAGATCAAC | CATTGCAGTGGTTCTTCCAG | 180 | 259 |
| CEC1 | AGAP000693 | GCTGAAGAAGCTGGGAAAGA | ATGTTAGCAGAGCCGTCGTC | 158 | 247 |
| TEP1 | AGAP010815 | GACGTCCAAATACGGATCTCA | CTTTCAGGCATCACCCGTAT | 184 | NA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Powers, J.C.; Turangan, R.; Joosse, B.A.; Hillyer, J.F. Adult Mosquitoes Infected with Bacteria Early in Life Have Stronger Antimicrobial Responses and More Hemocytes after Reinfection Later in Life. Insects 2020, 11, 331. https://doi.org/10.3390/insects11060331
Powers JC, Turangan R, Joosse BA, Hillyer JF. Adult Mosquitoes Infected with Bacteria Early in Life Have Stronger Antimicrobial Responses and More Hemocytes after Reinfection Later in Life. Insects. 2020; 11(6):331. https://doi.org/10.3390/insects11060331
Chicago/Turabian StylePowers, Joseph C., Raymar Turangan, Bryan A. Joosse, and Julián F. Hillyer. 2020. "Adult Mosquitoes Infected with Bacteria Early in Life Have Stronger Antimicrobial Responses and More Hemocytes after Reinfection Later in Life" Insects 11, no. 6: 331. https://doi.org/10.3390/insects11060331