Effects of Pyriproxyfen Exposure on Reproduction and Gene Expressions in Silkworm, Bombyx mori

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Strains
2.2. Chemicals
2.3. Sample Preparation
2.4. Histopathological Examination
2.5. Isolation of Total RNA
2.6. Quantitative RT-PCR (qRT-PCR)
2.7. Statistical Analysis
3. Results
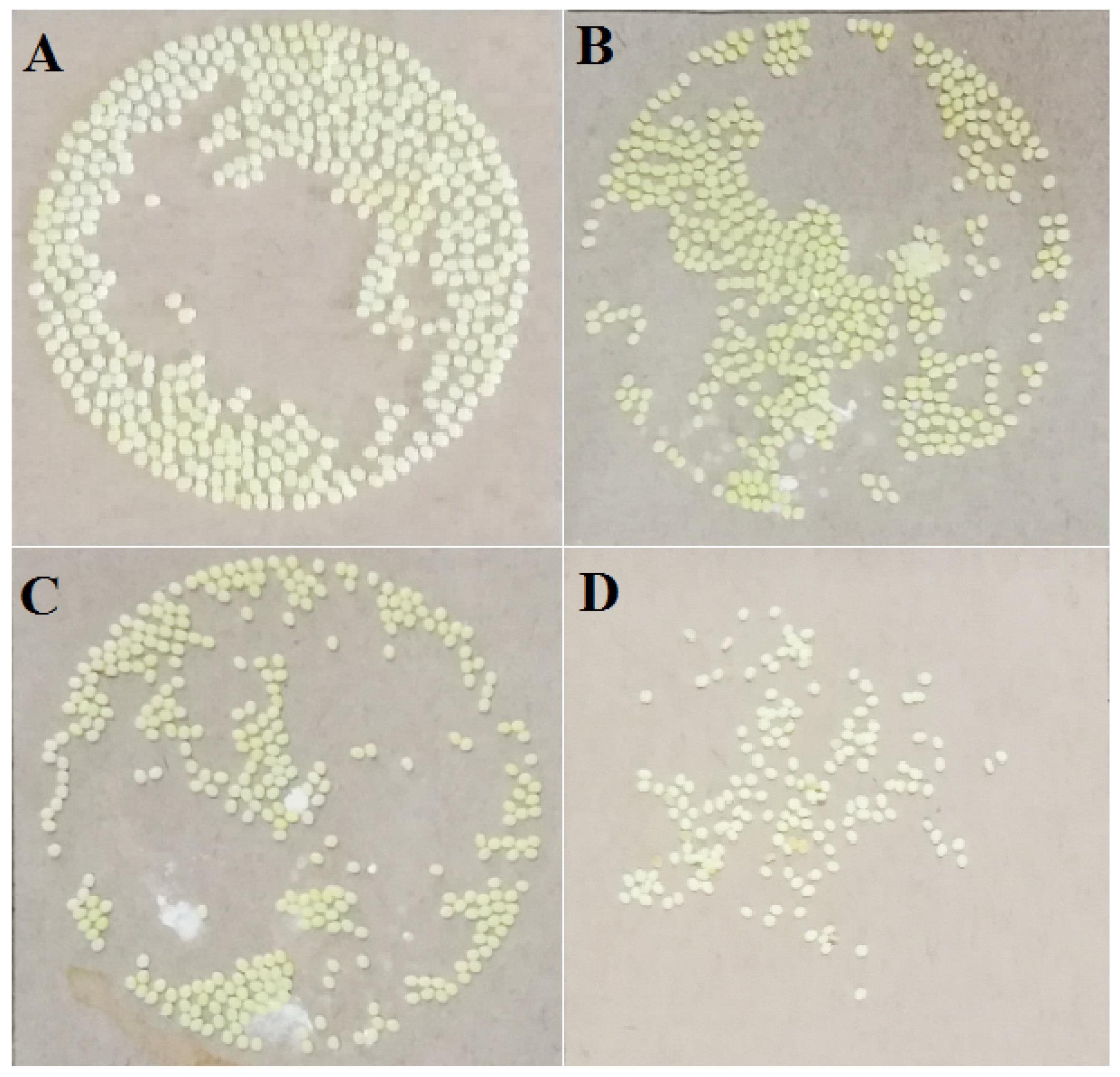
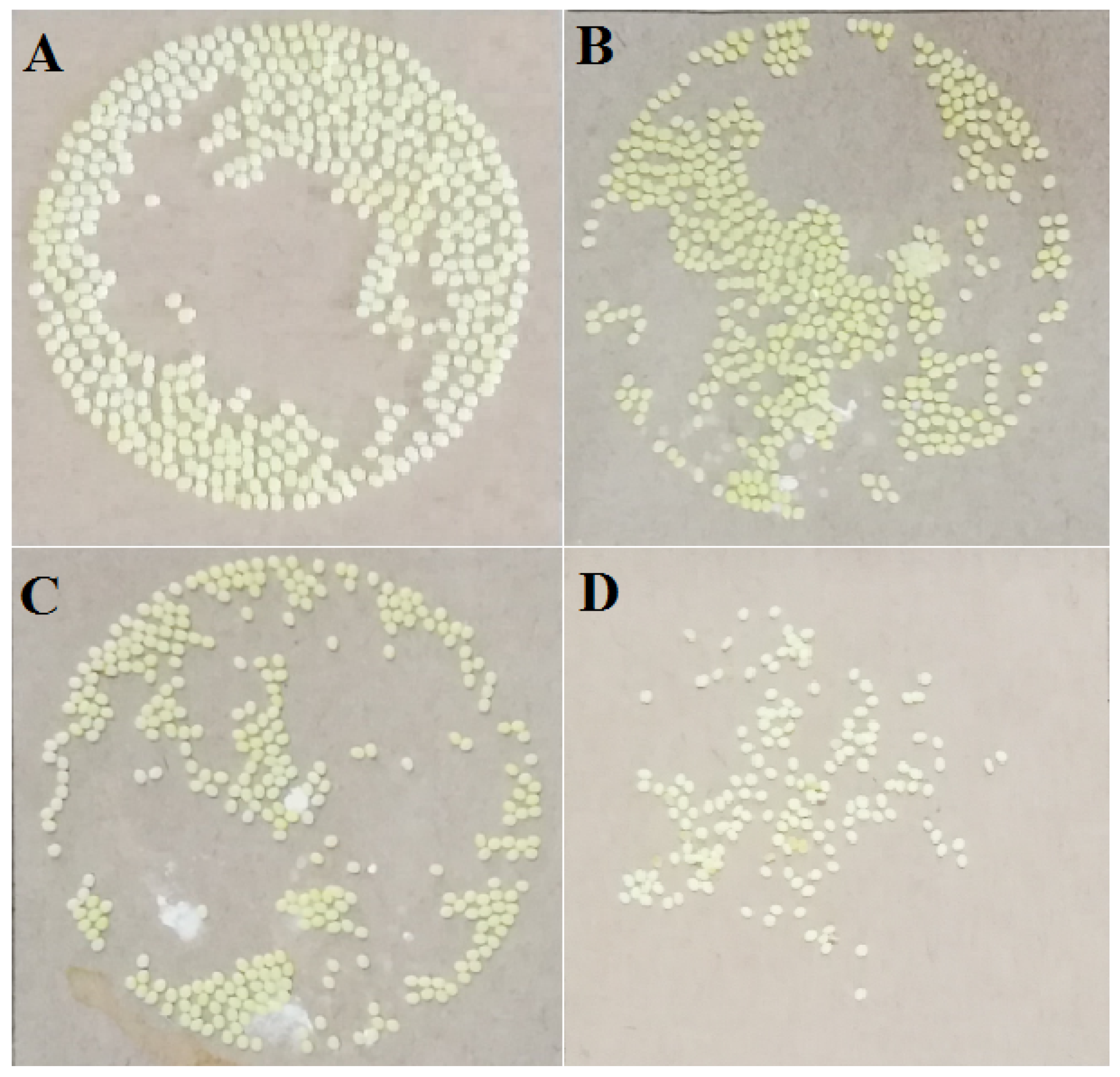
3.1. The Effects of Pyriproxyfen on the Oviposition of Silkworm at Larvae Stage
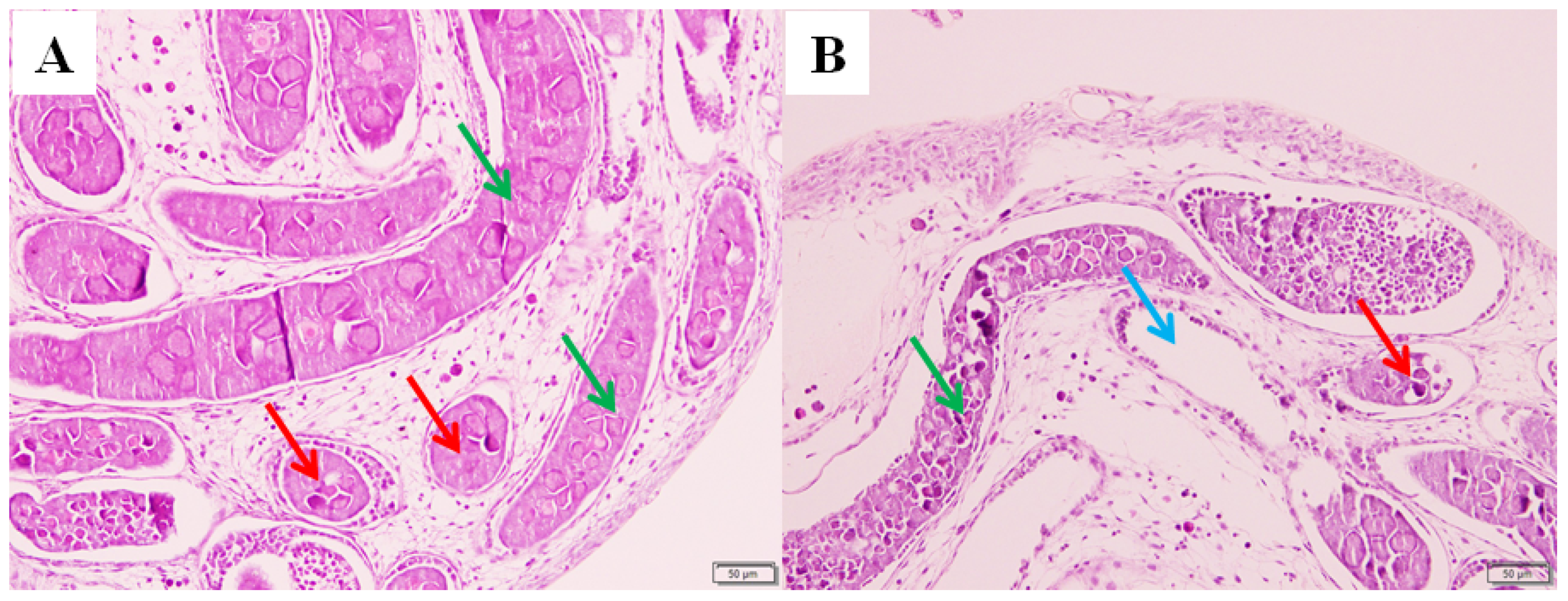
3.2. The Pathological Evaluation of the Ovary after Pyriproxyfen Exposure
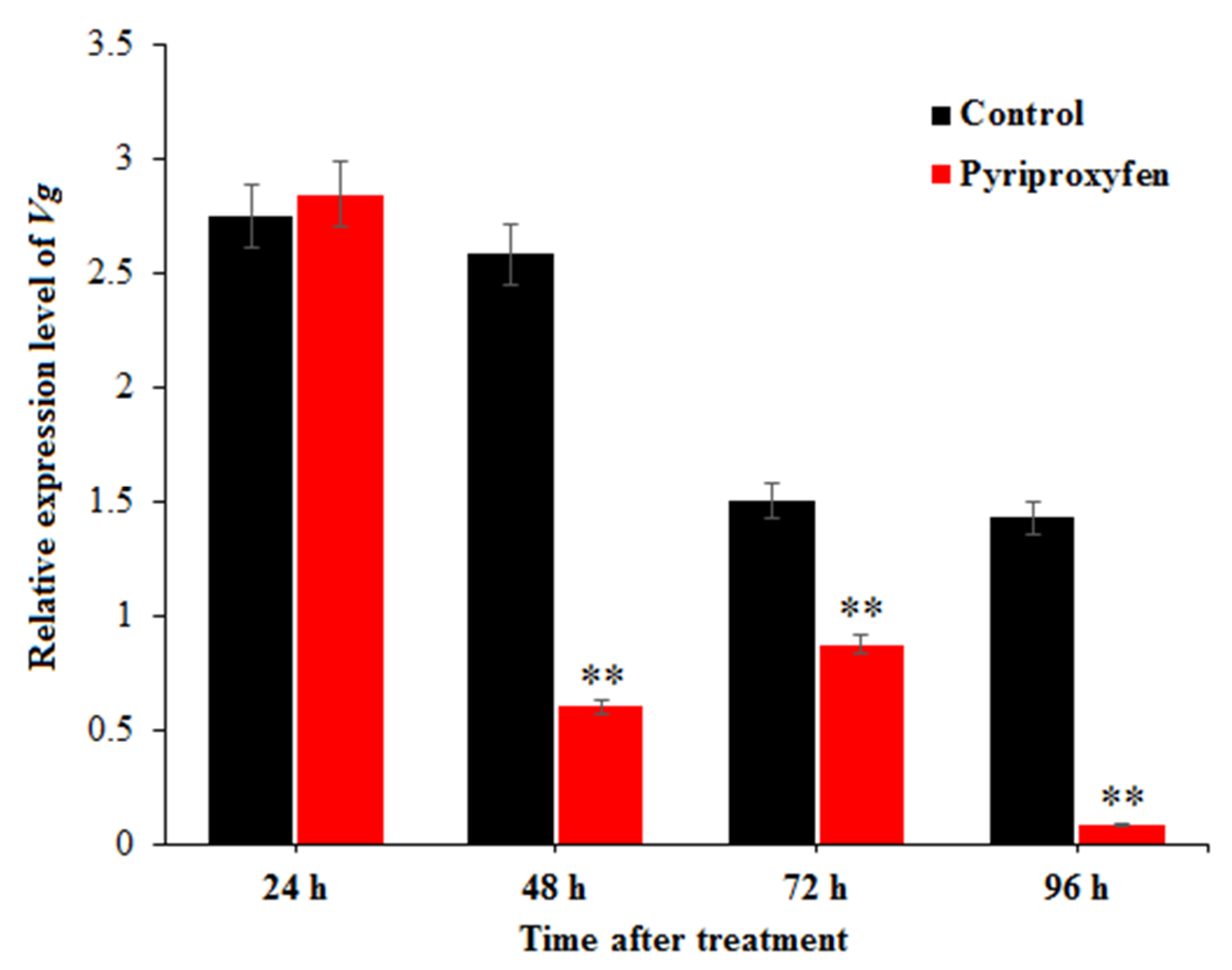
3.3. The Effect of Pyriproxyfen Exposure on the Expression Level of Vg in the Ovary
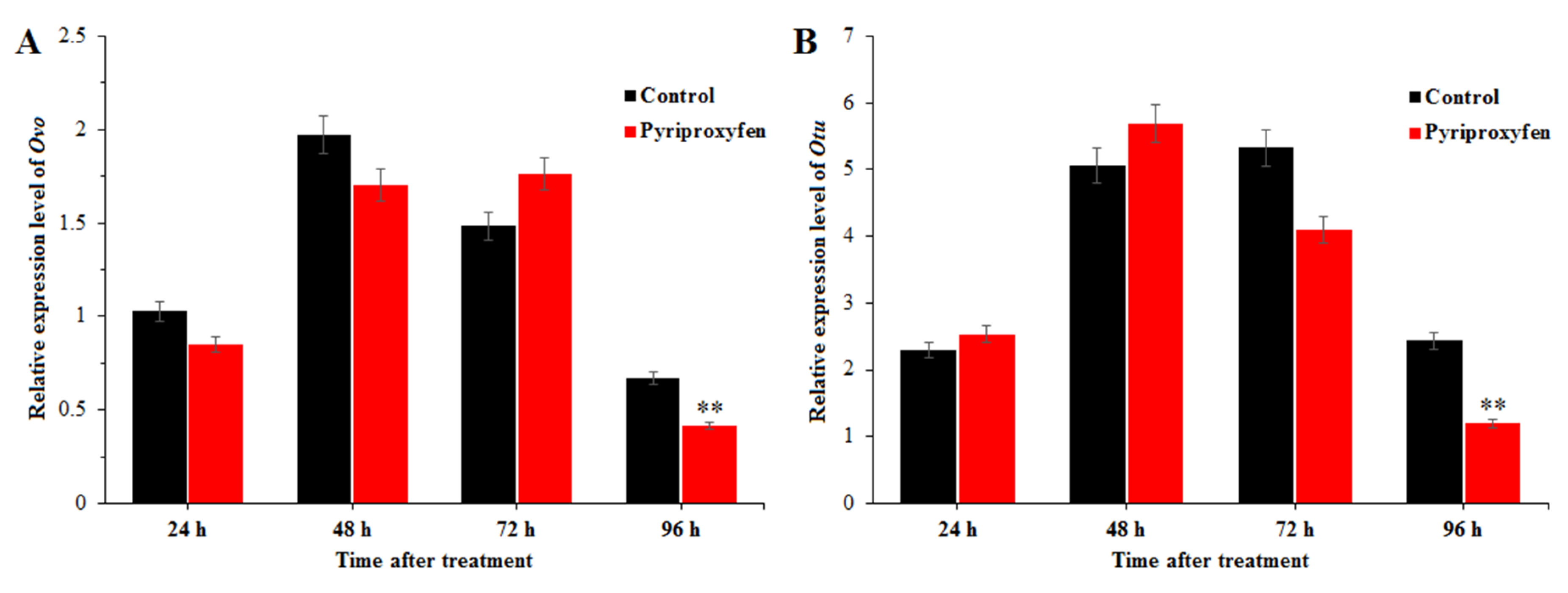
3.4. Transcriptional Analysis of Genes Related to Ovarian Development after Pyriproxyfen Exposure
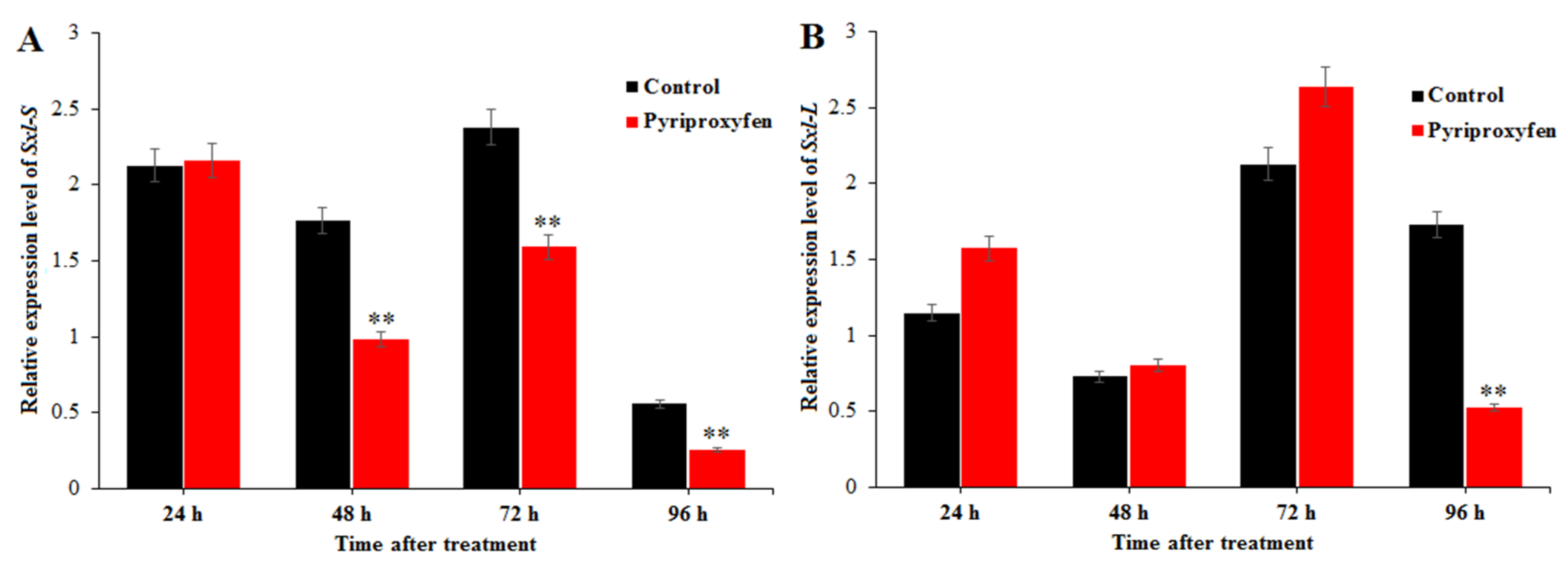
3.5. The Effect of Pyriproxyfen Exposure on the Expression Levels of Genes Related to Sex Differentiation in the Ovary
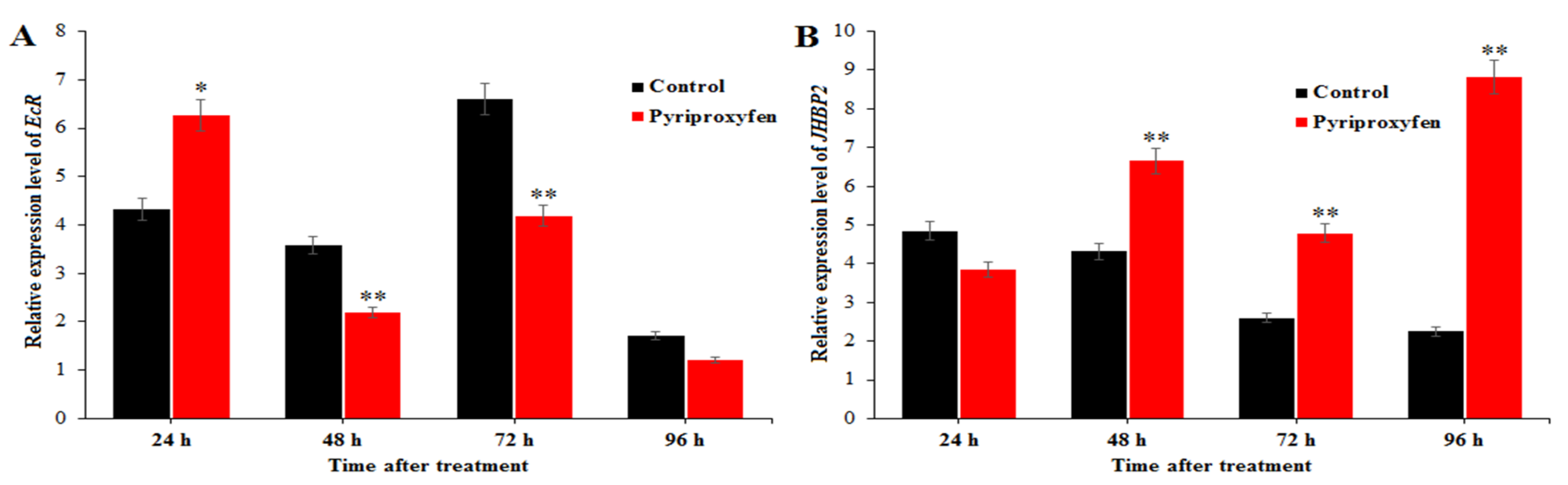
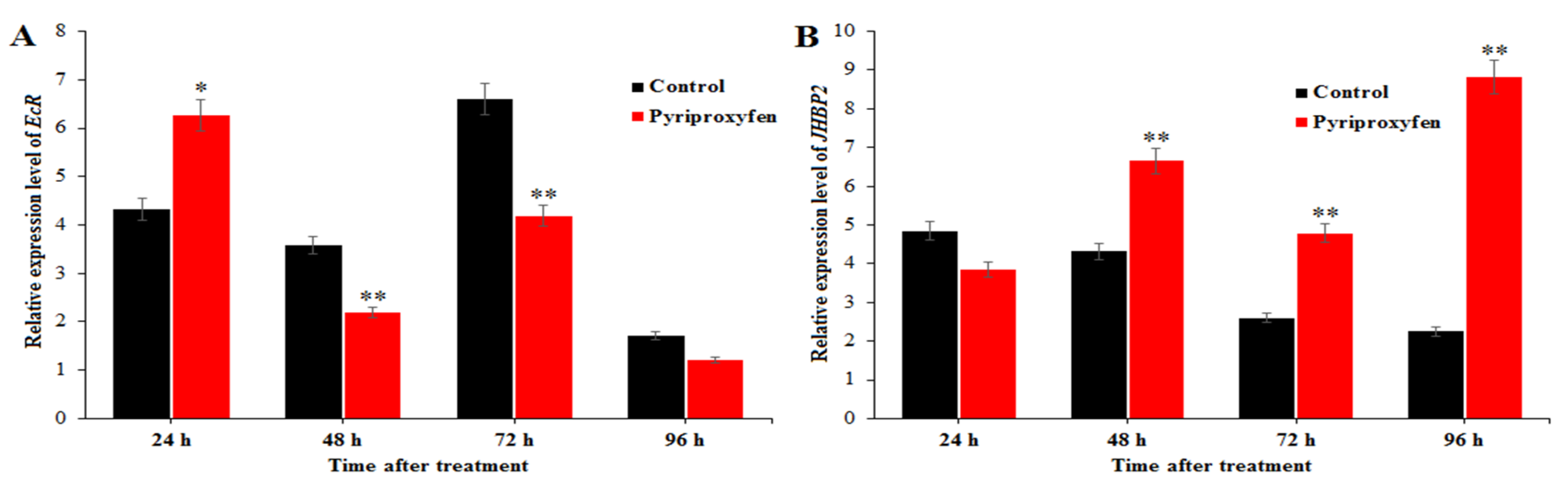
3.6. Changes in the Expression Levels of Genes Related to 20E and JH Regulation in Pyriproxyfen Treated Silkworms
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ishaaya, I.; Horowitz, A.R. Novel phenoxy juvenile hormone analog (pyriproxyfen) suppresses embryogenesis and adult emergence of sweetpotato whitefly (Homoptera: Aleyrodidae). J. Econ. Entomol. 1992, 85, 2113–2117. [Google Scholar] [CrossRef]
- Xu, J.T.; Sun, D.W.; Zhao, Q.C.; Shi, Q.; Shen, J.L. Study on the biological activity of chlorpyrine and mosquitin against nymph and egg of Bemisia tabaci. Acta Agric. Shanghai 2011, 27, 125–128. [Google Scholar]
- Ishaaya, I.; Horowitz, A.R. Pyriproxyfen, a novel insect growth regulator for controlling whiteflies: Mechanisms and resistance management. Pest Manag. Sci. 1995, 43, 227–232. [Google Scholar] [CrossRef]
- Magagula, C.N.; Samways, M.J. Effects of insect growth regulators on Chilacorus nigritus (Fabricius) (Coleoptera: Coccinellidae), a non-target natural enemy of citrus red scale, Aonidiella aurantii (Maskell) (Homoptera: Diaspididae), in southern Africa: Evidence from laboratory and field trials. Afr. Entomol. 2000, 8, 47–56. [Google Scholar]
- Chen, T.Y.; Liu, T.X. Susceptibility of immature stages of Chrysoperla rufilabris (Neurop. Chrysopidae) to pyriproxyfen, a juvenile hormone analog. J. Appl. Ent. 2002, 126, 125–129. [Google Scholar] [CrossRef]
- Tunaz, H.; Uygun, N. Insect growth regulators for insect. Turk. J. Agric. For. 2004, 28, 377–387. [Google Scholar]
- Lee, Y.S.; Lee, S.Y.; Park, E.C.; Kim, J.H.; Kim, G.H. Comparative Toxicities of Pyriproxyfen and Thiamethoxam against the Sweetpotato Whitefly, Bemisia tabaci (Homoptera: Aleyrodidae). J. Asia-Pac. Entomol. 2002, 5, 117–122. [Google Scholar] [CrossRef]
- Palumbo, J.C.; Horowitz, A.R.; Prabhaker, N. Insecticidal control and resistance management for Bemisia tabaci. Crop Prot. 2001, 20, 739–765. [Google Scholar] [CrossRef]
- Sun, H.Y.; Chen, W.G.; Dong, R.H.; Dai, J.Z. Toxicity test of 10% Pyriproxyfenr EC on silkworm. Bull. Seric. 2008, 39, 18–20. [Google Scholar]
- Zhang, Q.; Cui, X.Q.; Jiang, H.; Wang, X.J.; Wang, K.Y. Toxicity evaluation of insecticide mosquito fly ether on silkworm. Sci. Seric. 2011, 37, 176–180. [Google Scholar]
- Yang, C.W.; Lin, Y.; Liu, H.L.; Shen, G.W.; Luo, J.; Zhang, H.Y.; Peng, Z.X.; Chen, E.X.; Xing, R.M.; Han, C.S.; et al. The Broad Complex isoform 2 (BrC-Z2) transcriptional factor plays a critical role in vitellogenin transcription in the silkworm Bombyx mori. Biochim. Biophys. Acta 2014, 1840, 2674–2684. [Google Scholar] [CrossRef]
- Lin, Y.; Meng, Y.; Wang, Y.X.; Luo, J.; Katsuma, S.; Yang, C.W.; Banno, Y.; Kusakabe, T.; Shimada, T.; Xia, Q.Y. Vitellogenin Receptor Mutation Leads to the Oogenesis Mutant Phenotype “scanty vitellin” of the Silkworm, Bombyx mori. J. Biol. Chem. 2013, 288, 13345–13355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinson, S.; Nagoshi, R.N. Regulatory and functional interactions between the somatic sex regulatory gene transformer and the germline genes ovo and ovarian tumor. Development 1999, 126, 861–871. [Google Scholar] [PubMed]
- Xue, R.Y.; Hu, X.L.; Cao, G.L.; Huang, M.L.; Xue, G.X.; Qian, Y.; Song, Z.W.; Gong, C.L. Bmovo-1 Regulates Ovary Size in the Silkworm, Bombyx mori. PLoS ONE 2014, 9, e104928. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Spradling, A.C. Fusome asymmetry and oocyte determination in Drosophila. Dev. Gene 1995, 16, 6–12. [Google Scholar] [CrossRef]
- Rodesch, C.; Pettus, J.; Nagoshi, R.N. The Drosophilaovarian tumor Gene Is Required for the Organization of Actin Filaments during Multiple Stages in Oogenesis. Dev. Biol. 1997, 190, 153–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Q.H. Some advances in insect molecular biology sex determination, reproduction and hormone. Acta Entomol. 1993, 36, 115–127. [Google Scholar]
- Schütt, C.; Nöthiger, R. Structure, function and evolution of sex-determining systems in Dipteran insects. Development 2000, 127, 667–677. [Google Scholar] [PubMed]
- Goldsmith, M.R.; Shimada, T.; Abe, H. The genetics and genomics of the silkworm, Bombyx mori. Annu. Rev. Entomol. 2001, 50, 71–100. [Google Scholar] [CrossRef]
- White, B.H.; Ewer, J. Neural and hormonal control of postecdysial behaviors in insects. Annu. Rev. Entomol. 2014, 50, 363–381. [Google Scholar] [CrossRef] [Green Version]
- Weller, J.; Sun, G.C.; Zhou, B.; Lan, Q.; Hiruma, K.; Riddiford, L.M. Isolation and developmental expression of two nuclear receptors, MHR4 and beta FTZ-F1, in the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. Biol. 2001, 31, 827–837. [Google Scholar] [CrossRef]
- Xu, J.; Wang, Y.Q.; Li, Z.Q.; Ling, L.; Zeng, B.S.; You, L.; Chen, Y.Z.; Aslam, A.F.; Huang, Y.P.; Tan, A.J. Functional characterization of the vitellogenin promoter in the silkworm, Bombyx mori. Insect Mol. Biol. 2014, 23, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Bownes, M.; Scott, A.; Blair, M. The use of an inhibitor of protein synthesis to investigate the roles of ecdysteroids and sexdetermination genes on the expression of the genes encoding the Drosophila yolk proteins. Development 1987, 101, 931–941. [Google Scholar] [PubMed]
- Zhou, L.N. Effects of juvenile hormone on insect growth, development and reproduction. Shanghai Agric. Sci. Tech. 2017, 2, 131–133. [Google Scholar]
- Orth, A.P.; Tauchman, S.J.; Doll, S.C.; Goodman, W.G. Embryonic expression of juvenile hormone binding protein and its relationship to the toxic effects of juvenile hormone in Manduca sexta. Insect Biochem. Mol. Biol. 2003, 33, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.L.; Thummel, C.S. The RXR homolog ultraspiracle is an essential component of the Drosophila ecdysone receptor. Development 1998, 125, 4709–4717. [Google Scholar]
- Cheng, X.Y.; Hu, J.H.; Li, J.X.; Chen, J.; Wang, H.; Mao, T.T.; Xue, B.; Li, B. The silk gland damage and the transcriptional response to detoxifying enzymes-related genes of Bombyx mori under phoxim exposure. Chemosphere 2018, 209, 964–971. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wu, F.A.; Yu, M.D.; Cheng, J.L.; Tao, S.Q.; Wang, J. Determination of residual toxicity period of 40% LeSang emulsifiable oil, a new insect control drug in mulberry garden, to silkworm. Sci. Seric. 2005, 31, 471–473. [Google Scholar]
- Wang, Z.H.; Du, J.X.; Liang, M.Z.; Sun, R.Y.; Gong, L.M.; Hu, Y.L. Residual toxicity test of several commonly used insecticides in mulberry field on silkworm. Sci. Seric. 2002, 28, 146–150. [Google Scholar]
- Ponnuvel, K.M.; Nakazawa, H.; Furukawa, S.; Asaoka, A.; Ishibashi, J.; Tanaka, H.; Yamakawa, M. A lipase isolated from the silkworm Bombyx mori shows antiviral activity against nucleopolyhedro virus. J. Virol. 2003, 77, 10725–10729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begum, A.N.; Shivanandappa, T. Effect of sub-lethal dosage of insecticides on biological attributes of the silkworm, Bombyx mori L. (Lepidoptera: Bombycidae). Indian J. Seric. 2003, 42, 50–56. [Google Scholar]
- Rahman Khan, M.A.; Saha, B.N. Effect of Synthetic Pyrethroid on the Reproductive Potential of the Mulberry Silkworm, Bombyx mori L. Bangladesh J. Zool. 2005, 33, 117–119. [Google Scholar]
- Rahman Khan, M.A.; Saha, B.N. Growth and development of the mulberry silkworm, Bombyx mori L. on feed supplemented with alanine and glutamine. Sericologia 1995, 35, 657–666. [Google Scholar]
- Matozzo, V.; Marin, M.G. Can 4-nonylphenol induce vitellogenin-like proteins in the clam Tapes philippinarum? Environ. Res. 2005, 97, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.W.; Lin, Y.; Shen, G.W.; Chen, E.X.; Wang, Y.X.; Luo, J.; Zhang, H.Y.; Xing, R.M.; Xia, Q.Y. Female qualities in males: Vitellogenin synthesis induced by ovary transplants into the male silkworm, Bombyx mori. Biochem. Biophys. Res. Commun. 2014, 453, 31–36. [Google Scholar] [CrossRef]
- Xue, G.X. Study on Ovo and Gene of Silkworm. Master’s Thesis, Suzhou University, Suzhou, China, 2011; pp. 78–100. [Google Scholar]
- Sahut-Barnola, I.; Pauli, D. The Drosophila gene stand still encodes a germline chromatin-associated protein that controls the transcription of the ovarian tumor gene. Development 1999, 126, 1917–1926. [Google Scholar]
- Oliver, B.; Kim, Y.J.; Baker, B.S. Sex-lethal, master and slave: A hierarchy of germ-line sex determination in Drosophila. Development 1993, 119, 897–908. [Google Scholar]
- Eystathioy, T.; Swevers, L.; Iatrou, K. The orphan nuclear receptor BmHR3A of Bombyx mori: Hormonal control, ovarian expression and functional properties. Mech. Dev. 2001, 103, 107–115. [Google Scholar] [CrossRef]
- He, X.T.; Zhang, Y.N.; Li, F.; Li, G.Q.; Dong, S.L. Cloning, temporal expression of a juvenile hormone binding protein gene in Laodelphax striatellus (Fallén). J. Nanjing Agric. Univ. 2012, 35, 59–64. [Google Scholar]
- Suzuki, R.; Fujimoto, Z.; Shiotsuki, T.; Tsuchiya, W.; Momma, M.; Tase, A.; Miyazawa, M.; Yamazaki, T. Structural mechanism of JH delivery in hemolymph by JHBP of silkworm, Bombyx mori. Sci. Rep. 2011, 1, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5’–3’) |
|---|---|
| GAPDH | F: TGTTGAGGGCTTGATGAC |
| R: ACCTTACCCACAGCTTTG | |
| Vg | F: CTGCAACGCAAGGAAACCAA |
| R: TGGCCGTACTTGAAGTGCAT | |
| Ovo | F: GCAGCTGCTTTAGGACTACCAG |
| R: CGTTAGCTTCAGTCGCCAAA | |
| Otu | F: AACCACAACGCTGACCAGAA |
| R: GTGGCCCTTGTTCTGATGGT | |
| Sxl-S | F: CGCGTTACCTATTTAACATTTCGTG |
| R: CCTCGGTACTGCTGTTGGAT | |
| Sxl-L | F: CGGGATACTTGTTTGGTGGC |
| R: AGACATGCTGCCCCAGTATC | |
| EcR | F: TGATGGAGCAGAACAGGCAG |
| R: CCTCTTCATCCGACTGCGTT | |
| JHBP2 | F: CAATGCCTTAGCAGTGCGAC |
| R: TGAAGCGTATCACGACTCCC |
| Concentration of Solution/(μg/L) | Average Number of Eggs/(Grain) | Average Number of Hatching Eggs/(Grain) | Hatching Rate (%) | Percentage of Fertilized Eggs (%) |
|---|---|---|---|---|
| 0 (Control) | 459 ± 18 | 457 ± 19 | 99.56 ± 5.32 | 100 ± 0.00 |
| 0.001 | 408 ± 13 * | 387 ± 16 * | 94.85 ± 4.38 * | 96.81 ± 4.96 * |
| 1 | 370 ± 15 ** | 312 ± 13 ** | 84.32 ± 2.15 ** | 88.92 ± 2.66 * |
| 100 | 142 ± 8 ** | 45 ± 4 ** | 31.69 ± 0.86 ** | 45.77 ± 1.08 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, H.-Y.; Zhang, X.; Zhao, G.-D.; Guo, H.-M.; Li, G.; Xu, A.-Y. Effects of Pyriproxyfen Exposure on Reproduction and Gene Expressions in Silkworm, Bombyx mori. Insects 2020, 11, 467. https://doi.org/10.3390/insects11080467
Qian H-Y, Zhang X, Zhao G-D, Guo H-M, Li G, Xu A-Y. Effects of Pyriproxyfen Exposure on Reproduction and Gene Expressions in Silkworm, Bombyx mori. Insects. 2020; 11(8):467. https://doi.org/10.3390/insects11080467
Chicago/Turabian StyleQian, He-Ying, Xiao Zhang, Guo-Dong Zhao, Hui-Min Guo, Gang Li, and An-Ying Xu. 2020. "Effects of Pyriproxyfen Exposure on Reproduction and Gene Expressions in Silkworm, Bombyx mori" Insects 11, no. 8: 467. https://doi.org/10.3390/insects11080467
APA StyleQian, H.-Y., Zhang, X., Zhao, G.-D., Guo, H.-M., Li, G., & Xu, A.-Y. (2020). Effects of Pyriproxyfen Exposure on Reproduction and Gene Expressions in Silkworm, Bombyx mori. Insects, 11(8), 467. https://doi.org/10.3390/insects11080467