Essential Oils as Post-Harvest Crop Protectants against the Fruit Fly Drosophila suzukii: Bioactivity and Organoleptic Profile

 ,
, 
 ,
, 
 ,
,  , , and
, , and 
Simple Summary
Abstract
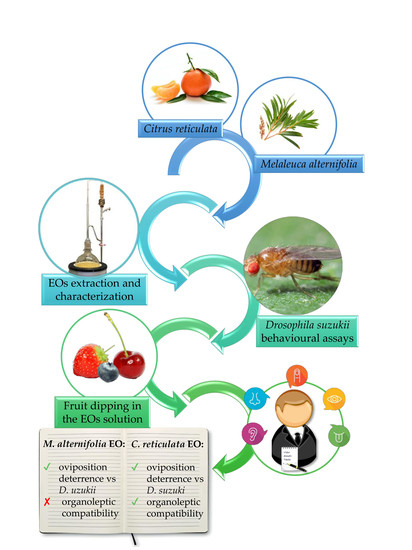
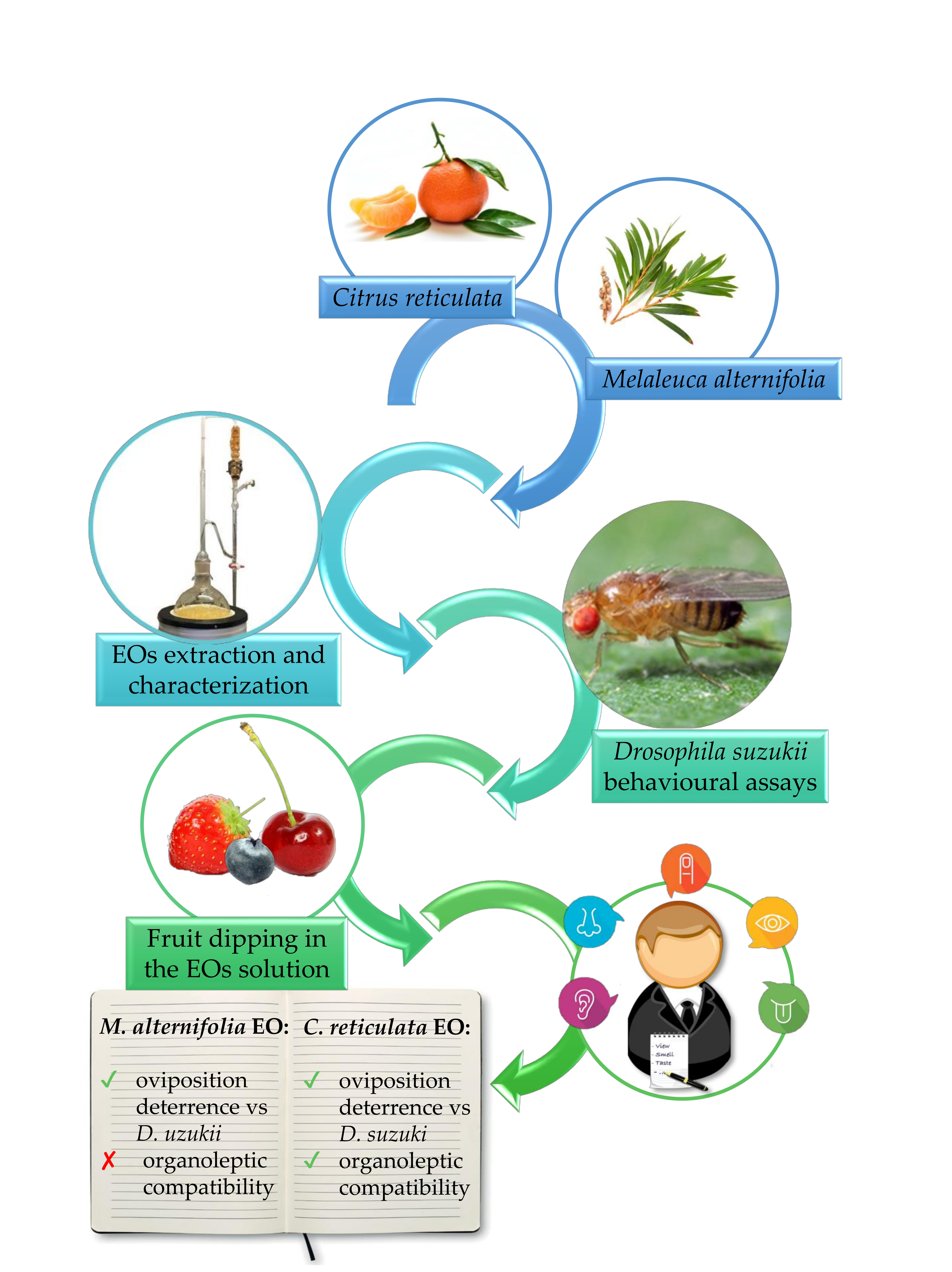
1. Introduction
2. Materials and Methods
2.1. Mass Rearing of Drosophila suzukii
2.2. Essential Oils Extraction and Chemical Analysis
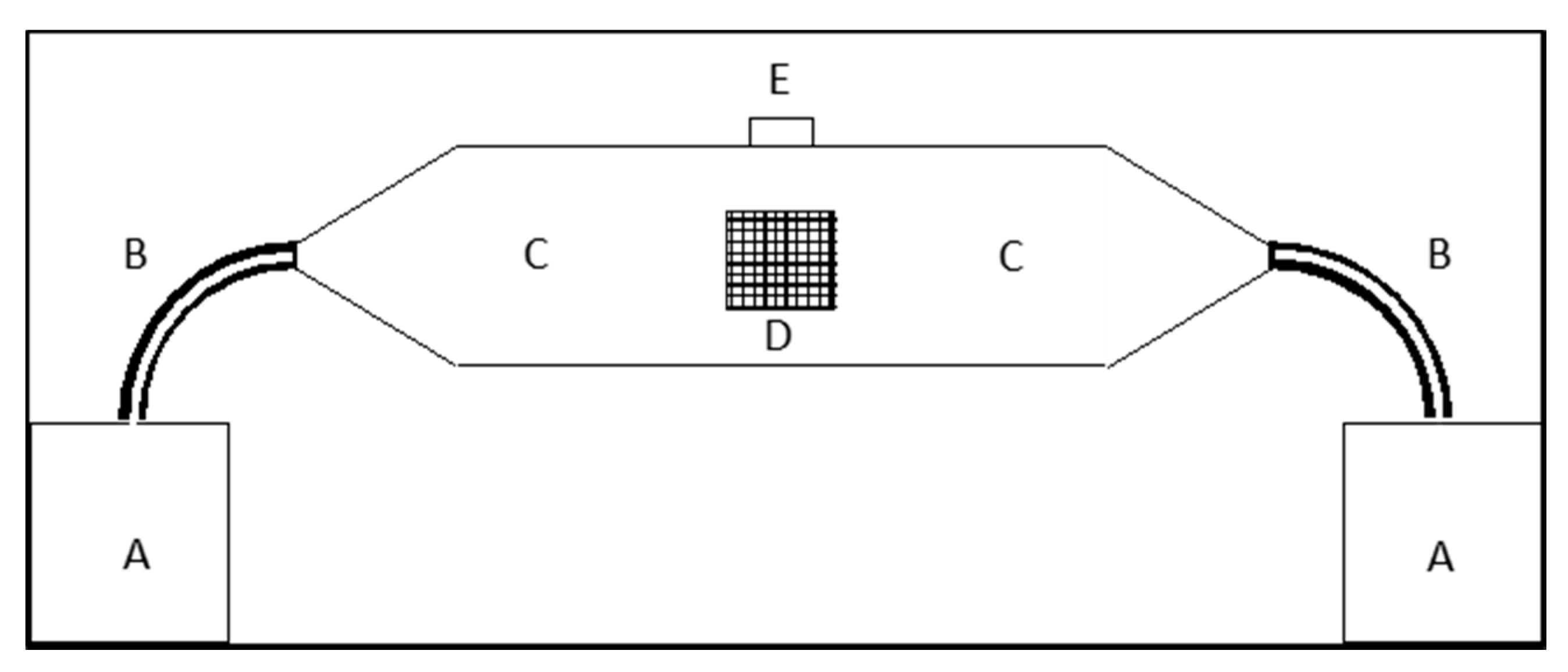
2.3. Behavioral Assays
- OD = oviposition deterrence;
- T = the number of eggs in the treated oviposition substrate;
- C = the number of eggs in the control oviposition substrate.
2.4. Organoleptic Characterization of EO-Treated Fruits
2.5. Statistical Analyses
3. Results
3.1. EOs Chemical Composition
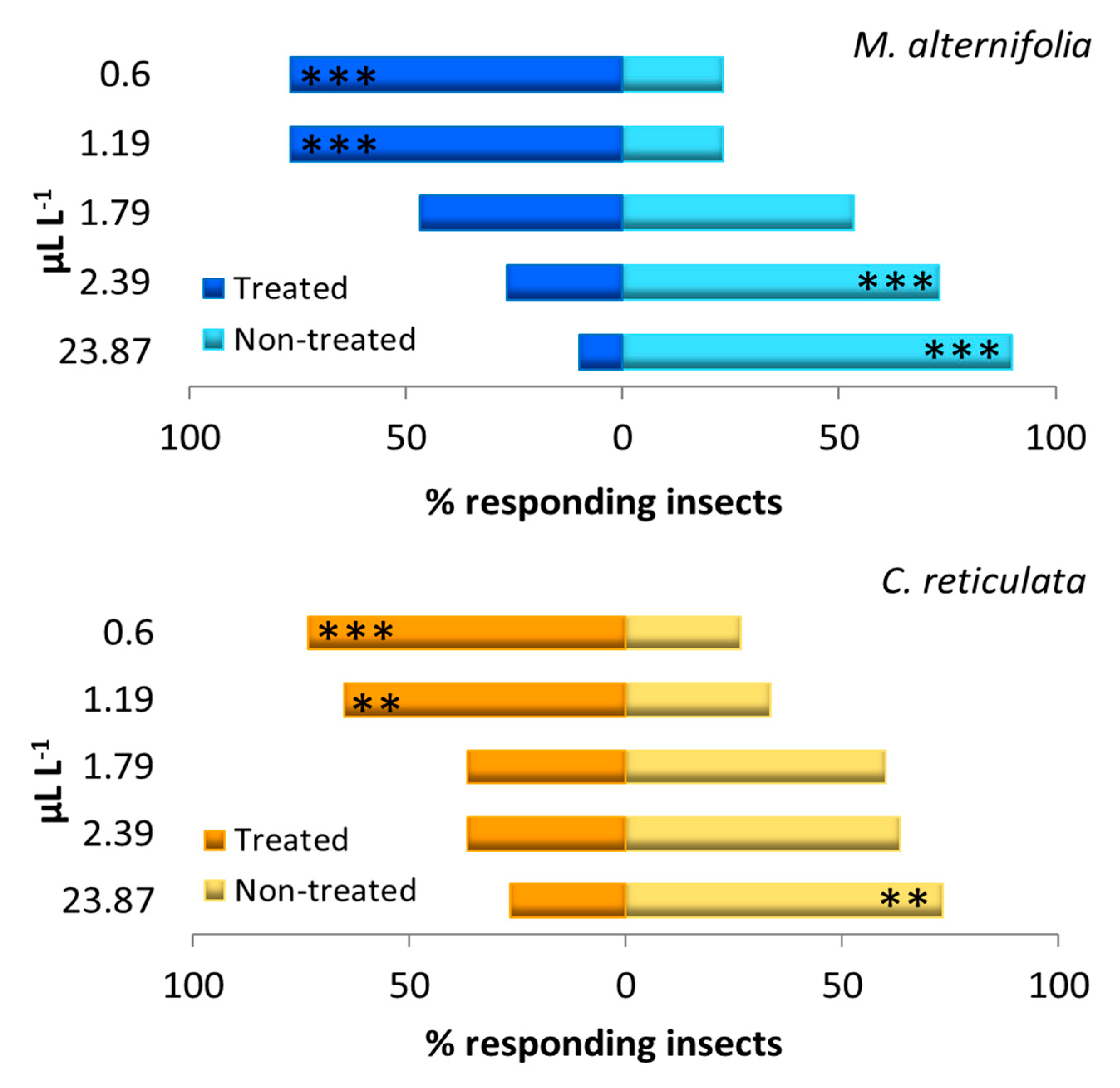
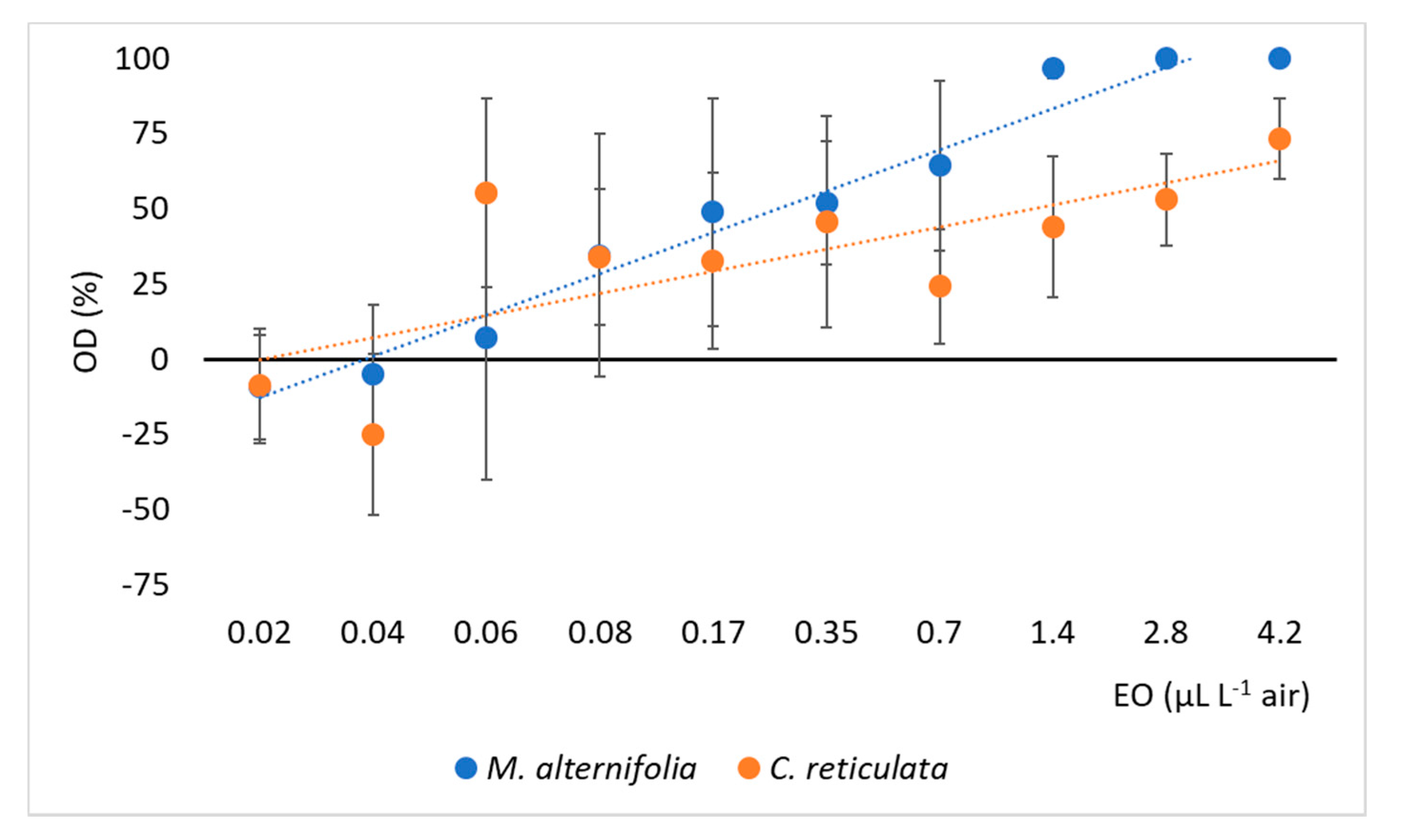
3.2. Behavioral Assays
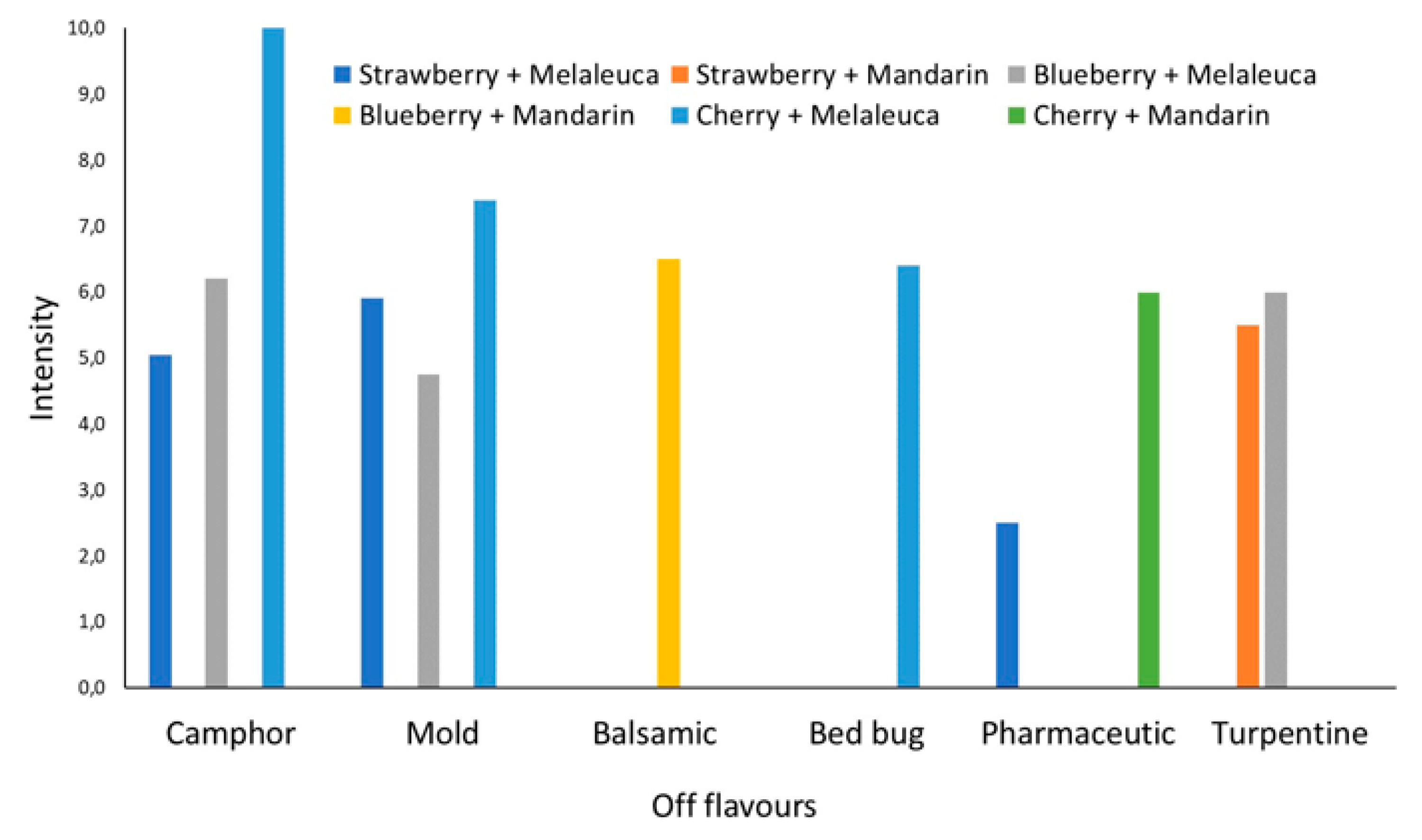
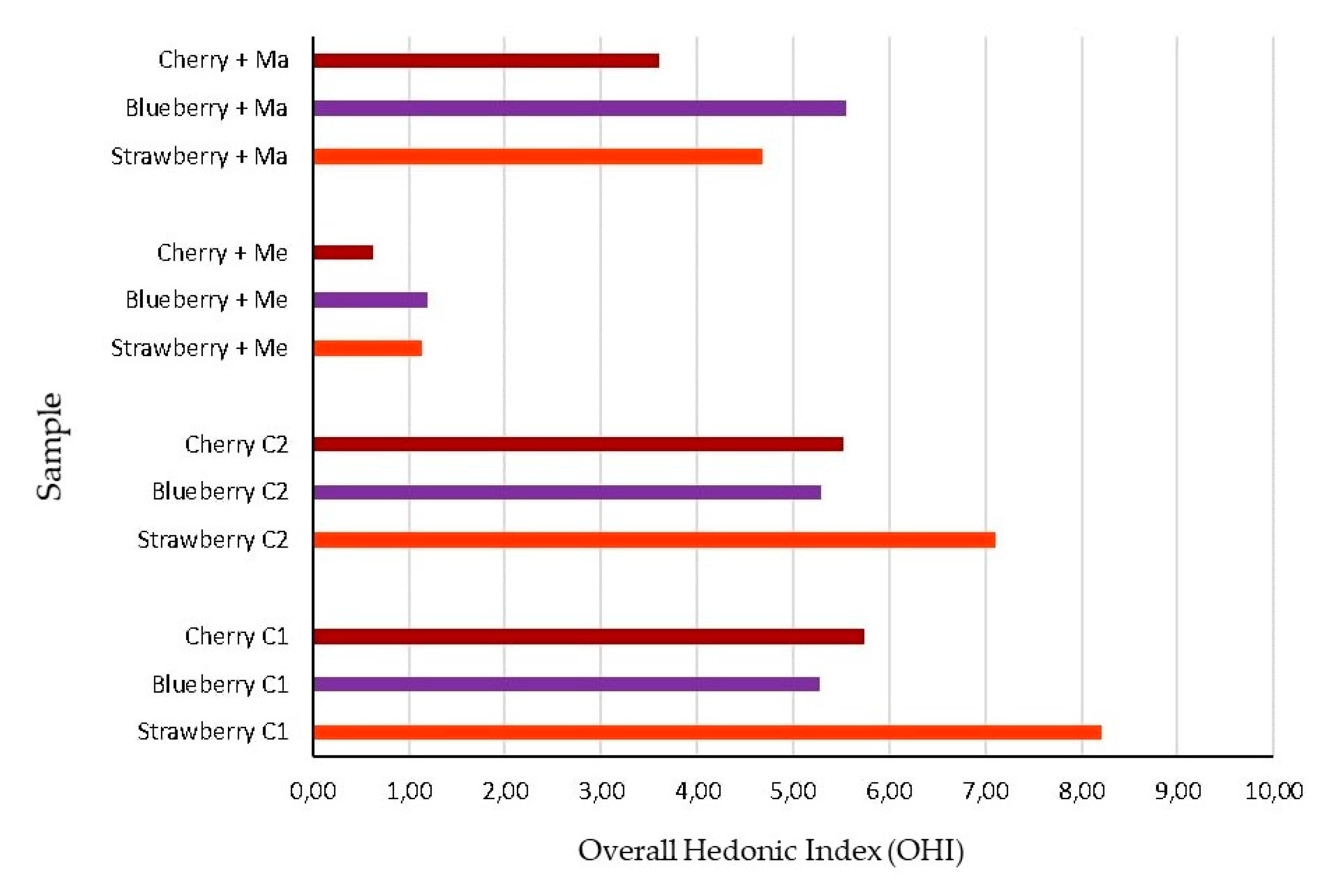
3.3. Sensorial Analysis of EOs Treated Fruit
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andreazza, F.; Bernardi, D.; dos Santos, R.S.S.; Garcia, F.R.M.; Oliveira, E.E.; Botton, M.; Nava, D.E. Drosophila suzukii in Southern Neotropical Region: Current Status and Future Perspectives. Neotrop. Entomol. 2017, 46, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. (2004) 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Calabria, G.; Máca, J.; Bächli, G.; Serra, L.; Pascual, M. First records of the potential pest species Drosophila suzukii (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectology 2012, 65, 149–160. [Google Scholar]
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef]
- Bruck, D.J.; Bolda, M.; Tanigoshi, L.; Klick, J.; Kleiber, J.; Defrancesco, J.; Gerdeman, B.; Spitler, H. Laboratory and field comparisons of insecticides to reduce infestation of Drosophila suzukii in berry crops. Pest Manag. Sci. 2011, 67, 1375–1385. [Google Scholar] [CrossRef]
- Mitsui, H.; Beppu, K.; Kimura, M.T. Seasonal life cycles and resource uses of flower- and fruit-feeding drosophilid flies (Diptera: Drosophilidae) in central Japan. Entomol. Sci. 2010, 13, 60–67. [Google Scholar] [CrossRef]
- Tait, G.; Grassi, A.; Pfab, F.; Crava, C.M.; Dalton, D.T.; Magarey, R.; Ometto, L.; Vezzulli, S.; Rossi-Stacconi, M.V.; Gottardello, A.; et al. Large-scale spatial dynamics of Drosophila suzukii in Trentino, Italy. J. Pest Sci. (2004) 2018, 91, 1213–1224. [Google Scholar] [CrossRef]
- Tonina, L.; Mori, N.; Giomi, F.; Battisti, A. Development of Drosophila suzukii at low temperatures in mountain areas. J. Pest Sci. 2016, 89, 667–678. [Google Scholar] [CrossRef]
- Bellamy, D.E.; Sisterson, M.S.; Walse, S.S. Quantifying Host Potentials: Indexing Postharvest Fresh Fruits for Spotted Wing Drosophila, Drosophila suzukii. PLoS ONE 2013, 8, e61227. [Google Scholar] [CrossRef] [PubMed]
- Burrack, H.J.; Fernandez, G.E.; Spivey, T.; Kraus, D.A. Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumara (Diptera: Drosophilidae), an invasive frugivore. Pest Manag. Sci. 2013, 69, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Ioriatti, C.; Walton, V.; Dalton, D.; Anfora, G.; Grassi, A.; Maistri, S.; Mazzoni, V. Drosophila suzukii (Diptera: Drosophilidae) and Its Potential Impact to Wine Grapes During Harvest in Two Cool Climate Wine Grape Production Regions. J. Econ. Entomol. 2015, 108, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Kenis, M.; Tonina, L.; Eschen, R.; van der Sluis, B.; Sancassani, M.; Mori, N.; Haye, T.; Helsen, H. Non-crop plants used as hosts by Drosophila suzukii in Europe. J. Pest Sci. 2016, 89, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Poyet, M.; Le Roux, V.; Gibert, P.; Meirland, A.; Prévost, G.; Eslin, P.; Chabrerie, O. The wide potential trophic niche of the asiatic fruit fly Drosophila suzukii: The key of its invasion success in temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef]
- Karageorgi, M.; Bräcker, L.B.; Lebreton, S.; Minervino, C.; Cavey, M.; Siju, K.P.; Grunwald Kadow, I.C.; Gompel, N.; Prud’homme, B. Evolution of Multiple Sensory Systems Drives Novel Egg-Laying Behavior in the Fruit Pest Drosophila suzukii. Curr. Biol. 2017, 27, 847–853. [Google Scholar] [CrossRef]
- Van Timmeren, S.; Isaacs, R. Control of spotted wing drosophila, Drosophila suzukii, by specific insecticides and by conventional and organic crop protection programs. Crop Prot. 2013, 54, 126–133. [Google Scholar] [CrossRef]
- Haye, T.; Girod, P.; Cuthbertson, A.G.S.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Baroffio, C.; Zhang, J.P.; Desneux, N. Current SWD IPM tactics and their practical implementation in fruit crops across different regions around the world. J. Pest Sci. 2016, 89, 643–651. [Google Scholar] [CrossRef]
- Burrack, H.J.; Asplen, M.; Bahder, L.; Collins, J.; Drummond, F.A.; Guédot, C.; Isaacs, R.; Johnson, D.; Blanton, A.; Lee, J.C.; et al. Multistate comparison of attractants for monitoring Drosophila suzukii (Diptera: Drosophilidae) in blueberries and caneberries. Environ. Entomol. 2015, 44, 704–712. [Google Scholar] [CrossRef]
- Mazzetto, F.; Gonella, E.; Crotti, E.; Vacchini, V.; Syrpas, M.; Pontini, M.; Mangelinckx, S.; Daffonchio, D.; Alma, A. Olfactory attraction of Drosophila suzukii by symbiotic acetic acid bacteria. J. Pest Sci. 2016, 89, 783–792. [Google Scholar] [CrossRef]
- Tonina, L.; Grassi, A.; Caruso, S.; Mori, N.; Gottardello, A.; Anfora, G.; Giomi, F.; Vaccari, G.; Ioriatti, C. Comparison of attractants for monitoring Drosophila suzukii in sweet cherry orchards in Italy. J. Appl. Entomol. 2018, 142, 18–25. [Google Scholar] [CrossRef]
- Grassi, A.; Anfora, G.; Maistri, S.; Gottardello, A.; Maddalena, G.; De Cristofaro, A.; Savini, G.; Ioriatti, C. Development and efficacy of Droskidrink, a food bait for trapping Drosophila suzukii. IOBC-WPRS Bull 2015, 109, 197–204. [Google Scholar]
- Wallingford, A.K.; Cha, D.H.; Loeb, G.M. Evaluating a push–pull strategy for management of Drosophila suzukii Matsumura in red raspberry. Pest Manag. Sci. 2018, 74, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Leach, H.; Moses, J.; Hanson, E.; Fanning, P.; Isaacs, R. Rapid harvest schedules and fruit removal as non-chemical approaches for managing spotted wing Drosophila. J. Pest Sci. (2004) 2018, 91, 219–226. [Google Scholar] [CrossRef]
- Rossi Stacconi, M.V.; Grassi, A.; Ioriatti, C.; Anfora, G. Augmentative releases of Trichopria drosophilae for the suppression of early season Drosophila suzukii populations. BioControl 2019, 64, 9–19. [Google Scholar] [CrossRef]
- Beers, E.H.; Van Steenwyk, R.A.; Shearer, P.W.; Coates, W.W.; Grant, J.A. Developing Drosophila suzukii management programs for sweet cherry in the western United States. Pest Manag. Sci. 2011, 67, 1386–1395. [Google Scholar] [CrossRef]
- Van Timmeren, S.; Mota-Sanchez, D.; Wise, J.C.; Isaacs, R. Baseline susceptibility of spotted wing Drosophila (Drosophila suzukii) to four key insecticide classes. Pest Manag. Sci. 2018, 74, 78–87. [Google Scholar] [CrossRef]
- Haddi, K.; Turchen, L.M.; Viteri Jumbo, L.O.; Guedes, R.N.C.; Pereira, E.J.G.; Aguiar, R.W.S.; Oliveira, E.E. Rethinking biorational insecticides for pest management: Unintended effects and consequences. Pest Manag. Sci. 2020, 76, 2286–2293. [Google Scholar] [CrossRef] [PubMed]
- Bedini, S.; Bougherra, H.H.; Flamini, G.; Cosci, F.; Belhamel, K.; Ascrizzi, R.; Conti, B. Repellency of anethole- and estragole-type fennel essential oils against stored grain pests: The different twins. Bull. Insectology 2016, 69, 149–157. [Google Scholar]
- Mossa, A.T.H. Green Pesticides: Essential oils as biopesticides in insect-pest management. J. Environ. Sci. Technol. 2016, 9, 354–378. [Google Scholar] [CrossRef]
- Conti, B.; Canale, A.; Cioni, P.L.; Flamini, G. Repellence of essential oils from tropical and Mediterranean Lamiaceae against Sitophilus zeamais. Bull. Insectology 2010, 63, 197–202. [Google Scholar]
- Erland, L.A.E.; Rheault, M.R.; Mahmoud, S.S. Insecticidal and oviposition deterrent effects of essential oils and their constituents against the invasive pest Drosophila suzukii (Matsumura) (Diptera: Drosophilidae). Crop Prot. 2015, 78, 20–26. [Google Scholar] [CrossRef]
- Park, C.G.; Jang, M.; Shin, E.; Kim, J. Myrtaceae plant essential oils and their β-triketone components as insecticides against Drosophila suzukii. Molecules 2017, 22, 1050. [Google Scholar] [CrossRef] [PubMed]
- Renkema, J.M.; Wright, D.; Buitenhuis, R.; Hallett, R.H. Plant essential oils and potassium metabisulfite as repellents for Drosophila suzukii (Diptera: Drosophilidae). Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jang, M.; Shin, E.; Kim, J.; Lee, S.H.; Park, C.G. Fumigant and contact toxicity of 22 wooden essential oils and their major components against Drosophila suzukii (Diptera: Drosophilidae). Pestic. Biochem. Physiol. 2016, 133, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Park, C.G.; Jang, M.; Yoon, K.A.; Kim, J. Insecticidal and acetylcholinesterase inhibitory activities of Lamiaceae plant essential oils and their major components against Drosophila suzukii (Diptera: Drosophilidae). Ind. Crops Prod. 2016, 89, 507–513. [Google Scholar] [CrossRef]
- Callander, J.T.; James, P.J. Insecticidal and repellent effects of tea tree (Melaleuca alternifolia) oil against Lucilia cuprina. Vet. Parasitol. 2012, 184, 271–278. [Google Scholar] [CrossRef]
- Benelli, G.; Canale, A.; Flamini, G.; Cioni, P.L.; Demi, F.; Ceccarini, L.; Macchia, M.; Conti, B. Biotoxicity of Melaleuca alternifolia (Myrtaceae) essential oil against the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae), and its parasitoid Psyttalia concolor (Hymenoptera: Braconidae). Ind. Crops Prod. 2013, 50, 596–603. [Google Scholar] [CrossRef]
- Conti, B.; Flamini, G.; Cioni, P.L.; Ceccarini, L.; Macchia, M.; Benelli, G. Mosquitocidal essential oils: Are they safe against non-target aquatic organisms? Parasitol. Res. 2014, 113, 251–259. [Google Scholar] [CrossRef]
- Fouad, H.A.; da Camara, C.A.G. Chemical composition and bioactivity of peel oils from Citrus aurantiifolia and Citrus reticulata and enantiomers of their major constituent against Sitophilus zeamais (Coleoptera: Curculionidae). J. Stored Prod. Res. 2017, 73, 30–36. [Google Scholar] [CrossRef]
- Liu, L.; Zhou, Q. Olfactory response of female Bactrocera minax to chemical components of the preference host Citrus volatile oils. J. Asia. Pac. Entomol. 2016, 19, 637–642. [Google Scholar] [CrossRef]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Caprile, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef]
- Ascrizzi, R.; Taglieri, I.; Sgherri, C.; Flamini, G.; Macaluso, M.; Sanmartin, C.; Venturi, F.; Quartacci, M.F.; Pistelli, L.; Zinnai, A. Nutraceutical oils produced by olives and Citrus peel of Tuscany varieties as sources of functional ingredients. Molecules 2019, 24, 65. [Google Scholar] [CrossRef] [PubMed]
- Masada, Y. Analysis of Essential Oils by Gas Chromatography and Mass Spectrometry; Wiley and Sons.: New York, NY, USA, 1976. [Google Scholar]
- Adams, R.P. Identification of essential oil components by gas chromatography / mass spectroscopy Robert P. Adams. J. Am. Soc. Mass Spectrom. 1997, 8, 671–672. [Google Scholar] [CrossRef]
- Romani, R.; Bedini, S.; Salerno, G.; Ascrizzi, R.; Flamini, G.; Echeverria, M.C.; Farina, P.; Conti, B. Andean flora as a source of new repellents against insect pests: Behavioral, morphological and electrophysiological studies on Sitophilus zeamais (coleoptera: Curculionidae). Insects 2019, 10. [Google Scholar] [CrossRef]
- Wei, Y.; Wei, Y.; Xu, F.; Shao, X. The combined effects of tea tree oil and hot air treatment on the quality and sensory characteristics and decay of strawberry. Postharvest Biol. Technol. 2018, 136, 139–144. [Google Scholar] [CrossRef]
- Venturi, F.; Bartolini, S.; Sanmartin, C.; Orlando, M.; Taglieri, I.; Macaluso, M.; Lucchesini, M.; Trivellini, A.; Zinnai, A.; Mensuali, A. Potato Peels as a Source of Novel Green Extracts Suitable as Antioxidant Additives for Fresh-Cut Fruits. Appl. Sci. 2019, 9, 2431. [Google Scholar] [CrossRef]
- Venturi, F.; Sanmartin, C.; Taglieri, I.; Xiaoguo, Y.; Quartacci, M.F.; Sgherri, C.; Andrich, G.; Zinnai, A. A kinetic approach to describe the time evolution of red wine as a function of packaging conditions adopted: Influence of closure and storage position. Food Packag. Shelf Life 2017, 13, 44–48. [Google Scholar] [CrossRef]
- Venturi, F.; Sanmartin, C.; Taglieri, I.; Xiaoguo, Y.; Andrich, G.; Zinnai, A. The influence of packaging on the sensorial evolution of white wine as a function of the operating conditions adopted during storage. Agrochimica 2016, 60, 150–160. [Google Scholar] [CrossRef]
- Kemp, S.E.; Hort, J.; Hollowood, T. Descriptive Analysis in Sensory Evaluation, 1st ed.; Kemp, S.E., Hort, J., Hollowood, T., Eds.; John Wiley & Sons Ltd All: Hoboken, NJ, USA, 2018; ISBN 9781118991671. [Google Scholar]
- O’Mahony, M. Sensory Evaluation of Food; Marcel Dekker, INC.: New York, NY, USA, 1986; ISBN 0824773373. [Google Scholar]
- Sanmartin, C.; Taglieri, I.; Venturi, F.; Macaluso, M.; Zinnai, A.; Tavarini, S.; Botto, A.; Serra, A.; Conte, G.; Flamini, G.; et al. Flaxseed cake as a tool for the improvement of nutraceutical and sensorial features of sourdough bread. Foods 2020, 9, 204. [Google Scholar] [CrossRef]
- Fanciullino, A.L.; Tomi, F.; Luro, F.; Desjobert, J.M.; Casanova, J. Chemical variability of peel and leaf oils of mandarins. Flavour Fragr. J. 2006, 21, 359–367. [Google Scholar] [CrossRef]
- Njoroge, S.M.; Koaze, H.; Mwaniki, M.; Minh Tu, N.T.; Sawamura, M. Essential oils of Kenyan Citrus fruits: Volatile components of two varieties of mandarins (Citrus reticulata) and a tangelo (C. paradisi x C. tangerina). Flavour Fragr. J. 2005, 20, 74–79. [Google Scholar] [CrossRef]
- Mohamed, M.I.E.; Abdelgaleil, S.A.M. Chemical composition and insecticidal potential of essential oils from Egyptian plants against Sitophilus oryzae (L.) (Coleoptera: Curculionidae) and Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). Appl. Entomol. Zool. 2008, 43, 599–607. [Google Scholar] [CrossRef]
- Viuda-Martos, M.; Ruiz-Navajas, Y.; Fernández-López, J.; Pérez-Álvarez, J.A. Chemical composition of mandarin (C. reticulata l.), grapefruit (C. paradisi l.), lemon (C. limon l.) and orange (C. sinensis l.) essential oils. J. Essent. Oil-Bearing Plants 2009, 12, 236–243. [Google Scholar] [CrossRef]
- Wang, J. The essential oil chemotype of Citrus reticulata blanco peel. J. Chem. Pharm. Res. 2014, 6, 16–18. [Google Scholar]
- Penfold, A.R.; Morrison, F.R.; Mckern, H.H.G.; Willis, J. Studies in the physiological forms of the Myrtaceae. Part II: The occurrence of physiological forms in Melaleuca alternifolia Cheel. Researches on Essential Oils of the Australian Flora. Mus. Technol. Appl. Sci. 1950, 2, 5–14. [Google Scholar]
- Butcher, P.A.; Doran, J.C.; Slee, M.U. Intraspecific variation in leaf oils of Melaleuca alternifolia (Myrtaceae). Biochem. Syst. Ecol. 1994, 22, 419–430. [Google Scholar] [CrossRef]
- Lee, L.S.; Brooks, L.O.; Homer, L.E.; Rossetto, M.; Henry, R.J.; Baverstock, P.R. Geographic variation in the essential oils and morphology of natural populations of Melaleuca alternifolia (Myrtaceae). Biochem. Syst. Ecol. 2002, 30, 343–360. [Google Scholar] [CrossRef]
- Keszei, A.; Hassan, Y.; Foley, W.J. A biochemical interpretation of terpene chemotypes in Melaleuca alternifolia. J. Chem. Ecol. 2010, 36, 652–661. [Google Scholar] [CrossRef]
- Padovan, A.; Keszei, A.; Hassan, Y.; Krause, S.T.; Köllner, T.G.; Degenhardt, J.; Gershenzon, J.; Külheim, C.; Foley, W.J. Four terpene synthases contribute to the generation of chemotypes in tea tree (Melaleuca alternifolia). BMC Plant Biol. 2017, 17, 1–14. [Google Scholar] [CrossRef]
- Shelton, D.; Aitken, K.; Doimo, L.; Leach, D.; Baverstock, P.; Henry, R. Genetic control of monoterpene composition in the essential oil of Melaleuca alternifolia (Cheel). Theor. Appl. Genet. 2002, 105, 377–383. [Google Scholar] [CrossRef]
- Bedini, S.; Flamini, G.; Cosci, F.; Ascrizzi, R.; Cioni, P.L.; Ioriatti, C.; Conti, B. Repellency of aromatic plants essential oils towards Drosophila suzukii. IOBC-WPRS Bull 2017, 128, 13–17. [Google Scholar]
- Klauck, V.; Pazinato, R.; Radavelli, W.M.; Volpato, A.; Stefani, L.M.; Santos, R.C.V.; Vaucher, R.A.; Boligon, A.A.; Athayde, M.L.; Da Silva, A.S. In vitro repellent effect of tea tree (Melaleuca alternifolia) and andiroba (Carapa guianensis) oils on Haemotobia irritans and Chrysomya megacephala flies. Trop. Biomed. 2015, 32, 160–166. [Google Scholar] [PubMed]
- Bilal, H.; Akram, W.; Hassan, S.A.; Zia, A.; Bhatti, A.R.; Mastoi, M.I.; Aslam, S. Insecticidal and repellent potential of Citrus essential oils against Tribolium castaneum Herbst (Coleoptera: Tenebrionidae). Pak. J. Zool. 2015, 47, 997–1002. [Google Scholar]
- Revadi, S.; Vitagliano, S.; Rossi Stacconi, M.V.; Ramasamy, S.; Mansourian, S.; Carlin, S.; Vrhovsek, U.; Becher, P.G.; Mazzoni, V.; Rota-Stabelli, O.; et al. Olfactory responses of Drosophila suzukii females to host plant volatiles. Physiol. Entomol. 2015, 40, 54–64. [Google Scholar] [CrossRef]
- Wallingford, A.K.; Hesler, S.P.; Cha, D.H.; Loeb, G.M. Behavioral response of spotted-wing drosophila, Drosophila suzukii Matsumura, to aversive odors and a potential oviposition deterrent in the field. Pest Manag. Sci. 2016, 72, 701–706. [Google Scholar] [CrossRef]
- Dweck, H.K.M.; Ebrahim, S.A.M.; Kromann, S.; Bown, D.; Hillbur, Y.; Sachse, S.; Hansson, B.S.; Stensmyr, M.C. Olfactory preference for egg laying on Citrus substrates in Drosophila. Curr. Biol. 2013, 23, 2472–2480. [Google Scholar] [CrossRef]
- Renkema, J.M.; Buitenhuis, R.; Hallett, R.H. Reduced Drosophila suzukii infestation in berries using deterrent compounds and laminate polymer flakes. Insects 2017, 8, 117. [Google Scholar] [CrossRef]
- Pathare, P.B.; Opara, U.L.; Al-Said, F.A.J. Colour Measurement and Analysis in Fresh and Processed Foods: A Review. Food Bioprocess Technol. 2013, 6, 36–60. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Smell | Taste | Touch | Hedonic Parameter |
|---|---|---|---|
| Intensity | Sweet | Hardness | Overall pleasantness |
| Typical fruit | Acid | Resistance to chewing | Frankness of smell |
| Floral | Salty | Elasticity | Fineness of smell |
| Vegetal | Bitter | Springiness | |
| Spicy | Fibrous behavior | ||
| Citrusy | Granular behavior |
| Chemical Compounds | l.r.i. | Citrus reticulata EO | Melaleuca alternifolia EO |
|---|---|---|---|
| tricyclene | 928 | 0.7 | - |
| α-thujene | 931 | tr | 0.3 |
| α-pinene | 941 | - | 3.2 |
| camphene | 954 | - | tr |
| sabinene | 976 | 3.0 | - |
| β-pinene | 982 | 0.1 | 0.7 |
| myrcene | 993 | 2.4 | 0.6 |
| octanal | 1001 | 0.8 | - |
| α-phellandrene | 1005 | - | 0.2 |
| α-terpinene | 1018 | 0.1 | 4.5 |
| p-cymene | 1027 | - | 3.1 |
| limonene | 1032 | 83.6 | tr |
| 1,8-cineole | 1034 | - | 22.4 |
| (E)-β-ocimene | 1052 | 0.3 | - |
| γ-terpinene | 1062 | 0.2 | 9.0 |
| 1-octanol | 1071 | 0.4 | - |
| terpinolene | 1089 | tr | 1.7 |
| linalool | 1101 | 6.0 | tr |
| cis-p-menth-2-en-1-ol | 1123 | tr | 0.2 |
| δ-terpineol | 1170 | - | 0.2 |
| 4-terpineol | 1178 | 0.6 | 17.7 |
| p-cymen-8-ol | 1183 | - | tr |
| α-terpineol | 1189 | 0.5 | 5.5 |
| cis-piperitol | 1195 | - | tr |
| decanal | 1204 | 0.5 | - |
| trans-piperitol | 1207 | - | tr |
| nerol | 1230 | 0.3 | - |
| perilla aldehyde | 1273 | 0.3 | - |
| carvacrol | 1298 | - | tr |
| α-cubebene | 1350 | - | tr |
| isoledene | 1374 | - | 0.2 |
| α-copaene | 1376 | tr | 0.2 |
| β-patchoulene | 1380 | - | 0.1 |
| α-gurjunene | 1410 | - | 0.9 |
| β-caryophyllene | 1420 | tr | 1.1 |
| β-gurjunene | 1432 | - | 0.2 |
| α-guaiene | 1439 | - | 0.3 |
| aromadendrene | 1441 | tr | 2.9 |
| α-humulene | 1456 | - | 0.3 |
| alloaromadendrene | 1461 | - | 1.2 |
| trans-cadina-1(6),4-diene | 1470 | - | 0.8 |
| γ-muurolene | 1477 | - | 0.1 |
| β-selinene | 1485 | - | 0.5 |
| δ-selinene | 1493 | - | 0.4 |
| viridiflorene | 1494 | - | 2.0 |
| α-muurolene | 1498 | - | 0.4 |
| δ-cadinene | 1524 | tr | 4.6 |
| cubenene | 1534 | - | 0.5 |
| α-calacorene | 1546 | - | tr |
| germacrene B | 1556 | 0.1 | - |
| ledol | 1566 | - | 0.7 |
| spathulenol | 1576 | - | 1.0 |
| globulol | 1583 | - | 3.1 |
| viridiflorol | 1590 | - | 1.1 |
| guaiol | 1595 | - | 1.0 |
| 1-epi-cubenol | 1628 | - | 1.8 |
| epi-α-cadinol | 1640 | - | 0.5 |
| cubenol | 1643 | - | 0.9 |
| selin-11-en-4-α-ol | 1653 | - | 0.2 |
| 14-hydroxy-9-epi-(E)-caryophyllene | 1664 | - | 0.2 |
| Monoterpene hydrocarbons | 90.4 | 23.2 | |
| Oxygenated monoterpenes | 7.7 | 45.9 | |
| Sesquiterpene hydrocarbons | 0.1 | 16.8 | |
| Oxygenated sesquiterpenes | - | 10.4 | |
| Non-terpene derivatives | 1.7 | - | |
| Total identified (%) | 99.9 | 96.3 | |
| Sample | L* | a* | b* |
|---|---|---|---|
| Control 1 | 13.30 a | 37.30 a | 28.70 a |
| Control 2 | 13.50 a | 39.00 b | 29.07 a |
| C. reticulata EO | 17.300 b | 39.10 b | 30.10 b |
| M. alternifolia EO | 22.13 c | 37.47 a | 35.50 c |
| p-Value 1 | Control 1 | Control 2 | C. reticulata EO | M. alternifolia EO | |
|---|---|---|---|---|---|
| Control 1 | *** | - | 1.85 c | 4.65 b | 11.18 a |
| Control 2 | *** | 1.85 c | - | 4.02 b | 10.94 a |
| Fruit | Parameter | p-Value 1 | Control 1 | Control 2 | M. alternifolia | C. reticulata |
|---|---|---|---|---|---|---|
| Strawberry | Smell attributes | |||||
| Intensity | ns | 6.23 | 6.10 | 7.45 | 7.08 | |
| Typical fruit | *** | 7.92 a | 6.03 ab | 2.85 b | 5.13 ab | |
| Floral | ns | 2.07 | 1.57 | 0.33 | 1.75 | |
| Vegetal | ns | 1.00 | 1.20 | 0.67 | 0.83 | |
| Spicy | ns | 0.58 | 0.83 | 1.42 | 0.42 | |
| Citrusy | *** | 0.33 b | 0.17 b | 0.08 b | 5.68 a | |
| Taste/Touch attributes | ||||||
| Sweet | *** | 3.47 a | 4.15 a | n.a. | 3.32 a | |
| Acid | *** | 6.80 a | 5.40 a | n.a. | 5.38 a | |
| Salty | * | 1.00 ab | 1.92 a | n.a. | 1.08 ab | |
| Bitter | ** | 1.32 a | 1.90 a | n.a. | 3.07 a | |
| Hardness | *** | 4.00 a | 3.37 a | n.a. | 2.53 a | |
| Resistance to chewing | ** | 3.47 a | 3.18 a | n.a. | 2.42 ab | |
| Elasticity | *** | 1.87 a | 1.35 a | n.a. | 1.25 a | |
| Springiness | *** | 5.33 a | 4.83 a | n.a. | 4.80 a | |
| Fibrous behavior | ** | 2.00 a | 1.25 a | n.a. | 1.42 a | |
| Granular behavior | ** | 2.62 a | 1.42 ab | n.a. | 2.00 ab | |
| Blueberry | Smell attributes | |||||
| Intensity | * | 1.70 b | 2.43 b | 6.67 a | 5.92 a | |
| Typical fruit | ns | 0.33 | 0.17 | 0.33 | 0.17 | |
| Floral | ns | 0.00 | 0.00 | 0.67 | 0.67 | |
| Vegetal | ns | 0.67 | 0.35 | 2.58 | 1.58 | |
| Spicy | ns | 0.00 | 0.00 | 1.83 | 0.50 | |
| Citrusy | *** | 0.00 b | 0.00 b | 0.17 b | 7.75 a | |
| Taste/Touch attributes | ||||||
| Sweet | *** | 3.83 a | 2.53 a | n.a. | 3.92 a | |
| Acid | *** | 3.42 ab | 5.42 a | n.a. | 4.45 a | |
| Salty | ** | 0.75 a | 1.17 a | n.a. | 0.83 a | |
| Bitter | * | 0.72 a | 0.55 a | n.a. | 2.35 a | |
| Hardness | *** | 1.78 a | 2.03 a | n.a. | 1.70 a | |
| Resistance to chewing | *** | 1.37 a | 1.75 a | n.a. | 1.65 a | |
| Elasticity | *** | 2.48 a | 2.08 a | n.a. | 2.23 a | |
| Springiness | *** | 4.78 a | 5.62 a | n.a. | 5.58 a | |
| Fibrous behavior | ** | 0.75 ab | 1.17 a | n.a. | 0.83 ab | |
| Granular behavior | ns | 0.50 | 0.67 | n.a. | 0.57 | |
| Cherry | Smell Attributes | |||||
| Intensity | *** | 1.45 b | 1.57 b | 7.05 a | 5.75 a | |
| Typical fruit | ns | 1.25 | 1.38 | 0.00 | 0.33 | |
| Floral | ns | 0.33 | 0.33 | 0.00 | 0.83 | |
| Vegetal | ns | 0.75 | 0.33 | 1.67 | 1.00 | |
| Spicy | * | 0.00 b | 0.00 b | 2.17 a | 1.33 ab | |
| Citrusy | *** | 0.12 b | 0.12 b | 0.18 b | 7.67a | |
| Taste/Touch attributes | ||||||
| Sweet | *** | 4.67 a | 3.92 a | n.a. | 3.42 a | |
| Acid | *** | 4.37 a | 4.87 a | n.a. | 6.32 a | |
| Salty | * | 0.83 a | 1.08 a | n.a. | 0.67 a | |
| Bitter | *** | 1.33 b | 1.50 b | n.a. | 4.90 a | |
| Hardness | *** | 3.42 a | 3.50 a | n.a. | 3.03 a | |
| Resistance to chewing | *** | 3.42 a | 4.25 a | n.a. | 3.02 a | |
| Elasticity | *** | 3.15 a | 2.92 a | n.a. | 3.18 a | |
| Springiness | *** | 4.83 a | 6.07 a | n.a. | 6.27 a | |
| Fibrous behavior | *** | 2.50 a | 2.75 a | n.a. | 2.75 a | |
| Granular behavior | ns | 1.07 | 1.32 | n.a. | 1.00 | |
| Fruit | Hedonic Parameters | p-Value 1 | Control 1 | Control 2 | M. alternifolia | C. reticulata |
|---|---|---|---|---|---|---|
| Strawberry | Overall pleasantness | *** | 7.08 a | 5.28 a | 0.83 c | 3.22 b |
| Frankness of smell | *** | 7.97 a | 7.33 a | 1.12 c | 5.17 b | |
| Fineness of smell | *** | 7.13 a | 6.57 a | 1.12 c | 4.25 b | |
| Blueberry | Overall pleasantness | *** | 5.95 a | 4.83 a | 1.00 b | 5.67 a |
| Frankness of smell | *** | 7.58 ab | 8.33 a | 1.17 c | 5.67 b | |
| Fineness of smell | * | 0.72 b | 1.13 a | 1.05 a | 3.67 a | |
| Cherry | Overall pleasantness | *** | 6.17 a | 5.58 a | 0.80 c | 3.22 b |
| Frankness of smell | *** | 7.67 a | 7.58 a | 0.62 c | 3.57 b | |
| Fineness of smell | ns | 1.65 | 1.75 | 0.30 | 2.97 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bedini, S.; Cosci, F.; Tani, C.; Pierattini, E.C.; Venturi, F.; Lucchi, A.; Ioriatti, C.; Ascrizzi, R.; Flamini, G.; Ferroni, G.; et al. Essential Oils as Post-Harvest Crop Protectants against the Fruit Fly Drosophila suzukii: Bioactivity and Organoleptic Profile. Insects 2020, 11, 508. https://doi.org/10.3390/insects11080508
Bedini S, Cosci F, Tani C, Pierattini EC, Venturi F, Lucchi A, Ioriatti C, Ascrizzi R, Flamini G, Ferroni G, et al. Essential Oils as Post-Harvest Crop Protectants against the Fruit Fly Drosophila suzukii: Bioactivity and Organoleptic Profile. Insects. 2020; 11(8):508. https://doi.org/10.3390/insects11080508
Chicago/Turabian StyleBedini, Stefano, Francesca Cosci, Camilla Tani, Erika Carla Pierattini, Francesca Venturi, Andrea Lucchi, Claudio Ioriatti, Roberta Ascrizzi, Guido Flamini, Giuseppe Ferroni, and et al. 2020. "Essential Oils as Post-Harvest Crop Protectants against the Fruit Fly Drosophila suzukii: Bioactivity and Organoleptic Profile" Insects 11, no. 8: 508. https://doi.org/10.3390/insects11080508
APA StyleBedini, S., Cosci, F., Tani, C., Pierattini, E. C., Venturi, F., Lucchi, A., Ioriatti, C., Ascrizzi, R., Flamini, G., Ferroni, G., Taglieri, I., & Conti, B. (2020). Essential Oils as Post-Harvest Crop Protectants against the Fruit Fly Drosophila suzukii: Bioactivity and Organoleptic Profile. Insects, 11(8), 508. https://doi.org/10.3390/insects11080508