A Review of the Biology and Control of Whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae), with Special Reference to Biological Control Using Entomopathogenic Fungi


 , , ,
, , , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Taxonomy, Origin, and Distribution of Whitefly, B. tabaci
3. Biology of Whitefly, B. tabaci
4. Damage and Losses Caused by B. tabaci
4.1. Feeding Damage
4.2. Bemisia tabaci as a Virus Vector
5. Control and Management of Whitefly
6. Biological Control of B. tabaci with Entomopathogenic Fungi
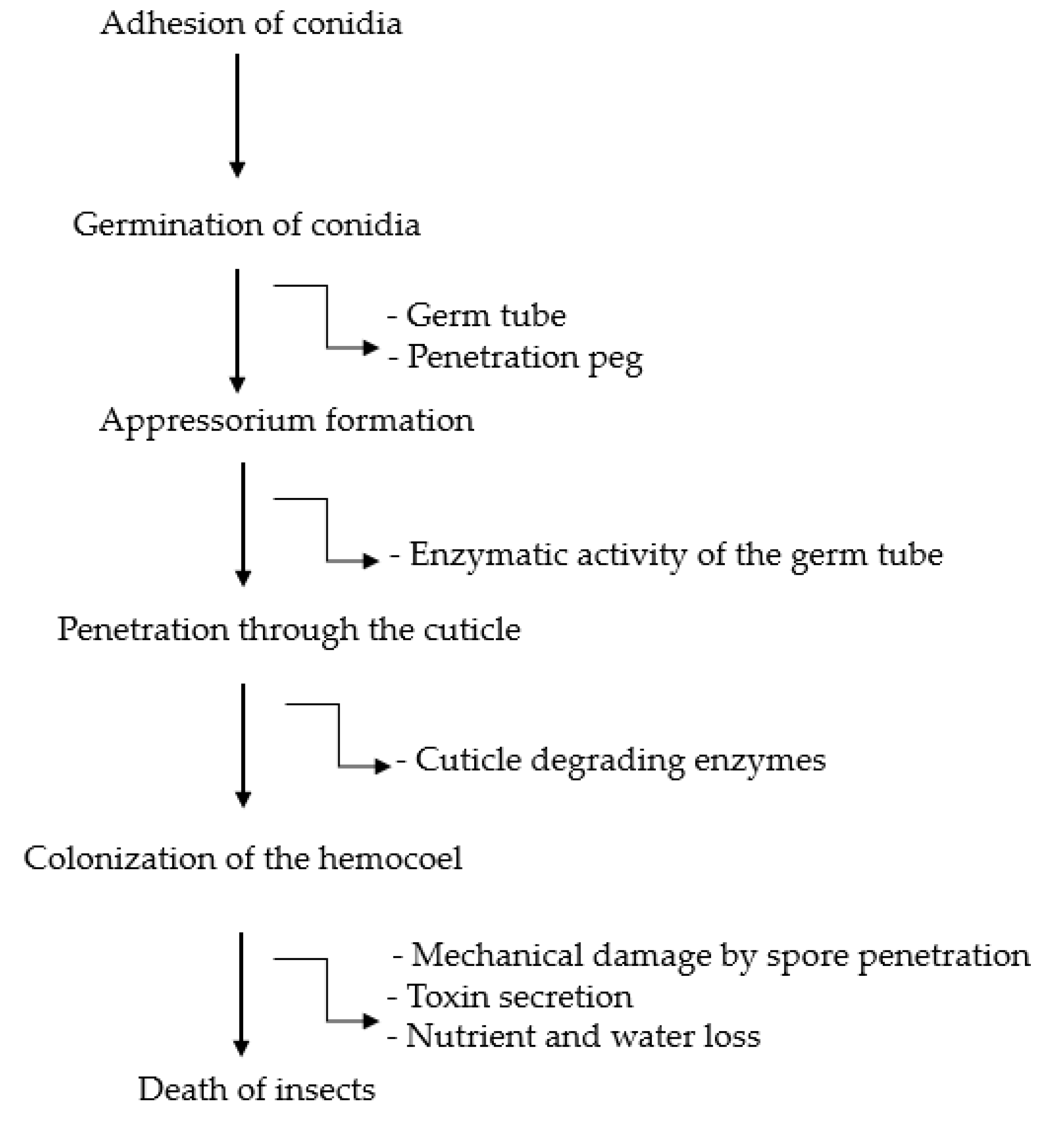
7. The Infection Process and Life Cycle of Entomopathogenic Fungi on B. tabaci
8. Merits and Demerits of Using EPF as Biological Control Measures
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Perring, T.M.; Stansly, P.A.; Liu, T.X.; Smith, H.A.; Andreason, S.A. Whiteflies: Biology, ecology, and management. In Sustainable Management of Arthropod Pests of Tomato; Wakil, W., Brust, G.E., Perring, T.M., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2018; pp. 73–110. [Google Scholar]
- Alessandro, C.P.D.; Cuthbertson, A.G.S.; Lechner, B.E. Control of Bemisia tabaci by entomopathogenic fungi isolated from arid soils in Argentina. Biocontrol Sci. Technol. 2016, 26, 1668–1682. [Google Scholar]
- Kunjwal, N.; Srivastava, R.M. Insect pests of vegetables. In Pests and Their Management; Springer Nature Singapore Pte Ltd.: Uttarakhand, India, 2018; pp. 163–221. [Google Scholar]
- Khan, I.A.; Wan, F.H. Life history of Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) biotype B on tomato and cotton host plants. J. Entomol. Zool. Stud. 2015, 3, 117–121. [Google Scholar]
- Mascarin, G.M.; Kobori, N.N.; Quintela, E.D.; Delalibera, I. The virulence of entomopathogenic fungi against Bemisia tabaci biotype B (Hemiptera: Aleyrodidae) and their conidial production using solid substrate fermentation. Biol. Control 2013, 66, 209–218. [Google Scholar] [CrossRef]
- Eslamizadeh, R.; Sajap, A.S.B.; Omar, D.B.; Azura, N.; Adam, B. Evaluation of different isolates of the entomopathogenic fungus, Paecilomyces fumosoroseus (Deuteromycotina: Hyphomycetes) against Bemisia tabaci (Hemiptera: Aleyrodidae). Biol. Control Plant Prot. 2015, 2, 82–90. [Google Scholar]
- Hasan, I.; Rasul, S.; Malik, T.H.; Qureshi, M.K.; Aslam, K.; Shabir, G.; Athar, H.; Manzoor, H. Present status of cotton leaf curl virus disease (CLCUVD): A major threat to cotton production. Int. J. Cott. Res. Technol. 2019, 1, 1–13. [Google Scholar]
- Choudhary, H.; Parihar, S.; Singh, S.; Parvez, N. Technical Bulletin on Whiteflies; Choudhary, H., Parihar, S., Singh, S., Parvez, N., Eds.; National Innovation Foundation-India: Gujarat, India, 2017. [Google Scholar]
- Polston, J.E.; Capobianco, H. Transmitting plant viruses using whiteflies. J. Vis. Exp. 2013, 81, 1–10. [Google Scholar] [CrossRef]
- Kumarasinghe, N.; Salim, N.; Wijayarathne, W. Identification and biology of two whitefly species on cassava in Sri Lanka. J. Plant Prot. Res. 2009, 49, 373–377. [Google Scholar] [CrossRef]
- Stansly, P.A.; Natwick, E.T. Integrated systems for managing Bemisia tabaci in protected and open field agriculture. In Bemisia: Bionomics and Management of a Global Pest; Springer Science+Business Media, B.V.: Dordrecht, The Netherlands, 2009; pp. 467–497. [Google Scholar]
- Gao, T.; Wang, Z.; Huang, Y.; Keyhani, N.O.; Huang, Z. Lack of resistance development in Bemisia tabaci to Isaria fumosorosea after multiple generations of selection. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S. Update on the status of Bemisia tabaci in the UK and the use of entomopathogenic fungi within eradication programmes. Insects 2013, 4, 198–205. [Google Scholar] [CrossRef]
- Smith, H.A.; Stansly, P.A.; Seal, D.R.; Mcavoy, E.; Polston, J.E.; Phyllis, R.; Schuster, D.J. Management of Whiteflies, Whitefly-Vectored Plant Virus, and Insecticide Resistance for Tomato Production; ENY-735; University of Florida, IFAS, Florida A&M University and Cooperative Extension Program: Gainesville, FL, USA, 2018; pp. 1–8. [Google Scholar]
- Skinner, M.; Parker, B.L.; Kim, J.S. Role of entomopathogenic fungi in integrated pest management. In Integrated Pest Management: Current Concepts and Ecological Perspective; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 109–191. [Google Scholar]
- Smith, H.A.; Krey, K.L. Three release rates of Dicyphus hesperus (Hemiptera: Miridae) for management of Bemisia tabaci (Hemiptera: Aleyrodidae) on greenhouse tomato. Insects 2019, 10, 213. [Google Scholar] [CrossRef]
- Jiang, W.; Peng, Y.; Ye, J.; Wen, Y.; Liu, G.; Xie, J. Effects of the entomopathogenic fungus Metarhizium anisopliae on the mortality and immune response of Locusta migratoria. Insects 2020, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Bugti, G.A.; Bin, W.; Na, C.; Feng, L.H. Pathogenicity of Beauveria bassiana strain 202 against sap-sucking insect pests. Plant Prot. Sci. 2018, 54, 111–117. [Google Scholar]
- Rios-Velasco, C.; Pérez-Corral, D.A.; Salas-Marina, M.Á.; Berlanga-Reyes, D.I.; Ornelas-Paz, J.J.; Muñiz, C.H.A.; Cambero-Campos, J.; Jacobo-Cuellar, J.L. Pathogenicity of the hypocreales fungi Beauveria bassiana and Metarhizium anisopliae against insect pests of tomato. Southwest. Entomol. 2014, 39, 739–750. [Google Scholar] [CrossRef]
- Sain, S.K.; Monga, D.; Kumar, R.; Nagrale, D.T.; Kranthi, S.; Kranthi, K.R. Comparative effectiveness of bioassay methods in identifying the most virulent entomopathogenic fungal strains to control Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Egypt. J. Biol. Pest Control 2019, 29, 1–11. [Google Scholar] [CrossRef]
- Panyasiri, C.; Attathom, T.; Poehling, H.M. Pathogenicity of entomopathogenic fungi-potential candidates to control insect pests on tomato under protected cultivation in Thailand. J. Plant Dis. Prot. 2007, 114, 278–287. [Google Scholar] [CrossRef]
- Borisade, O.A. Rearing tomato whitefly and field evaluation of modified and unmodified conidia of Beauveria bassiana, Isaria farinosa, Metarhizium anisopliae and low rates of chlorpyrifos under tropical conditions. Afr. Crops Sci. J. 2015, 23, 177–195. [Google Scholar]
- Abdel-Raheem, M.A.; Lamya, A.A.-K. Virulence of three entomopathogenic fungi against whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in tomato crop. J. Entomol. 2016, 14, 155–159. [Google Scholar]
- Zhang, C.; Ali, S.; Musa, P.D.; Wang, X.M.; Qiu, B.L. Evaluation of the pathogenicity of Aschersonia aleyrodis on Bemisia tabaci in the laboratory and greenhouse. Biocontrol Sci. Technol. 2017, 27, 210–221. [Google Scholar] [CrossRef]
- del Prado, E.N.; Iannacone, J.; Gómez, H. Effect of two entomopathogenic fungi in controlling Aleurodicus cocois (Curtis, 1846) (Hemiptera: Aleyrodidae). Chil. J. Agric. Res. 2008, 68, 21–30. [Google Scholar] [CrossRef]
- Kanakala, S.; Ghanim, M. Global genetic diversity and geographical distribution of Bemisia tabaci and its bacterial endosymbionts. PLoS ONE 2019, 14, e0213946. [Google Scholar] [CrossRef]
- Gangwar, R.K.; Charu, G. Lifecycle, distribution, nature of damage and economic importance of whitefly, Bemisia tabaci (Gennadius). Acta Sci. Agric. 2018, 2, 36–39. [Google Scholar]
- Hodges, G.S.; Evans, G.A. An Identification guide to the whiteflies (Hemiptera: Aleyrodidae) of the Southeastern United States. Fla. Entomol. 2005, 88, 518–534. [Google Scholar]
- Baig, M.M.; Dubey, A.K.; Ramamurthy, V.V. Determination of sexual dimorphism in the puparia of four whitefly pest species from India (Hemiptera: Aleyrodidae). Acta Entomol. Musei Natl. Pragae 2016, 56, 447–460. [Google Scholar]
- Boykin, L.M.; Bell, C.D.; Evans, G.; Small, I.; De Barro, P.J. Is agriculture driving the diversification of the Bemisia tabaci species complex (Hemiptera: Sternorrhyncha: Aleyrodidae)?: Dating, diversification and biogeographic evidence revealed. BMC Evol. Biol. 2013, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.-X.; Stansly, P.A.; Gerling, D. Whitefly Parasitoids: Distribution, life history, bionomics, and utilization. Annu. Rev. Entomol. 2015, 60, 273–292. [Google Scholar] [CrossRef] [PubMed]
- de Moraes, L.A.; Muller, C.; de Bueno, R.C.O.F.; Santos, A.; Bello, V.H.; De Marchi, B.R.; Watanabe, L.F.M.; Marubayashi, J.M.; Santos, B.R.; Yuki, V.A.; et al. Distribution and phylogenetics of whiteflies and their endosymbiont relationships after the Mediterranean species invasion in Brazil. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Marubayashi, J.M.; Yuki, V.A.; Rocha, K.C.G.; Mituti, T.; Pelegrinotti, F.M.; Ferreira, F.Z.; Moura, M.F.; Navas-Castillo, J.; Moriones, E.; Pavan, M.A.; et al. At least two indigenous species of the Bemisia tabaci complex are present in Brazil. J. Appl. Entomol. 2013, 137, 113–121. [Google Scholar] [CrossRef]
- De Marchi, B.R.; Kinene, T.; Mbora Wainaina, J.; Krause-Sakate, R.; Boykin, L. Comparative transcriptome analysis reveals genetic diversity in the endosymbiont hamiltonella between native and exotic populations of Bemisia tabaci from Brazil. PLoS ONE 2018, 13, e0201411. [Google Scholar]
- Shadmany, M.; Boykin, L.M.; Muhamad, R.; Omar, D. Genetic diversity of Bemisia tabaci (Hemiptera: Aleyrodidae) species complex across Malaysia. J. Econ. Entomol. 2019, 112, 75–84. [Google Scholar] [CrossRef]
- Curnutte, L.B.; Simmons, A.M.; Abd-Rabou, S. Climate change and Bemisia tabaci (Hemiptera: Aleyrodidae): Impacts of temperature and carbon dioxide on life history. Ann. Entomol. Soc. Am. 2014, 107, 933–943. [Google Scholar] [CrossRef]
- Li, N.; Li, Y.; Zhang, S.; Fan, Y.; Liu, T. Effect of elevated CO2 concentration and temperature on antioxidant capabilities of multiple generations of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae). J. Insect Physiol. 2017, 103, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Fekrat, L.; Shishehbor, P. Some biological features of cotton whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) on various host plants. Pak. J. Biol. Sci. 2007, 10, 3180–3184. [Google Scholar] [PubMed]
- Solanki, R.D.; Jha, S. Population dynamics and biology of whitefly (Bemisia tabaci Gennadius) on sunflower (Helianthus annuus L.). J. Pharmacogn. Phytochem. 2018, 7, 3055–3058. [Google Scholar]
- Smith, P.E. Crop and Food Research. In Whitefly: Identification and Biology in New Zealand Greenhouse Tomato Crops; Smith, P.E., Ed.; AsureQuality Ltd.: Auckland, New Zealand, 2009; pp. 1–8. [Google Scholar]
- Kedar, S.C.; Saini, R.K.; Kumaranag, K.M. Biology of cotton whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae) on cotton. J. Entomol. Res. 2014, 38, 135–139. [Google Scholar]
- Naranjo, S.E.; Frei, G. Chapter 6 Biology, ecology, and management of sweetpotato whiteflies on cotton. In Sticky Cotton: Causes, Effects, and Prevention; Hequet, E., Ed.; USAD: Orange County, CA, USA, 2007; pp. 51–67. [Google Scholar]
- Lindquist, R.K.; Cloyd, R.A. Identification of Insects and Related Pests of Horticultural Plants; Cuthbert, C., Carver, S.C., Eds.; O.F.A. Services, Inc.: Columbus, OH, USA, 2005; pp. 1–50. [Google Scholar]
- Samih, M.A.; Zarabi, M.; Yazdani, M.; Rouhani, M. Biological traits and life table parameters A and B biotype of Bemisia tabaci (Genn.) on cotton and rapeseed. Braz. Arch. Biol. Technol. 2014, 57, 309–316. [Google Scholar] [CrossRef][Green Version]
- Ou, D.; Ren, L.M.; Liu, Y.; Ali, S.; Wang, X.M.; Ahmed, M.Z.; Qiu, B.L. Compatibility and efficacy of the parasitoid Eretmocerus hayati and the entomopathogenic fungus Cordyceps javanica for biological control of whitefly Bemisia tabaci. Insects 2019, 10, 425. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.R. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol. 2003, 109, 195–219. [Google Scholar] [CrossRef]
- Götz, M.; Winter, S. Diversity of Bemisia tabaci in Thailand and Vietnam and indications of species replacement. J. Asia. Pac. Entomol. 2016, 19, 537–543. [Google Scholar] [CrossRef]
- Lu, S.; Chen, M.; Li, J.; Shi, Y.; Gu, Q.; Yan, F. Changes in Bemisia tabaci feeding behaviors caused directly and indirectly by cucurbit chlorotic yellows virus. Virol. J. 2019, 16, 1–14. [Google Scholar] [CrossRef]
- Legg, J.P.; Shirima, R.; Tajebe, L.S.; Guastella, D.; Boniface, S.; Jeremiah, S.; Nsami, E.; Chikoti, P.; Rapisarda, C. Biology and management of Bemisia whitefly vectors of cassava virus pandemics in Africa. Pest Manag. Sci. 2014, 70, 1446–1453. [Google Scholar] [CrossRef]
- Iqbal, M.; State, K.; Academy, M.; Naeem, M.; Aziz, U.; Khan, M. An overview of cotton leaf curl virus disease, persistent challenge for cotton production an overview of cotton leaf curl virus disease, persistent challenge for cotton production. Bulg. J. Agric. Sci. 2014, 20, 405–415. [Google Scholar]
- Liu, B.; Preisser, E.L.; Chu, D.; Pan, H.; Xie, W.; Wang, S.; Wu, Q.; Zhou, X. Multiple forms of vector manipulation by a plant-infecting virus: Bemisia tabaci and tomato yellow leaf curl virus. J. Virol. 2013, 87, 4929–4937. [Google Scholar] [CrossRef] [PubMed]
- Dutta, B.; Myers, B.; Coolong, T.; Srinivasan, B.; Sparks, A.; Riley, D. Whitefly-transmitted plant viruses in South Georgia. UGA Coop. Ext. Bull. 2018, 1507, 1–7. [Google Scholar]
- Fiallo-Olivé, E.; Pan, L.L.; Liu, S.S.; Navas-Castillo, J. Transmission of begomoviruses and other whitefly-borne viruses: Dependence on the vector species. Phytopathology 2020, 110, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Ning, W.; Shi, X.; Liu, B.; Pan, H.; Wei, W.; Zeng, Y.; Sun, X.; Xie, W.; Wang, S.; Wu, Q.; et al. Transmission of tomato yellow leaf curl virus by Bemisia tabaci as affected by whitefly sex and biotype open. Sci. Rep. 2015, 5, 10744. [Google Scholar] [CrossRef] [PubMed]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef]
- Czosnek, H.; Hariton-Shalev, A.; Sobol, I.; Gorovits, R.; Ghanim, M. The incredible journey of Begomoviruses in their whitefly vector. Viruses 2017, 9, 273. [Google Scholar] [CrossRef]
- Horowitz, A.R.; Antignus, Y.; Gerling, D. Management of Bemisia tabaci whiteflies. In The Whitefly, Bemisia Tabaci (Homoptera: Aleyrodidae) Interaction with Geminivirus-Infected Host Plants; Thompson, W.M.O., Ed.; Springer: Dordrecht, The Netherlands, 2011; pp. 293–322. [Google Scholar]
- Firdaus, S.; van Heusden, A.W.; Hidayati, N.; Supena, E.D.J.; Visser, R.G.F.; Vosman, B. Resistance to Bemisia tabaci in tomato wild relatives. Euphytica 2012, 187, 31–45. [Google Scholar] [CrossRef]
- Syed, T.S.; Abro, G.H.; Khuhro, R.D.; Dhauroo, M.H. Relative resistance of cotton varieties against sucking pests. Pak. J. Biol. Sci. 2003, 6, 1232–1233. [Google Scholar]
- Ullah, F.; Baloch, A.F.; Badshah, H. Studies of varietal resistance and chemical control against whiteflies in cotton. J. Biol. Sci. 2006, 6, 261–264. [Google Scholar] [CrossRef][Green Version]
- Silva, K.F.A.S.; Michereff-Filho, M.; Fonseca, M.E.N.; Silva-Filho, J.G.; Texeira, A.C.A.; Moita, A.W.; Torres, J.B.; Fernández-Muñoz, R.; Boiteux, L.S. Resistance to Bemisia tabaci biotype B of Solanum pimpinellifolium is associated with higher densities of type IV Glandular trichomes and acylsugar accumulation. Entomol. Exp. Appl. 2014, 151, 218–230. [Google Scholar] [CrossRef]
- Bugti, G.A. Varietal preference of insect pests on tomato crop in district Naseerabad Balochistan Pakistan. J. Entomol. Zool. Stud. JEZS 2016, 4, 328–330. [Google Scholar]
- Hamid, M.N.; Noor, M.R.; Piang, L. Non pesticide methods for controlling diseases and insect. In APO Seminar on Non Pesticide Methods for Controlling Diseases and Insect Pests; Ooi, P.A.C., Ed.; Asian Productivity Organization: Tokyo, Japan, 2002; pp. 100–111. [Google Scholar]
- Takikawa, Y.; Matsuda, Y.; Kakutani, K.; Nonomura, T.; Kusakari, S.I.; Okada, K.; Kimbara, J.; Osamura, K.; Toyoda, H. Electrostatic insect sweeper for eliminating whiteflies colonizing host plants: A complimentary pest control device in an electric field screen-guarded greenhouse. Insects 2015, 6, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Parrella, M.P.; Hanse, L.S.; van Lenteren, J.C. Glasshouse Environments. In Handbook of Biological Control; Academic Press: Cambridge, MA, USA, 1999; pp. 819–839. [Google Scholar]
- Halimatunsadiah, A.B.; Norida, M.; Omar, D.; Kamarulzaman, N.H. Application of pesticide in pest management: The case of lowland vegetable growers. Int. Food Res. J. 2016, 23, 85–94. [Google Scholar]
- Palumbo, J.C.; Horowitz, A.R.; Prabhaker, N. Insecticidal control and resistance management for Bemisia Tabaci. Crop Prot. 2001, 20, 739–765. [Google Scholar] [CrossRef]
- Naveen, N.C.; Chaubey, R.; Kumar, D.; Rebijith, K.B.; Rajagopal, R.; Subrahmanyam, B.; Subramanian, S. Insecticide resistance status in the whitefly, Bemisia tabaci genetic groups Asia-I, Asia-II-1 and Asia-II-7 on the Indian subcontinent. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Avery, P.B.; Kumar, V.; Skvarch, E.A.; Mannion, C.M.; Powell, C.A.; Mckenzie, C.L.; Osborne, L.S. An ecological assessment of Isaria fumosorosea applications compared to a neonicotinoid treatment for regulating invasive ficus whitefly. J. Fungi 2019, 5, 36. [Google Scholar] [CrossRef]
- Shinde, B.D.; Mokal, A.J.; Narangalkar, A.; Naik, K.V. Chemical management of whiteflies infesting chili. Int. J. Chem. Stud. 2018, 6, 2813–2816. [Google Scholar]
- Xu, D.; Ali, S.; Huang, Z.; Zhou, F.C.; Afzal, M.; Bashir, M.H. Influence of the entomopathogenic fungus, Verticillium lecanii on the whitefly predator, Axinoscymnus cardilobus (Coleoptera: Coccinellidae) Under Laboratory Conditions. Pak. J. Zool. 2009, 41, 289–295. [Google Scholar]
- Colmenarez, Y.; Jahnke, S.M.; Sampaio, M.V. Use of parasitoids as a biocontrol agent in the Neotropical region: Challenges and potential. In Hymenopteran Wasps-The Parasitoids; Intechopen: London, UK, 2018; pp. 1–25. [Google Scholar]
- Rodríguez, E.; Téllez, M.; Janssen, D. Whitefly Control strategies against tomato leaf curl New Delhi virus in greenhouse zucchini. Int. J. Environ. Res. Public Health 2019, 16, 2673. [Google Scholar] [CrossRef]
- Bale, J.S.; Van Lenteren, J.C.; Bigler, F. Biological control and sustainable food production. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 761–776. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Raina, T.K.; Singh, J. Entomopathogenic fungi: An effective biocontrol agent for management of insect populations naturally. J. Pharm. Sci. Res. 2017, 9, 830–839. [Google Scholar]
- Shahid, A.A.L.I.; Rao, A.Q.; Bakhsh, A.; Husnain, T. Entomopathogenic fungi as biological controllers: New insights into their virulence and pathogenicity. Arch. Biol. Sci. Belgrade 2012, 64, 21–42. [Google Scholar] [CrossRef]
- Lahey, A.; Lahey, Z.; Stansly, P. An updated list of parasitoid Hymenoptera reared from the Bemisia tabaci species complex (Hemiptera: Aleyrodidae). Fla. Entomol. 2020, 98, 456–463. [Google Scholar] [CrossRef]
- Shah, M.M.R.; Zhang, S.; Liu, T. Whitefly, Host plant and parasitoid: A Review on their whitefly, host plant and parasitoid: A review on their interactions. Asian J. Appl. Sci. Eng. 2015, 4, 48–61. [Google Scholar]
- Xu, H.; Yang, N.; Wan, F. Competitive interactions between parasitoids provide new insight into host suppression. PLoS ONE 2013, 8, e82003. [Google Scholar] [CrossRef]
- Kidane, D.; Yang, N.; Wan, F. Evaluation of a banker plant system for biological control of Bemisia tabaci (Hemiptera: Aleyrodidae) on tomato, using two aphelinid parasitoids under field-cage conditions. Biocontrol Sci. Technol. 2018, 28, 1054–1073. [Google Scholar] [CrossRef]
- Kedar, S.C.; Saini, R.K.; Kumaranag, K.M.; Sharma, S.S. Record of natural enemies of whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in some cultivated crops in Haryana. J. Biopestic. 2014, 7, 57–59. [Google Scholar]
- Nomikou, M.; Janssen, A.; Sabelis, M.W. Phytoseiid predators of whiteflies feed and reproduce on non-prey food sources. Exp. Appl. Acarol. 2003, 31, 15–26. [Google Scholar] [CrossRef]
- Khan, S.; Guo, L.; Maimaiti, Y.; Mijit, M.; Qiu, D. Entomopathogenic fungi as microbial biocontrol agent. Mol. Plant Breed. 2012, 3, 63–79. [Google Scholar] [CrossRef]
- Abdelghany, T.M. Entomopathogenic Fungi and Their Role in Biological Control; El-Ghany, T.M.A., Ed.; OMICS Group eBooks: Foster City, CA, USA, 2015; pp. 1–42. [Google Scholar]
- Ibrahim, A.A.; Mohamed, H.F.; El-Naggar, S.E.M.; Swelim, M.A.; Elkhawaga, O.E. Isolation and selection of entomopathogenic fungi as biocontrol agent against the greater wax moth, Galleria mellonella l. (Lepidoptera: Pyralidae). Egypt. J. Biol. Pest Control 2016, 26, 249–253. [Google Scholar]
- Uztan, A.H.; Gunyar, O.A.; Yoltas, A.; Keskin, N. Isolation and identification of entomopathogenic fungi Beauveria bassiana from Turkey. Fresenius Environ. Bull. 2016, 25, 5180–5185. [Google Scholar]
- Ravensberg, W.J. A Roadmap to the Successful Development and Commercialization of Microbial Pest Control Products for Control of Arthropods; Progress in Biological Control Series; Springer Science+Business Media, B.V.: Dordrecht, The Netherlands, 2011; pp. 59–127. [Google Scholar]
- Mora, M.A.E.; Rouws, J.R.C.; Fraga, M.E. Occurrence of entomopathogenic fungi in Atlantic forest soils. Microbiol. Discov. 2016, 4, 1. [Google Scholar] [CrossRef][Green Version]
- Tahira, H.G.; Saeed, S.; Khan, F.Z.A. Entomopathogenic fungi as effective insect pest management tactic: A Review. Appl. Sci. Bus. Econ. 2014, 1, 10–18. [Google Scholar]
- Anwar, W.; Ali, S.; Nawaz, K.; Iftikhar, S.; Javed, M.A.; Hashem, A.; Alqarawi, A.A.; Abd Allah, E.F.; Akhter, A. Entomopathogenic fungus Clonostachys rosea as a biocontrol agent against whitefly (Bemisia tabaci). Biocontrol Sci. Technol. 2018, 28, 750–760. [Google Scholar] [CrossRef]
- Wang, P.; Song, X.; Zhang, H. Isolation and characterization of Aschersonia placenta from citrus orchards and its pathogenicity towards Dialeurodes citri (Ashmead). J. Invertebr. Pathol. 2013, 112, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Chaverri, P.; Hodge, K.T. A taxonomic revision of the insect biocontrol fungus Aschersonia aleyrodis, its allies with white stromata and their Hypocrella sexual states. Mycol. Res. 2006, 110, 537–554. [Google Scholar] [CrossRef]
- Zhang, C.; Shao, Z.F.; Han, Y.Y.; Wang, X.M.; Wang, Z.Q.; Musa, P.D.; Qiu, B.L.; Ali, S. Effects of Aschersonia aleyrodis on the life table and demographic parameters of Bemisia Tabaci. J. Integr. Agric. 2018, 17, 389–396. [Google Scholar] [CrossRef]
- Prayogo, Y.; Bayu, M.S.Y.I. Biological control of Bemisia tabaci gennadius by using entomopathogenic fungi Aschersonia aleyrodis. In IOP Conference Series: Earth and Environmental Science; IOP Publishing Ltd.: Bristol, UK, 2020; Volume 456, pp. 1–8. [Google Scholar]
- Al-Deghari, M.A. Bioassay Evaluation of the entomopathogenic fungi, Beauveria bassiana Vuellemin against eggs and nymphs of Bemisia tabaci Gennadius (Homoptera: Aleyrodidae). Pak. J. Biol. Sci. 2008, 11, 1551–1560. [Google Scholar] [CrossRef][Green Version]
- Zafar, J.; Freed, S.; Khan, B.A.; Farooq, M. Effectiveness of Beauveria bassiana against cotton whitefly, Bemisia tabaci (Gennadius) (Aleyrodidae: Homoptera) on different host plants. Pak. J. Zool. 2016, 48, 91–99. [Google Scholar]
- Prithiva, J.N.; Ganapathy, N.; Jeyarani, S. Efficacy of different formulations of Beauveria bassiana (Bb 112) against Bemisia tabaci on tomato. J. Entomol. Zool. Stud. 2017, 5, 1239–1243. [Google Scholar]
- Arcía, G.T.O.; Eña, A.H.U.E.L.A.P.; Daniel, L.; Artínez, O.R.; Antonio, J.; Agallón, S.A. Interaction of Beauveria bassiana strain HPI-019/14 and Bacillus thuringiensis strain GP139 for the biological control of Bemisia tabaci in strawberry. Bull. Insectol. 2018, 71, 201–209. [Google Scholar]
- Nascimento Silva, J.; Mascarin, G.M.; Paula Vieira de Castro, R.; Castilho, L.R.; Freire, D.M. Novel combination of a biosurfactant with entomopathogenic fungi enhance efficacy against Bemisia whitefly. Pest Manag. Sci. 2019, 75, 2882–2891. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Omar, D.; Latif, M.A.; Morshed, M.M. The integrated use of entomopathogenic fungus, Beauveria bassiana with botanical insecticide, neem against Bemisia tabaci on eggplant. Afr. J. Microbiol. Res. 2011, 5, 3409–3413. [Google Scholar]
- Jaber, L.R.; Araj, S.E.; Qasem, J.R. Compatibility of endophytic fungal entomopathogens with plant extracts for the management of sweetpotato whitefly Bemesia tabaci Gennadius (Homoptera: Aleyrodidae). Biol. Control 2018, 117, 164–171. [Google Scholar] [CrossRef]
- Islam, M.T.; Castle, S.J.; Ren, S. Compatibility of the insect pathogenic fungus Beauveria bassiana with neem against sweetpotato whitefly, Bemisia tabaci, on eggplant. Entomol. Exp. Appl. 2010, 134, 28–34. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Collins, D.A. Tri-Tek (Petroleum Horticultural Oil) and Beauveria bassiana: Use in eradication strategies for Bemisia tabaci Mediterranean species in UK glasshouses. Insects 2015, 6, 133–140. [Google Scholar] [CrossRef]
- Huang, Z.; Ali, S.; Ren, S.; Wu, J. Effect of Isaria fumosoroseus on mortality and fecundity of Bemisia tabaci and Plutella xylostella. Insect Sci. 2010, 17, 140–148. [Google Scholar] [CrossRef]
- Kereselidze, M.; Draganova, S.; Pilarska, D.; Linde, A. Susceptibility of Lymantria monacha and L. dispar to the entomopathogenic fungus Isaria fumosorosea wize. Sumar. List 2014, 138, 185–191. [Google Scholar]
- Mascarin, G.M.; Alves, S.B.; Lopes, R.B. Culture media selection for mass production of Isaria fumosorosea and Isaria farinosa. Braz. Arch. Biol. Technol. 2010, 53, 753–761. [Google Scholar] [CrossRef]
- Loong, C.; Ahmad, S.S.; Hafidzi, M.N.; Dzolkifli, O.; Faizah, A. Effect of UV-B and solar radiation on the efficacy of Isaria fumosorosea and Metarhizium anisopliae (Deuteromycetes: Hyphomycetes) for controlling bagworm, Pterona pendula (Lepidoptera: Psychidae). J. Entomol. 2013, 10, 53–65. [Google Scholar]
- Rahim, E.; Ahmad, S.S.; Dzolkhifli, O.; Nur, A.A. First record of Isaria fumosorosea Wize (Deuteromycotina: Hyphomycetes) infecting Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in Malaysia. J. Entomol. 2013, 10, 182–190. [Google Scholar]
- Zou, C.; Li, L.; Dong, T.; Zhang, B.; Hu, Q. Joint action of the entomopathogenic fungus Isaria fumosorosea and four chemical insecticides against the whitefly Bemisia tabaci. Biocontrol Sci. Technol. 2014, 24, 315–324. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Walters, K.F.A. Pathogenicity of the entomopathogenic fungus, Lecanicillium muscarium, against the sweetpotato whitefly Bemisia tabaci under laboratory and glasshouse conditions. Mycopathologia 2005, 160, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, D.J.L.; da Silva, I.H.S.; Duarte, R.T.; Goncalves, K.C.; Polanczyk, R.A. Potential of entomopathogenic fungi as biological control agents of whitefly (Bemisia tabaci biotype B) (Genn.) (Hemiptera: Aleyrodidae). J. Exp. Agric. Int. 2019, 38, 1–8. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Walters, K.F.A.; Deppe, C. Compatibility of the entomopathogenic fungus Lecanicillium muscarium and insecticides for eradication of sweetpotato whitefly, Bemisia Tabaci. Mycopathologia 2005, 160, 35–41. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Northing, P.; Weiqi, L.; Cannon, R.J.C.; Walters, K.F.A. Further compatibility tests of the entomopathogenic fungus Lecanicillium muscarium with conventional insecticide products for control of sweetpotato whitefly, Bemisia tabaci on poinsettia plants. Insect Sci. 2008, 15, 355–360. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Northing, P.; Luo, W.; Cannon, R.J.C.; Walters, K.F.A. Chemical compatibility of the entomopathogenic fungus Lecanicillium muscarium to control Bemisia tabaci in glasshouse environment. Int. J. Environ. Sci. Technol. 2010, 7, 405–409. [Google Scholar] [CrossRef]
- Malsam, O.; Kilian, M.; Dehne, H.W. Metarhizium anisopliae biological control of the greenhouse whitefly (Trialeurodes vaporariorum). IOBC–WPRS Bull. 1998, 21, 125–128. [Google Scholar]
- Meekes, E.T.M. Entomopathogenic Fungi against Whiteflies: Trophic Interactions between Aschersonia Species, Trialeurodes Vaporariorum and Bemisia Argentifolii, and Glasshouse Crops; Wageningen University: Wageningen, The Netherlands, 2001. [Google Scholar]
- Flores, M.; Pucheta, D.; Rodriguez, N.; Torre, M.; Ramos, L. Mycoinsecticide effects of Beauveria bassiana, Metarhizium anisopliae, and Isaria fumosorosea on the whitefly Bemisia tabaci (Homoptera: Aleyrodidae) in different strata of bean. Afr. J. Microbiol. Res. 2012, 6, 7246–7252. [Google Scholar]
- Norhelina, L.; Sajap, A.S.; Mansour, S.A.; Idris, A.B. Infectivity of five Metarhizium anisopliae (Deuteromycota: Hyphomycetales) strains on whitefly, Bemisia tabaci (homoptera: Aleyrodidae) infesting brinjal, Solanum Melongena. Acad. J. Entomol. 2013, 6, 127–132. [Google Scholar]
- Islam, M.T.; Omar, D.; Shabanimofrad, M. Molecular identification and virulence of six isolates of Metarhizium anisopliae (Deuteromycotina: Hyphomycetes) to Bemisia tabaci Q biotype. J. Asia. Pac. Entomol. 2014, 17, 237–241. [Google Scholar] [CrossRef]
- Islam, M.T.; Omar, D.; Shah, A.K.M.A.; Hasan, M.F. Virulence of entomopathogenic fungus, Metarhizium anisopliae to Sweetpotato whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae) under osmotic stress. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 2016, 86, 617–622. [Google Scholar] [CrossRef]
- Qiu, J.; Song, F.; Mao, L.; Tu, J.; Guan, X. Time-dose-mortality data and modeling for the entomopathogenic fungus. Can. J. Microbiol. 2013, 101, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Maranha, E.A.; Maranha, E. Host plant influences pathogenicity of Beauveria bassiana to Bemisia tabaci and its sporulation on cadavers. BioControl 2006, 51, 519–532. [Google Scholar]
- Imam, I.I. Role of certain Beauveria bassiana isolate as biological control agent against whitefly, Bemisia tabaci (Genn.) and its effect on the predator Chrysopela carnea (stephens). Egypt. J. Desert Res. 2017, 67, 351–359. [Google Scholar] [CrossRef][Green Version]
- Cabanillas, H.E.; Jones, W.A. Pathogenicity of Isaria sp. (Hypocreales: Clavicipitaceae) against the sweet potato whitefly B biotype, Bemisia tabaci (Hemiptera: Aleyrodidae). Crop Prot. 2009, 28, 333–337. [Google Scholar] [CrossRef]
- Tian, J.; Diao, H.; Liang, L.; Arthurs, S.; Ma, R. Pathogenicity of Isaria fumosorosea to Bemisia tabaci, with some observations on the fungal infection process and host immune response. J. Invertebr. Pathol. 2015, 130, 147–153. [Google Scholar] [CrossRef]
- Wang, L.; Huang, J.; You, M.; Guan, X.; Liu, B. Toxicity and feeding deterrence of crude toxin extracts of Lecanicillium (Verticillium) Lecanii (Hyphomycetes) against sweetpotato whitefly, Bemisia tabaci (Homoptera: Aleyrodidae). Pest Manag. Sci. 2007, 63, 381–387. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Walters, K.F.A.; Northing, P. The susceptibility of immature stages of Bemisia tabaci to the entomopathogenic fungus Lecanicillium muscarium on tomato and verbena foliage. Mycopathologia 2005, 159, 23–29. [Google Scholar] [CrossRef]
- Keerio, A.U.; Nazir, T.; Abdulle, Y.A.; Jatoi, G.H.; Gadhi, M.A.; Anwar, T.; Sokea, T.; Qiu, D. In vitro pathogenicity of the fungi Beauveria bassiana and Lecanicillium lecanii at different temperatures against the whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt. J. Biol. Pest Control 2020, 300, 1–9. [Google Scholar] [CrossRef]
- Reyad, N.F. Empact of entomopathogenic fungi on whitefly, Bemisia tabaci in tomato crop in Egypt. Int. J. ChemTech Res. 2017, 10, 372–377. [Google Scholar]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, S.S.; Sharma, A.K.; Beniwal, V.; Goel, G.; Batra, P.; Kumar, A.; Jaglan, S.; Sharma, A.K.; Malhotra, S. Myco-Biocontrol of insect pests: Factors involved, mechanism, and regulation. J. Pathog. 2012, 2012, 126819. [Google Scholar] [CrossRef]
- Vega, F.E.; Meyling, N.V.; Luangsa-Ard, J.J.; Blackwell, M. Fungal entomopathogens. In Insect Pathology; Vega, F.E., Harry, K.K., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; pp. 171–220. [Google Scholar]
- Sinha, K.K.; Choudhary, A.K.; Kumari, P. Entomopathogenic Fungi. In Ecofriendly Pest Management for Food Security; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 475–505. [Google Scholar]
- Augustyniuk-Kram, A.; Kram, A.J. Entomopathogenic fungi as an important natural regulator of insect outbreaks in forests (Review). In Forest Ecosystems-More Than Just Trees; IntechOpen: London, UK, 2012; pp. 266–294. [Google Scholar]
- Agro, S.; Road, M.; Nadu, T. Entomopathogenic Fungi for the Control of Economically Important Whiteflies. J. Biol. Control 2007, 21, 29–36. [Google Scholar]
- Meyling, N.V. Methods for Isolation of Entomopathogenic Fungi from the Soil Environment. Available online: https://orgprints.org/11200/ (accessed on 8 September 2020).
- Mantzoukas, S.; Eliopoulos, P.A. Endophytic entomopathogenic fungi: A valuable biological control tool against plant pests. Appl. Sci. 2020, 10, 360. [Google Scholar] [CrossRef]
- Butt, T.M.; Jackson, C.; Magan, N. Fungal Biological Control Agents: Progress, problems and potential. In Fungi as Biocontrol Agents: Progress, Problems and Potential; Butt, T.M., Jackson, C., Magan, N., Eds.; CAB International: Wallingford, UK, 2001; pp. 1–8. [Google Scholar]
- Inglis, G.D.; Goettel, M.S.; Butt, T.M.; Strasser, H. Use of hyphomycetous fungi for managing insect pests. In Fungi as Biocontrol Agents; Butt, T.M., Jackson, C., Magan, N., Eds.; CAB International: Wallingford, UK, 2001; pp. 26–69. [Google Scholar]


{kind=link}
| Species | Bioassay Method | Significant Effects/Results | Country Where Work Was Conducted | Ref. |
|---|---|---|---|---|
| Aschersonia aleyrodis | Second and third instars on eggplant leaves were sprayed with fungal spores at 1 × 107 conidia/mL. | Mortality: Second instar: 71.21% Third instar: 69.31% Pupae: 53.36% LC50: Second instar: 7.93 × 106; third instar: 1.08 × 107; pupae: 1.56 × 107 conidia/mL LT50: 4.60 days for second instar. | China | [24] |
| A. aleyrodis | First, second, and third instars on eggplant were sprayed with spores at 1 × 107 conidia/mL on eggplant leaves. | The survival of first, second, and third nymphal instars was significantly affected. | China | [93] |
| A. aleyrodis | Eggs; first, second, third, fourth instars; and adults on the leaves of soybean were sprayed with 1 × 106 conidia/mL. | The highest mortality (99%) was observed for the first, second, and third instars and the lowest mortality in the adult stage. LD50: 6–7 × 106 conidia/mL LT50: 3.50–3.75 days for nymph stage; 4.50 days for adults. | Indonesia | [94] |
| A. placenta | First, second, and third instars on tomato were sprayed with 1.5 mL of fungal suspension. | Mortality ranged from 93% to 100%. LD50 and LD90 values decreased with time and increased with instar. LT50 values decreased with conidial concentrations. | China | [121] |
| Beauveria bassiana | Eggs and nymphs were sprayed with different concentrations on plant leaves of cotton, tomato, eggplant, and bell pepper. | Most effective isolate (Bb-01) on cotton mortality: eggs: 65.30% nymphs: 88.82% LC50 value: 2.4 × 107 spores/mL. LT50: lowest on cotton, 5.40 days | Pakistan | [96] |
| B. bassiana | Nymphs and adults on tomato leaves were sprayed with different formulations of 108 spores/mL. | Reduction of the population over control in formulations: Oil: 45.86%. Talc: 29.62%. Crude: 21.63%. | India | [97] |
| B. bassiana | Eggs and first, second, third, and fourth instars were immersed in 1 mL of conidia suspension for 10 min. | First and second instars were more susceptible than the third and fourth instars. Nymphs were highly susceptible compared to eggs. | Saudi Arabia | [95] |
| B. bassiana | Fourth instars from cucumber, tomato, melon, green pepper, potato, eggplant, marrow, cabbage, bean, and cotton plants were immersed in 1.0 × 107 conidia/mL for 10 s. | Mortality and average survival time after 8 days of inoculation were significantly influenced by the host plants. Mean mortality ranged between 52.3 ± 7.3 for nymphs reared on cotton and 91.8 ± 5.8 for nymphs reared on cucumber. | Spain | [122] |
| B. bassiana | Adult B. tabaci (2–3 days old) on cotton were sprayed with three concentrations (1 × 103, 1 × 105, and 1 × 107 spores/mL) of 1 mL of fungal suspension. | Mortality recorded at the lowest dose (1 × 103 spores/mL) was 11%, while the highest percentage mortality (56%) was recorded at a high dose of 1 × 107 spores/mL, and the recorded natural mortality was only 5%. | Egypt | [123] |
| Isaria spp. | Second, third, and fourth instars were sprayed with spore concentrations in clip-screen cages on sweet potatoes. | LC50 and LT50 values when exposed to 1000 spores/mm2: LC50: second instar: 72–118 spores/mm2; third instar: 166–295 spores/mm2; fourth instar: 166–295 spores/mm2 LT50: second instar: 3 days; third instar: 4 days. | USA | [124] |
| I. fumosoroseus | Eggs and first, second, third, and fourth instars on eggplants were dipped in conidia suspension (1 × 106 conidia/mL) for 2–3 s. | Most effective isolate (UPM isolate) mortality: Eggs: 91% Second instar: 90% Third instar: 86% Fourth instar: 89% LT50: 3.94 to 6.28 days. | Malaysia | [108] |
| I. fumosoroseus | First, second, and third instars on cucumbers were sprayed with spores at 1 × 107 conidia/mL. | The second instar was the most susceptible life stage with mortality rate at 83% after 7 days of application. | China | [125] |
| I. fumosoroseus | Second instars on eggplants were dipped in five different concentrations (1 × 103,1 × 104,1 × 105,1 × 106,1 × 107 conidia/mL) for 20 s. | LC50 values: 1.10 × 104 conidia/mL after 12 days of treatment. At a concentration of 1 × 107 conidia/mL, minimum average longevity and number of progenies produced were 9 days and 10.92 eggs/female, respectively, as compared to 16.3 days and 83.67 eggs/female for the control. | China | [104] |
| Lecanicillium lecanii | Second instars on five host plants were sprayed with 107 conidia/mL. | The highest mortality was recorded in all host plants: Laboratory conditions: nymphal mortality: >90% Glasshouse conditions: nymphal mortality: 81%. | United Kingdom | [110] |
| L. lecanii | Toxin emulsion was applied to female adult B. tabaci on the tomato plants. | The toxin reduced the hatching of whitefly eggs, the survival rate of the nymphs, and the emergence and fecundity of the progeny adults. | China | [126] |
| L. muscarium | First, second, third, and fourth instars on tomato and verbena plants were sprayed with a fungal suspension (107 spores/mL). | First and second instar nymphs were more susceptible to L. muscarium than the third and fourth instar nymphs. | United Kingdom | [127] |
| Metarhizium anisopliae | Second instar nymphs were sprayed with 107 spores/mL of three vegetable oil formulations. | The highest mortality was observed with sunflower oil followed by olive oil and maize oil formulations. | Argentina | [2] |
| M. anisopliae | Second instar nymphs on eggplants were dipped into 108 conidia/mL for 10 s. | Mortality caused by two isolates under osmotic conditions was 83.9% and 83.8%. | Malaysia | [120] |
| M. anisopliae | Second and fourth instars on brinjals were sprayed with 2 mL of 107, 105, 103, and 10 conidia/mL. | LC50 value: Lowest on second instar, 6.62 × 10 conidia/mL. LT50: 2.25 days | Malaysia | [118] |
| M. anisopliae | Second instars on eggplants were dipped into 108 conidia/mL for 10 s. | The highest mortality of 84.3% was observed in the isolate GT3. | Malaysia | [119] |
| B. bassiana, I. fumosorosea, and L. muscarium | Nymphs were sprayed with conidia at 107 conidia/mL (150 conidia/mm2). | All fungi isolates were pathogenic to whitefly nymphs. Isolates of B. bassiana and I. fumosorosea were significantly more virulent than that of L. muscarium, with > 77% nymphal mortalities. | Brazil | [5] |
| B. bassiana, M. anisopliae, and I. fumosorosea | Conidia at 5 × 109 conidia/ha were sprayed on eggs; first, second, third, and fourth instars, and adult B. tabaci on the underside of the leaves to the point of runoff. | There were no significant differences in individual mortality for all life stages between the different strata (the top, middle, and bottom thirds). M. anisopliae was significantly more effective against eggs; first, second, and third instar nymphs; and pupae. | Mexico | [117] |
| B. bassiana and L. lecanii | Nymphs on tomato plants were sprayed with fungal filtrate, conidia, and filtrate + conidia of two fungal strains. | In all the three bioassays, the isolate BB-72 was the most virulent, causing high mortality using all three different concentrations of the two fungal strains tested. | China | [128] |
| M. anisopliae, B. bassiana, and V. lecanii | Three different concentrations (1 × 107, 1 × 108, and 1 × 109 spores/mL) were sprayed onto adult B. tabaci. | The percentage of reduction ranged between 52% and 100% in all concentrations. All the treatments caused 100% mortality with concentrations of 1 × 109 after the sixth day of application. | Egypt | [23] |
| M. anisopliae, B. bassiana, and V. lecanii | Three different concentrations (2 × 103, 2 × 104, and 2 × 105 conidia/mL) were sprayed onto adult B. tabaci. | The percentage of mortality ranged between 80–100% (V. lecanii), 45–75% (B. bassiana), and 45–70% (M. anisopliae) on the seventh day after treatment. The concentration of 2 × 105 conidia/mL was highly toxic under both laboratory and field conditions. | Egypt | [129] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sani, I.; Ismail, S.I.; Abdullah, S.; Jalinas, J.; Jamian, S.; Saad, N. A Review of the Biology and Control of Whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae), with Special Reference to Biological Control Using Entomopathogenic Fungi. Insects 2020, 11, 619. https://doi.org/10.3390/insects11090619
Sani I, Ismail SI, Abdullah S, Jalinas J, Jamian S, Saad N. A Review of the Biology and Control of Whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae), with Special Reference to Biological Control Using Entomopathogenic Fungi. Insects. 2020; 11(9):619. https://doi.org/10.3390/insects11090619
Chicago/Turabian StyleSani, Ibrahim, Siti Izera Ismail, Sumaiyah Abdullah, Johari Jalinas, Syari Jamian, and Norsazilawati Saad. 2020. "A Review of the Biology and Control of Whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae), with Special Reference to Biological Control Using Entomopathogenic Fungi" Insects 11, no. 9: 619. https://doi.org/10.3390/insects11090619
APA StyleSani, I., Ismail, S. I., Abdullah, S., Jalinas, J., Jamian, S., & Saad, N. (2020). A Review of the Biology and Control of Whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae), with Special Reference to Biological Control Using Entomopathogenic Fungi. Insects, 11(9), 619. https://doi.org/10.3390/insects11090619