Necrobia rufipes (De Geer) Infestation in Pet Food Packaging and Setup of a Monitoring Trap

 ,
, 
 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Behavioral Tests on Pet Food Packaging
2.3. Suitability Tests of Adhesives for Traps
2.4. Bioassays with Food Attractants
2.4.1. Four-Choice Pitfall Olfactometer Bioassays
2.4.2. Dual-Choice Arena Bioassays
2.5. Statistical Analysis
3. Results
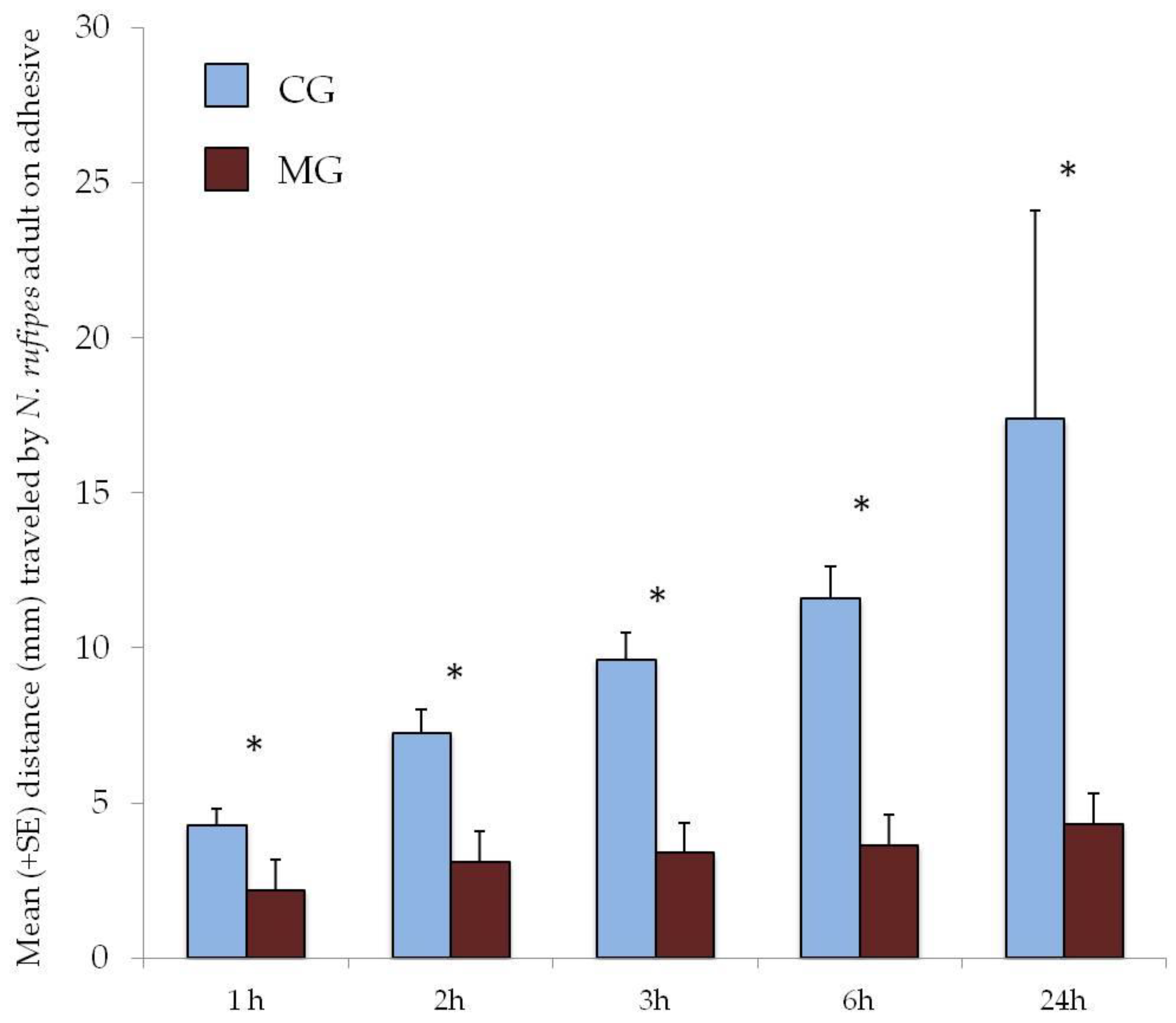
3.1. Behavioral Tests on Pet Food Packaging
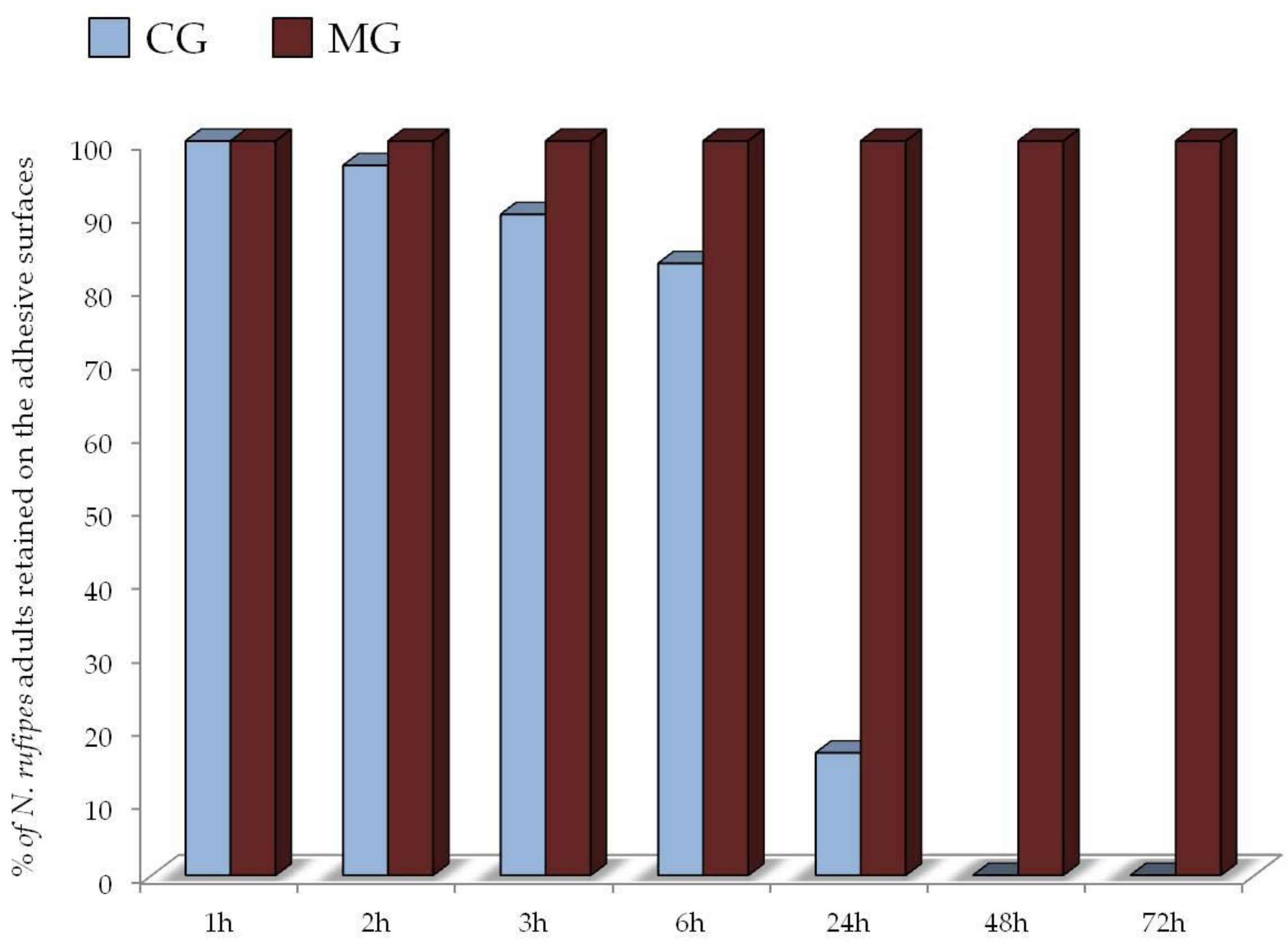
3.2. Suitability Test of Adhesives for Traps
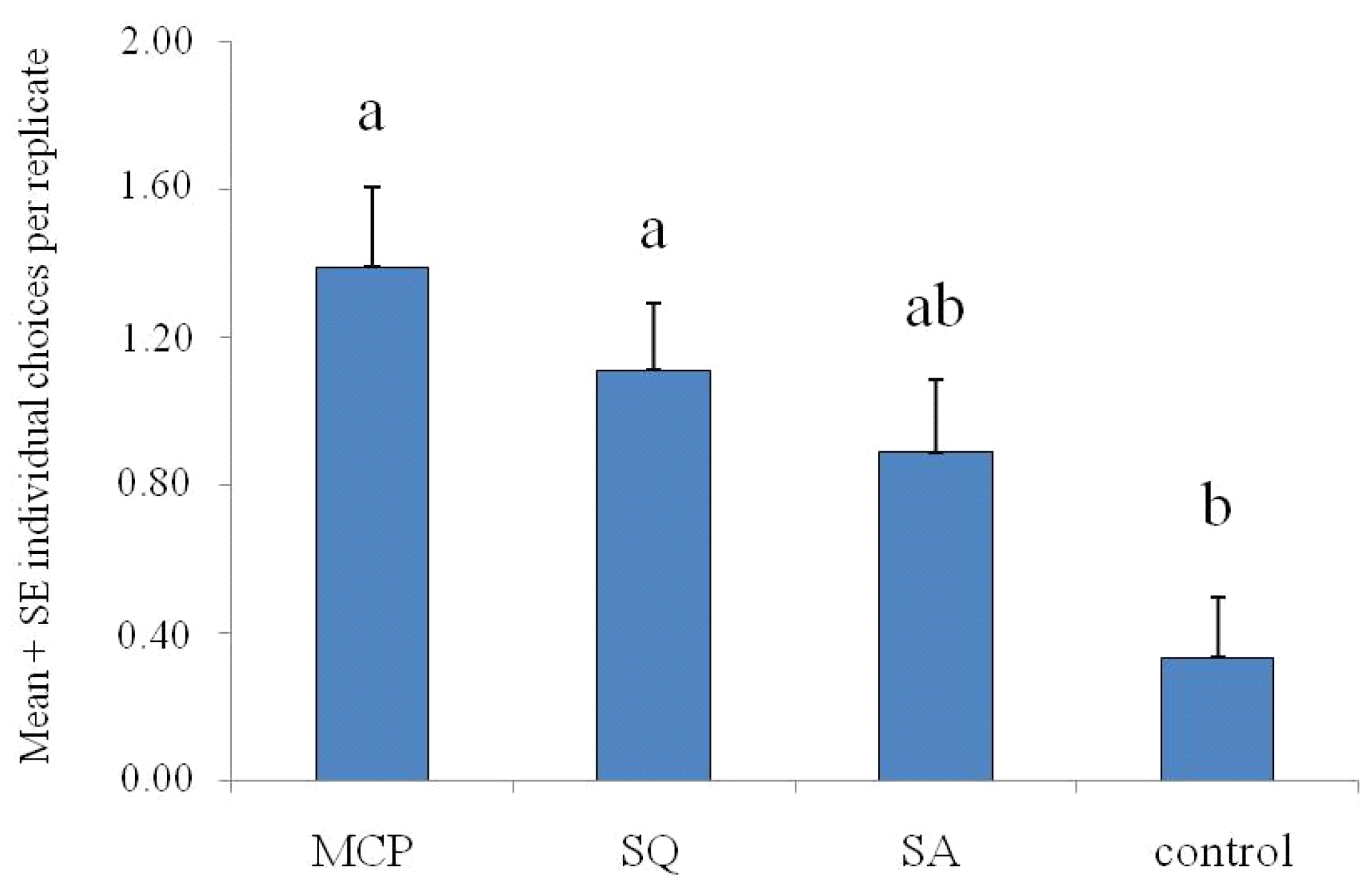
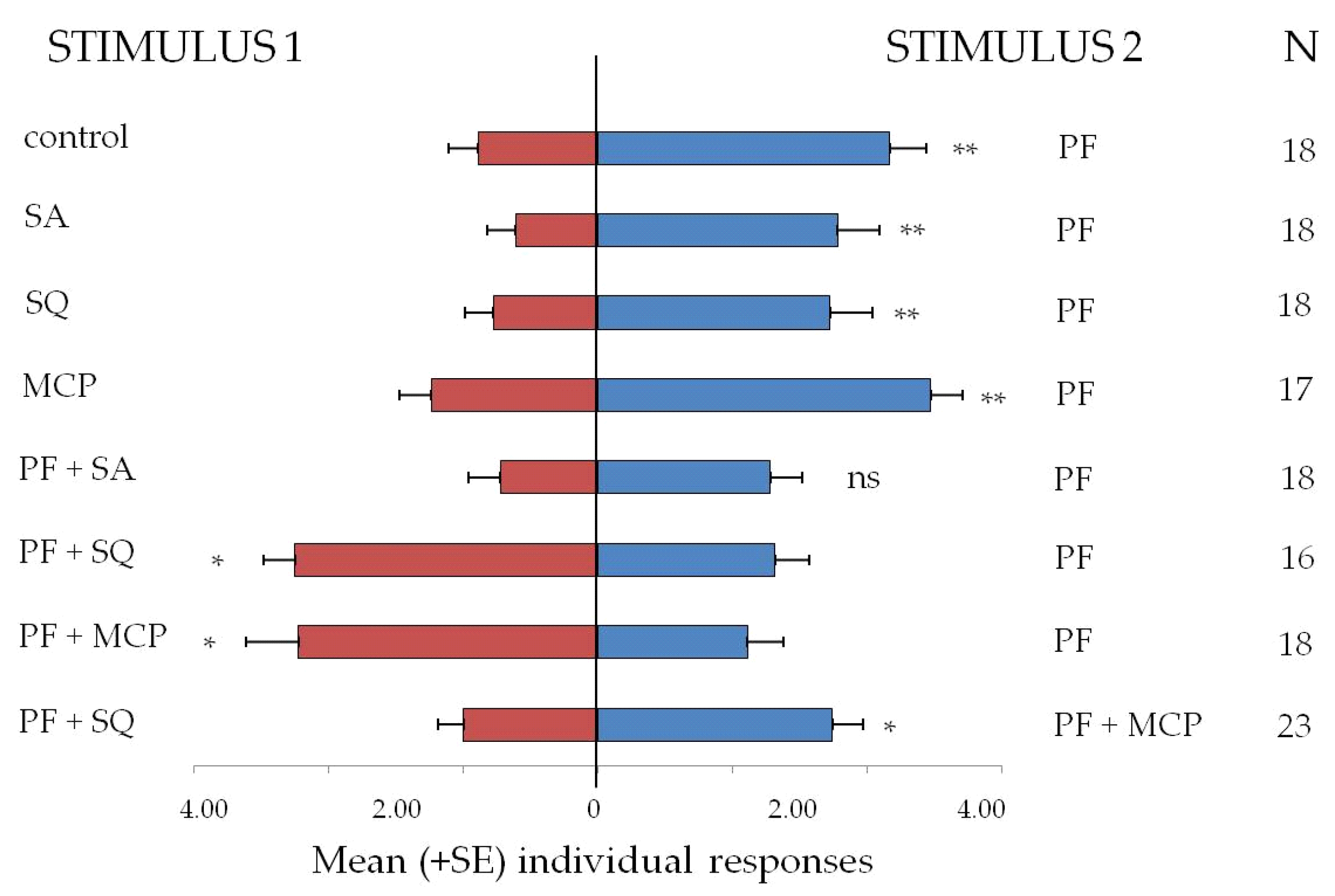
3.3. Bioassays with Food Attractants
3.3.1. Four-Choice Pitfall Olfactometer Bioassays
3.3.2. Dual-Choice Arena Bioassays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Son, S.J.; Bae, J.M.; Park, S.J.; Lee, H.J.; Lee, H.S. A new area of food industry; companion animal market. Food Sci. Ind. 2017, 50, 92–103. [Google Scholar]
- Viana, L.M.; Mothé, C.G.; Mothé, M.G. Natural food for domestic animals: A national and international technological review. Res. Vet. Sci. 2020, 130, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Market Data Forecast, Industry Reports & Business Intelligence. Available online: https://www.marketdataforecast.com (accessed on 29 June 2020).
- Roesli, R.; Subramanyam, B.; Campbell, J.F.; Kemp, K. Stored-product insects associated with a retail pet store chain in Kansas. J. Econ. Entomol. 2003, 96, 1958–1966. [Google Scholar] [CrossRef] [PubMed]
- Subramanyam, B.; Campbell, J.; Kemp, K. It’s in the detail for retail-A study is under way to determine the best management methods in the retail environment. Pest Control 2001, 69, 26–31. [Google Scholar]
- Platt, R.R.; Cuperus, G.W.; Payton, M.E.; Bonjour, E.L.; Pinkston, K.N. Integrated pest management perceptions and practices and insect populations in grocery stores in South-Central United States. J. Stored Prod. Res. 1998, 34, 1–10. [Google Scholar] [CrossRef]
- Arbogast, R.T.; Kendra, P.E.; Mankin, R.W.; McGovern, J.E. Monitoring insect pests in retail stores by trapping and spatial analysis. J. Econ. Entomol. 2000, 93, 1531–1542. [Google Scholar] [CrossRef]
- dos Santos, A.T.; Pereira, T.N.; Bicho, C.L. Necrobia rufipes (Degeer, 1775) (Coleoptera: Cleridae) Infesting Industrialized Foods for Pet Animals in Campina Grande, Paraiba State, Brazil. Entomol. News 2020, 129, 81–85. [Google Scholar] [CrossRef]
- Gredilha, R.; Lima, A.F. First record of Necrobia rufipes (De Geer, 1775) (Coleoptera; Cleridae) associated with pet food in Brazil. Braz. J. Biol. 2007, 67, 187. [Google Scholar] [CrossRef] [Green Version]
- Koç, N.; Arslanbaş, M.; Tiftikçioğlu, C.; Nalbantoğlu, S.; Çakmak, A. Evaluation of relation with pet food and first record of Necrobia rufipes (De Geer, 1775) (Coleoptera: Cleridae) associated with pet clinic in Turkey. Vet. Hekim Der. Derg. 2019, 91, 44–48. [Google Scholar] [CrossRef]
- Simmons, P.; Ellington, G.W. The ham beetle, Necrobia rufipes DE GEER. J. Agr. Res. 1925, 30, 845–863. [Google Scholar]
- Hasan, M.M.; Athanassiou, C.G.; Schilling, M.W.; Phillips, T.W. Biology and management of the red-legged ham beetle, Necrobia rufipes DeGeer (Coleoptera: Cleridae). J. Stored Prod. Res. 2020, 88, 101635. [Google Scholar] [CrossRef]
- Savoldelli, S.; Frignani, M.; Süss, L. Necrobia rufipes (De Geer): An emerging pest associated with pet store chain in Europe. In Proceedings of the 12th International Working Conference on Stored Product Protection (IWCSPP), Berlin, Germany, 7–11 October 2018; pp. 126–127. [Google Scholar]
- Ashman, F. Factors affecting the abundance of the copra beetle, Necrobia rufipes (Deg.) (Col., Cleridae). Bull. Entomol. Res. 1963, 53, 671–680. [Google Scholar] [CrossRef]
- Nalinakumari, T.; Visalakshi, A.; Mammen, K.V. Control of major pest of copra with phosphine fumigation. J. Trop. Agr. 1998, 36, 78–79. [Google Scholar]
- Hasan, M.M.; Aikins, M.J.; Schilling, M.W.; Phillips, T.W. Comparison of Methyl Bromide and Phosphine for Fumigation of Necrobia rufipes (Coleoptera: Cleridae) and Tyrophagus putrescentiae (Sarcoptiformes: Acaridae), Pests of High-Value Stored Products. J. Econ. Entomol. 2020, 113, 1008–1014. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.F.; Mullen, M.A.; Dowdy, A.K. Monitoring stored-product pests in food processing plants with pheromone trapping, contour mapping, and mark-recapture. J. Econ. Entomol. 2002, 95, 1089–1101. [Google Scholar] [CrossRef]
- Savoldelli, S.; Trematerra, P. Mass-trapping, mating-disruption and attracticide methods for managing stored-product insects: Success stories and research needs. Stewart Postharv. Rev. 2011, 7, 1–8. [Google Scholar]
- Guarino, S.; Basile, S.; Caimi, M.; Carratello, A.; Manachini, B.; Peri, E. Insect pests of the Herbarium of the Palermo botanical garden and evaluation of semiochemicals for the control of the key pest Lasioderma serricorne F. (Coleoptera: Anobiidae). J. Cul. Herit. 2019, 43, 37–44. [Google Scholar] [CrossRef]
- Hasan, M.M.; Phillips, T.W. Mass-rearing of the redlegged ham beetle, Necrobia rufipes De Geer (Coleoptera: Cleridae) for laboratory research. J. Stored Prod. Res. 2010, 46, 38–42. [Google Scholar] [CrossRef]
- Stejskal, V.; Bostlova, M.; Nesvorna, M.; Volek, V.; Dolezal, V.; Hubert, J. Comparison of the resistance of mono-and multilayer packaging films to stored-product insects in a laboratory test. Food Control 2017, 73, 566–573. [Google Scholar] [CrossRef]
- Forbes, S.L.; Stuart, B.H.; Dadour, I.R.; Dent, B.B. A preliminary investigation of the stages of adipocere formation. J. Forensic Sci. 2004, 49, JFS2002230-9. [Google Scholar] [CrossRef]
- Mpuchane, S.; Allotey, J.; Matsheka, I.; Simpanya, M.; Coetzee, S.; Jordaan, A.; Mrema, N.; Gashe, B.A. Carriage of micro-organisms by domestic cockroaches and implications on food safety. Int. J. Trop. Ins. Sci. 2006, 26, 166–175. [Google Scholar]
- Berger, R.G. Aroma compounds in food. In Aroma Biotechnology; Springer: Berlin/Heidelberg, Germany, 1995; pp. 1–10. [Google Scholar]
- Athanassiou, C.G.; Riudavets, J.; Kavallieratos, N.G. Preventing stored-product insect infestations in packaged-food products. Stew. Post. Rev. 2011, 3, 1–5. [Google Scholar]
- Mowery, S.V.; Mullen, M.A.; Campbell, J.F.; Broce, A.B. Mechanisms underlying sawtoothed grain beetle (Oryzaephilus surinamensis [L.]) (Coleoptera: Silvanidae) infestation of consumer food packaging materials. J. Econ. Entomol. 2002, 95, 1333–1336. [Google Scholar] [CrossRef] [PubMed]
- Riudavets, J.; Salas, I.; Pons, M.J. Damage characteristics produced by insect pests in packaging film. J. Stored Prod. Res. 2007, 43, 564–570. [Google Scholar] [CrossRef]
- Adler, C. Insect-proof packaging to avoid stored product insects. In Proceedings of the Working Group Integrated Protection of Stored Product, Poznan, Poland, 20–23 August 2007; Navarro, S., Adler, C., Stengård Hansen, L., Eds.; IOBC/Wprs Bulletin: Zurich, Switzerland, 2008; Volume 40, pp. 363–369. [Google Scholar]
- Trematerra, P.; Savoldelli, S. Pasta preference and ability to penetrate through packaging of Sitophilus zeamais Motschulsky (Coleoptera: Dryophthoridae). J. Stored Prod. Res. 2014, 59, 126–132. [Google Scholar] [CrossRef]
- Yee, W.L. Evaluation of yellow rectangle traps coated with hot melt pressure sensitive adhesive and sticky gel against Rhagoletis indifferens (Diptera: Tephritidae). J. Econ. Entomol. 2011, 104, 909–919. [Google Scholar] [CrossRef]
- Lo, P.L.; Wallis, R.; Bellamy, D.E. The effectiveness of two types of adhesive for catching insects in traps. N. Z. Plant Prot. 2019, 72, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Solomon, F.; Belayneh, F.; Kibru, G.; Ali, S. Vector potential of Blattella germanica (L.) (Dictyoptera: Blattidae) for medically important bacteria at food handling establishments in Jimma town, Southwest Ethiopia. Biomed Res. Int. 2016. [Google Scholar] [CrossRef] [Green Version]
- Walter, R.H.; Fagerson, I.S. Volatile Compounds from-Heated Glucose. J. Food Sci. 1968, 33, 294–297. [Google Scholar] [CrossRef]
- MacLeod, G. The flavor of beef. In Flavor of Meat and Meat Products; Springer: Boston, MA, USA, 1994; pp. 4–37. [Google Scholar]
- Sampels, S.; Pickova, J.; Wiklund, E. Influence of production system, age and sex on carcass parameters and some biochemical meat quality characteristics of reindeer (Rangifer tarandus tarandus L.). Rangifer 2005, 25, 85–96. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, J.; Pei, Z.; Wei, P.; Xiang, D.; Cao, X.; Shen, X.; Li, C. Volatile flavour components and the mechanisms underlying their production in golden pompano (Trachinotus blochii) fillets subjected to different drying methods: A comparative study using an electronic nose, an electronic tongue and SDE-GC-MS. Food Res. Int. 2019, 123, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Odeyemi, O.O.; Owoade, R.A.; Akinkurolere, R. Toxicity and population suppression effects of Parkia clappertoniana on dried fish pests, (Dermestes maculatus and Necrobia rufipes). Glob. J. Pure Appl. Sci. 2000, 6, 191–195. [Google Scholar]
- Koppel, K.; Adhikari, K.; Di Donfrancesco, B. Volatile compounds in dry dog foods and their influence on sensory aromatic profile. Molecules 2013, 18, 2646–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Chen, X.; Nsor-Atindana, J.; Masamba, K.G.; Ma, J.; Zhong, F. Optimization of key aroma compounds for dog food attractant. Anim. Feed Sci. Tech. 2017, 225, 173–181. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savoldelli, S.; Jucker, C.; Peri, E.; Arif, M.A.; Guarino, S. Necrobia rufipes (De Geer) Infestation in Pet Food Packaging and Setup of a Monitoring Trap. Insects 2020, 11, 623. https://doi.org/10.3390/insects11090623
Savoldelli S, Jucker C, Peri E, Arif MA, Guarino S. Necrobia rufipes (De Geer) Infestation in Pet Food Packaging and Setup of a Monitoring Trap. Insects. 2020; 11(9):623. https://doi.org/10.3390/insects11090623
Chicago/Turabian StyleSavoldelli, Sara, Costanza Jucker, Ezio Peri, Mokhtar Abdulsattar Arif, and Salvatore Guarino. 2020. "Necrobia rufipes (De Geer) Infestation in Pet Food Packaging and Setup of a Monitoring Trap" Insects 11, no. 9: 623. https://doi.org/10.3390/insects11090623
APA StyleSavoldelli, S., Jucker, C., Peri, E., Arif, M. A., & Guarino, S. (2020). Necrobia rufipes (De Geer) Infestation in Pet Food Packaging and Setup of a Monitoring Trap. Insects, 11(9), 623. https://doi.org/10.3390/insects11090623