
Host Plants of the Immature Stages of the Invasive Longan Lanternfly, Pyrops candelaria (L.) (Hemiptera, Fulgoridae) in Taiwan


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Locations
2.2. Insect Recording
3. Results
3.1. Egg Masses on Different Plant Species
3.2. Nymphs on Different Plant Species
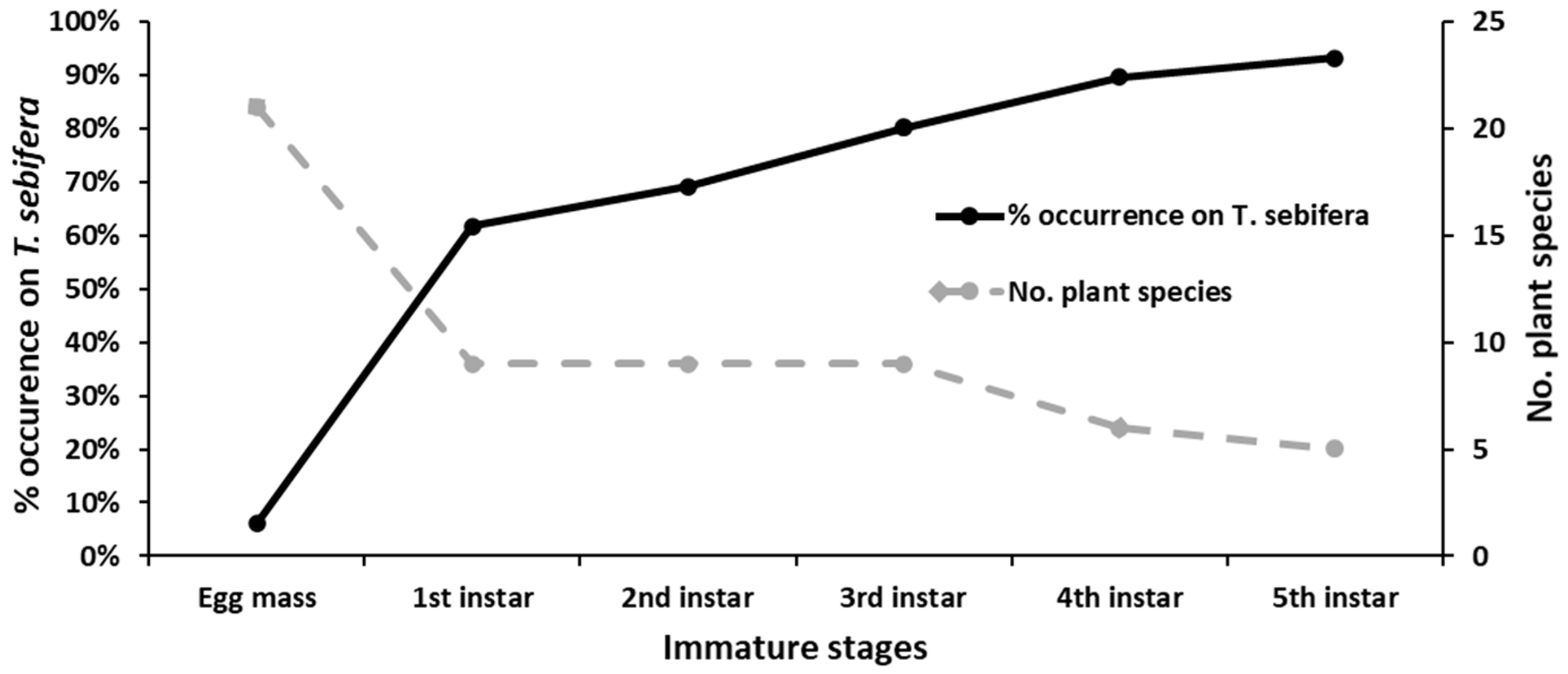
3.3. The Shift of Host Plant Preference According to the Developmental Stage
3.4. Monthly Records of the Immature Stages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wang, W.; Xu, S.; Qin, D. The lanternfly genus Pyrops Spinola (Hemiptera: Fulgoridae) from China with description of a new species. Entomotaxonomia 2018, 40, 296–309. [Google Scholar] [CrossRef]
- Wang, G.Y.; Huang, J.; Huang, B.K. Studies on the biology of Fulgora candelaria (L.) (Homoptera: Fulgoridae). Entomol. J. East China 2000, 9, 61–65. (In Chinese) [Google Scholar]
- Constant, J. Review of the effusus group of the lanternfly genus Pyrops Spinola, 1839, with one new species and notes on trophobiosis (Hemiptera: Fulgoromorpha: Fulgoridae). Eur. J. Taxon. 2015, 128, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-S.; Liao, J.-R.; Shiao, S.-F.; Ko, C.-C. Origin and potential expansion of the invasive longan lanternfly, Pyrops candelaria (Hemiptera: Fulgoridae) in Taiwan. Biology 2021, 10, 678. [Google Scholar] [CrossRef]
- Constant, J.; Phauk, S.; Bourgoin, T. Updating lanternflies biodiversity knowledge in Cambodia (Hemiptera: Fulgoromorpha: Fulgoridae) by optimizing field work surveys with citizen science involvement through Facebook networking and data access in FLOW website. Belg. J. Entomol. 2016, 37, 1–16. [Google Scholar]
- Bourgoin, T. FLOW (Fulgoromorpha Lists on The Web): A World Knowledge Base Dedicated to Fulgoromorpha. Version 8, Updated. Available online: http://hemiptera-databases.org/flow/ (accessed on 22 October 2021).
- Kim, J.G.; Lee, E.-H.; Seo, Y.-M.; Kim, N.-Y. Cyclic behavior of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae) on host plants. J. Insect Behav. 2011, 24, 423–435. [Google Scholar] [CrossRef]
- Kerchaw, J.C.W.; Kirkaldy, G.W. A memoir on the anatomy and life-history of the homopterous insect Pyrops candelaria (or “candle-fly”). Zool. Jahrb. Abt. Syst. 1910, 29, 105–124. [Google Scholar]
- Wang, C.H.; Wu, I.H.; Lu, C.N. Population Monitoring of Pyrops Watanabei after Its Removal from the Protected Species List; Project Report No. 100-30; Forestry Bureau: Taipei, Taiwan, 2011; 83p. (In Chinese)
- Constant, J.; Pham, H.-T. Review of the clavatus group of the lanternfly genus Pyrops (Hemiptera: Fulgoromorpha: Fulgoridae). Eur. J. Taxon. 2017, 305, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.-R.; Liu, J.-J.; Li, X.-Y.; Liang, A.-P.; Bourgoin, T. Relating antennal sensilla diversity and possible species behavior in the planthopper pest Lycorma delicatula (Hemiptera: Fulgoromorpha: Fulgoridae). PLoS ONE 2018, 13, e0194995. [Google Scholar] [CrossRef] [Green Version]
- Jubinsky, G. Chinese tallow gets worse! Palmetto 1993, 13, 3. [Google Scholar]
- Russell, L.H.; Schwartz, W.L.; Dollahite, J.W. Toxicity of Chinese tallow tree (Sapium sebiferum) for ruminants. Am. J. Vet. Res. 1969, 30, 1233–1238. [Google Scholar] [PubMed]
- Seip, E.; Ott, H.; Hecker, E. Skin irritant and tumor promoting diterpene esters of the tigliane type from the Chinese tallow tree (Sapium sebiferum). Planta Med. 1983, 49, 199–203. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Locations | Description | Major Plant Species | Latitude, Longitude |
|---|---|---|---|
| Guizikeng, Beitou | A park on a hill | Longan, pomelo, and Triadica sebifera | 25.152, 121.493 |
| Hsing Tian Kong, Beitou Branch | A park of a temple on a hillside | Longan and Acacia confusa | 25.138, 121.477 |
| Chungho Temple, Beitou | A trail on a hill | Longan and Sapium discolor | 25.131, 121.508 |
| Donghua Park, Beitou | A park on a hillside | Acacia confusa | 25.121, 121.508 |
| Zhoumei Village, Beitou | Lowland farm area | Longan and Triadica sebifera | 25.121, 121.509 |
| Shalaobie Park, Beitou | A park on a hillside | Triadica sebifera | 25.142, 121.459 |
| Beitou park, Beitou | A park on a hillside, by the road | Triadica sebifera | 25.137, 121.508 |
| Danfeng Mountain, Beitou | A trail on the mountain at an altitude ca. 120 m | Sapium discolor | 25.133, 121.513 |
| Grand Hotel, Shulin | A park of the hotel on a hillside | Longan and Triadica sebifera | 25.081, 121.528 |
| Bali Junior High School, Bali | A graveyard near the school on a hill | Longan and Triadica sebifera | 25.136, 121.405 |
| Hongshulin Station, Tamsui | A trail by the Metro line on lowland area | Longan and Triadica sebifera | 25.152, 121.459 |
| Jiantan Park, Shulin | By the road, on the foot of the hill | Longan and Triadica sebifera | 25.080, 121.525 |
| Plant Species or Objects | Family | n a | No. Egg Masses |
|---|---|---|---|
| Acacia confusa | Fabaceae | 13 | 15 |
| Ficus subpisocarpa | Moraceae | 2 | 6 |
| Celtis sinensis | Cannabaceae | 1 | 4 |
| Triadica sebifera | Euphorbiaceae | 3 | 3 |
| Wood b | - | 3 | 3 |
| Ficus microcarpa | Moraceae | 2 | 2 |
| Ardisia sieboldii | Primulaceae | 1 | 1 |
| Koelreuteria henryi | Spindaceae | 1 | 1 |
| Alocasia odora | Araceae | 1 | 1 |
| Lagerstroemia subcostata | Lythraceae | 1 | 1 |
| Semecarpus gigantifolius | Anacardiaceae | 1 | 1 |
| Millettia pinnata | Fabaceae | 1 | 1 |
| Dalbergia sissoo | Fabaceae | 1 | 1 |
| Morus australis | Moraceae | 1 | 1 |
| Cordyline fruticosa | Asparagaceae | 1 | 1 |
| Dimocarpus longan | Spindaceae | 1 | 1 |
| Eucalyptus robusta | Myrtaceae | 1 | 1 |
| Machilus zuihoensis | Lauraceae | 1 | 1 |
| Machilus thunbergii | Lauraceae | 1 | 1 |
| Macaranga tanarius | Euphorbiaceae | 1 | 1 |
| Cinnamomum camphora | Lauraceae | 1 | 1 |
| Elaeocarpus serratus | Elaeocarpaceae | 1 | 1 |
| Sum | 40 | 49 | |
| Plant Species | Family | n a | No. Nymphs | % | Mea c | Max d | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st Instar | 2nd Instar | 3rd Instar | 4th Instar | 5th Instar | Total b | ||||||
| Triadica sebifera | Euphorbiaceae | 169 | 229 | 271 | 453 | 535 | 325 | 1813 | 78.8 | 10.7 | 133 |
| Acacia confusa | Fabaceae | 36 | 5 | 47 | 57 | 40 | 15 | 164 | 7.1 | 4.2 | 22 |
| Lagerstroemia subcostata | Lythraceae | 13 | 54 | 35 | 29 | 5 | 1 | 124 | 5.4 | 9.5 | 24 |
| Cinnamomum camphora | Lauraceae | 3 | 50 | 3 | 1 | 0 | 0 | 54 | 2.3 | 18.0 | 50 |
| Psidium guajava | Myrtaceae | 6 | 5 | 7 | 19 | 11 | 0 | 42 | 1.8 | 7.0 | 18 |
| Citrus grandis | Rutaceae | 6 | 20 | 8 | 10 | 1 | 1 | 40 | 1.7 | 6.7 | 15 |
| Morus australis | Moraceae | 2 | 4 | 7 | 15 | 0 | 0 | 26 | 1.1 | 13.0 | 21 |
| Callicarpa formosana | Lamiaceae | 3 | 3 | 10 | 1 | 0 | 0 | 14 | 0.6 | 4.7 | 10 |
| Sapium discolor | Euphorbiaceae | 3 | 1 | 2 | 2 | 5 | 1 | 11 | 0.5 | 3.7 | 5 |
| Koelreuteria henryi | Spindaceae | 1 | 0 | 0 | 0 | 0 | 3 | 3 | 0.1 | 3.0 | 3 |
| Breynia officinalis | Euphorbiaceae | 1 | 0 | 1 | 2 | 0 | 0 | 3 | 0.1 | 3.0 | 3 |
| Ficus septica | Moraceae | 1 | 0 | 0 | 2 | 0 | 0 | 2 | 0.1 | 2.0 | 2 |
| Ficus subpisocarpa | Moraceae | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0.0 | 1.0 | 1 |
| Diospyros kaki | Ebenaceae | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0.0 | 1.0 | 1 |
| Celtis sinensis | Cannabaceae | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0.0 | 1.0 | 1 |
| Macaranga tanarius | Euphorbiaceae | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0.0 | 1.0 | 1 |
| Sum | 248 | 371 | 392 | 591 | 597 | 349 | 2300 | - | - | - | |
| Month | No. Egg Masses | No. Nymphs | |||||
|---|---|---|---|---|---|---|---|
| 1st Instar | 2nd Instar | 3rd Instar | 4th Instar | 5th Instar | Total a | ||
| April | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| May | 43 | 185 | 32 | 0 | 0 | 0 | 217 |
| June | 1 | 116 | 282 | 399 | 138 | 2 | 937 |
| July | 4 | 70 | 78 | 192 | 459 | 347 | 1146 |
| Sum | 49 | 371 | 392 | 591 | 597 | 349 | 2300 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, M.-H.; Yang, Y.-L.; Wu, M.-L.; Wang, L.-J. Host Plants of the Immature Stages of the Invasive Longan Lanternfly, Pyrops candelaria (L.) (Hemiptera, Fulgoridae) in Taiwan. Insects 2021, 12, 1022. https://doi.org/10.3390/insects12111022
Hsu M-H, Yang Y-L, Wu M-L, Wang L-J. Host Plants of the Immature Stages of the Invasive Longan Lanternfly, Pyrops candelaria (L.) (Hemiptera, Fulgoridae) in Taiwan. Insects. 2021; 12(11):1022. https://doi.org/10.3390/insects12111022
Chicago/Turabian StyleHsu, Meng-Hao, Yueh-Lin Yang, Meng-Ling Wu, and Liang-Jong Wang. 2021. "Host Plants of the Immature Stages of the Invasive Longan Lanternfly, Pyrops candelaria (L.) (Hemiptera, Fulgoridae) in Taiwan" Insects 12, no. 11: 1022. https://doi.org/10.3390/insects12111022
APA StyleHsu, M.-H., Yang, Y.-L., Wu, M.-L., & Wang, L.-J. (2021). Host Plants of the Immature Stages of the Invasive Longan Lanternfly, Pyrops candelaria (L.) (Hemiptera, Fulgoridae) in Taiwan. Insects, 12(11), 1022. https://doi.org/10.3390/insects12111022