
Ultramorphological Comparison of Proboscis and Associated Sensilla of Scotogramma trifolii and Protoschinia scutosa (Lepidoptera: Noctuidae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Sampling
2.2. Scanning Electron Microscopy (SEM)
2.3. Statistical Analysis
2.4. Terminology
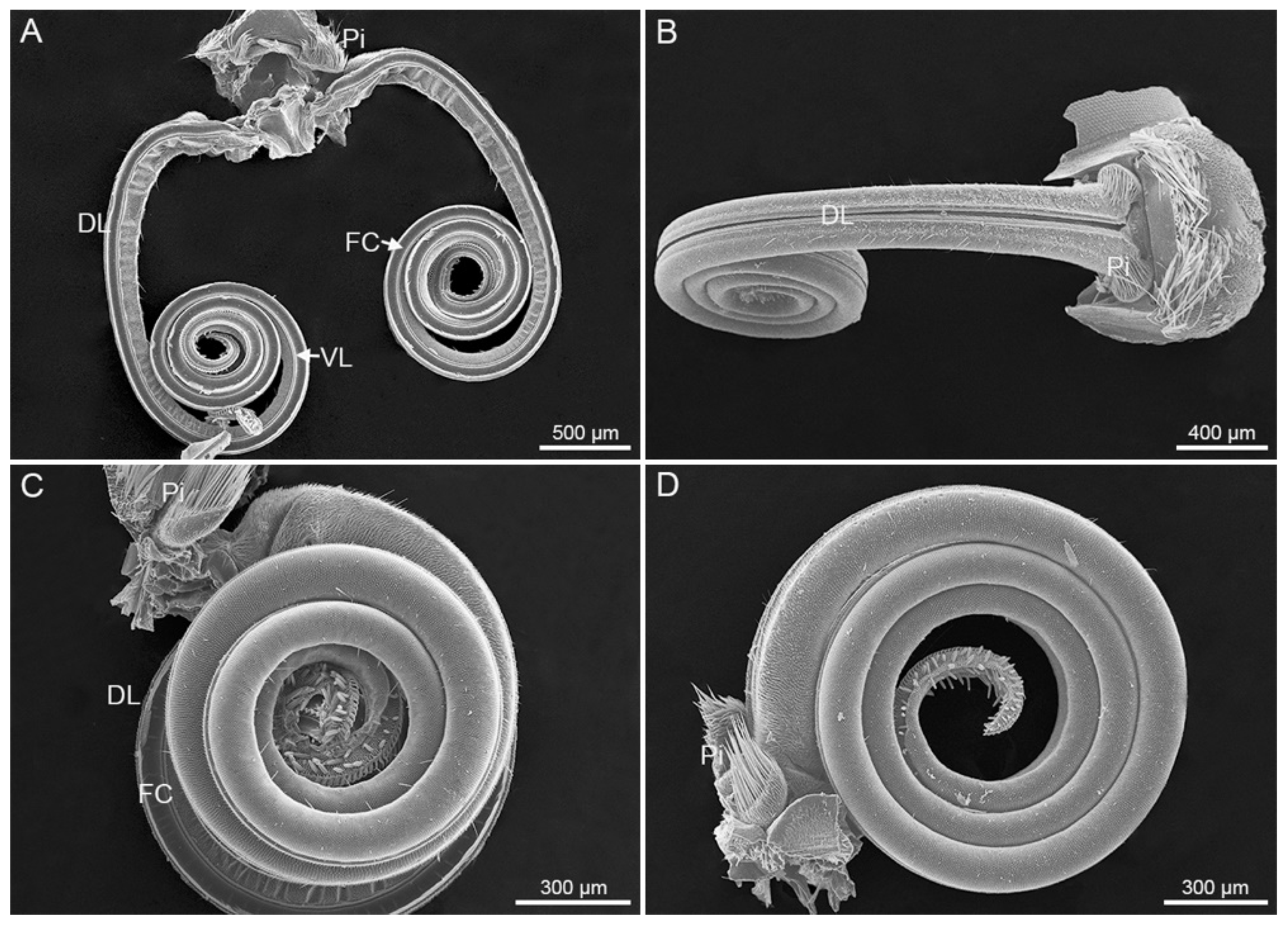
3. Results
3.1. General Morphology of Proboscis
3.1.1. Zone 1
3.1.2. Zone 2
3.1.3. Zone 3
3.2. Proboscis Sensilla
3.2.1. Sensilla Chaetica
3.2.2. Sensilla Basiconica
3.2.3. Sensilla Styloconica
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krenn, H.W. Feeding mechanisms of adult Lepidoptera: Structure, function, and evolution of the mouthparts. Annu. Rev. Entomol. 2010, 55, 307–327. [Google Scholar] [CrossRef] [Green Version]
- Blanke, A.; Ruhr, P.T.; Mokso, R.; Villanueva, P.; Wilde, F.; Stampanoni, M.; Uesugi, K.; Machida, R.; Misof, B. Structural mouthpart interaction evolved already in the earliest lineages of insects. Proc. Biol. Sci. 2015, 282, 20151033. [Google Scholar] [CrossRef] [Green Version]
- Krenn, H.W. Form and function of insect mouthparts. In Insect Mouthparts: Form, Function, Development and Performance; Krenn, H.W., Ed.; Springer Nature: Gewerbestrasse, Cham, Switzerland, 2019; pp. 9–46. [Google Scholar]
- Krenn, H.W.; Fournel, J.; Bauder, J.A.; Hugel, S. Mouthparts and nectar feeding of the flower visiting cricket Glomeremus orchidophilus (Gryllacrididae). Arthropod Struct. Dev. 2016, 45, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Betz, O.; Thayer, M.K.; Newton, A.F. Comparative morphology and evolutionary pathways of the mouthparts in spore–feeding Staphylinoidea (Coleoptera). Acta Zool. 2003, 84, 179–238. [Google Scholar] [CrossRef]
- Lei, G.; Fu, Y.; Wu, W. Fine structure of mouthparts and forelegs of Euplatypus parallelus (Coleoptera: Curculionidae) with emphasis on the behavior of gallery excavation. Micron 2020, 130, 102815. [Google Scholar] [CrossRef]
- Moon, M.J.; Park, J.G.; Kim, K.H. Fine structure of the mouthparts in the ambrosia beetle Platypus koryoensis (Coleoptera: Curculionidae: Platypodinae). Anim. Cells Syst. 2008, 12, 101–108. [Google Scholar] [CrossRef]
- Krenn, H.W.; Aspock, H. Form, function and evolution of the mouthparts of blood-feeding Arthropoda. Arthropod Struct. Dev. 2012, 41, 101–118. [Google Scholar] [CrossRef]
- Wang, T.; Pan, L.; Zhang, Y.; Dai, W. Morphology of the mouthparts of the spittlebug Philagra albinotata Uhler (Hemiptera: Cercopoidea: Aphrophoridae). Arthropod Struct. Dev. 2015, 44, 121–130. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Wang, W.; Brozek, J.; Dai, W. Unique fine morphology of mouthparts in Haematoloecha nigrorufa (Stal) (Hemiptera: Reduviidae) adapted to millipede feeding. Insects 2020, 11, 386. [Google Scholar] [CrossRef] [PubMed]
- Krenn, H.W.; Mauss, V.; Plant, J. Evolution of the suctorial proboscis in pollen wasps (Masarinae, Vespidae). Arthropod Struct. Dev. 2002, 31, 103–120. [Google Scholar] [CrossRef]
- Chen, Q.X.; Li, W.L.; Chen, Y.W.; Chen, J.; Song, Y.Q. Morphological comparison of proboscides and associated sensilla of Helicoverpa armigera and Mythimna separate (Lepidoptera: Noctuidae). Arthropod Struct. Dev. 2019, 49, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Krenn, H.W.; Kristensen, N.P. Early evolution of the proboscis of Lepidoptera (Insecta): External morphology of the galea in basal glossatan moth lineages, with remarks on the origin of the pilifers. Zool. Anz. 2000, 239, 179–196. [Google Scholar]
- Bauder, J.A.; Handschuh, S.; Metscher, B.D.; Krenn, H.W. Functional morphology of the feeding apparatus and evolution of proboscis length in metalmark butterflies (Lepidoptera: Riodinidae). Biol. J. Linn. Soc. 2013, 110, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Zenker, M.M.; Penz, C.; Paris, M.; Specht, A. Proboscis morphology and its relationship to feeding habits in noctuid moths. J. Insect Sci. 2011, 11, 1–10. [Google Scholar] [CrossRef]
- Faucheux, M.J. Sensillum types on the proboscis of the Lepidoptera: A review. Ann. Soc. Entomol. Fr. 2013, 49, 73–90. [Google Scholar] [CrossRef]
- Krenn, H.W.; Plant, J.D.; Szucsich, N.U. Mouthparts of flower-visiting insects. Arthropod Struct. Dev. 2005, 34, 1–40. [Google Scholar] [CrossRef]
- Xue, S.; Hu, Y.Q.; Hua, B.Z. Morphological comparison of proboscis sensilla between Plusiinae and Noctuinae (Lepidoptera: Noctuidae). Zool. Anz. 2016, 263, 75–83. [Google Scholar] [CrossRef]
- Xue, S.; Hua, B.Z. Proboscis sensilla of the black cutworm Agrotis ypsilon (Rottemberg) (Lepidoptera: Noctuidae). J. Asia–Pac. Entomol. 2014, 17, 295–301. [Google Scholar] [CrossRef]
- Hu, G.L.; Zhang, C.M.; Wang, Z.Q.; Chen, Q.X.; Lu, J.Q. Sensilla of the antenna and proboscis of Athetis lepigone (Möschler) (Lepidoptera: Noctuidae). J. Morphol. 2021, 282, 733–745. [Google Scholar] [CrossRef]
- Krenn, H.W.; Zulka, K.P.; Gatschnegg, T. Proboscis morphology and food preferences in nymphalid butterflies (Lepidoptera: Nymphalidae). J. Zool. 2001, 254, 17–26. [Google Scholar] [CrossRef]
- Lehnert, M.S.; Beard, C.E.; Gerard, P.D.; Kornev, K.G.; Adler, P.H. Structure of the Lepidopteran proboscis in relation to feeding guild. J. Morphol. 2016, 277, 167–182. [Google Scholar] [CrossRef]
- Molleman, F.; Krenn, H.W.; Van Alphen, M.E.; Brakefield, P.M.; Devries, P.J.; Zwaan, B.J. Food intake of fruit-feeding butterflies: Evidence for adaptive variation in proboscis morphology. Biol. J. Linn. Soc. 2005, 86, 333–343. [Google Scholar] [CrossRef]
- Kramer, V.R.; Reiter, K.E.; Lehnert, M.S. Proboscis morphology suggests reduced feeding abilities of hybrid Limenitis butterflies (Lepidoptera: Nymphalidae). Biol. J. Linn. Soc. 2018, 125, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.; Mitter, C.; Regier, J.C. Systematics and evolution of the cutworm moths (Lepidoptera: Noctuidae): Evidence from two protein-coding nuclear genes. Syst. Entomol. 2005, 31, 21–46. [Google Scholar] [CrossRef]
- Nieukerken, E.J.; Kaila, L.; Kitching, I.J.; Kristensen, N.P.; Lees, D.C.; Minet, J.; Mitter, C.; Mutanen, M.; Regier, J.C.; Simonsen, T.J.; et al. Order Lepidoptera Linnaeus, 1758. In: Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa 2011, 3148, 212–221. [Google Scholar] [CrossRef]
- Federici, B.A. A new type of insect pathogen in larvae of the clover cutworm, Scotogramma trifolii. J. Invertebr. Pathol. 1982, 40, 41–54. [Google Scholar] [CrossRef]
- Zhao, S.; Fu, X.; Guo, J.; Zhou, Y.; Wyckhuys, K.A.G.; Wu, K. Seasonal patterns of Protoschinia scutosa (Lepidoptera: Noctuidae) migration across China’s Bohai strait. Environ. Entomol. 2018, 47, 927–934. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Chen, L.; Cheng, D.F.; Zhang, Y.J.; Jiang, Y.Y.; Jiang, J.W. Radar observation and population analysis on the migration of the clover cutworm, Scotogramma trifolii Rottemberg (Lepidoptera: Pyralidae). Acta Entomol. Sin. 2007, 50, 494–500. [Google Scholar]
- Cao, K.L.; Wang, S.Y.; Yu, B.J.; Liu, C.C.; Jashenko, R.; Ji, R. Entomological radar observation of migratory insects in China–Kazakhstan border areas. Plant Prot. 2020, 46, 30–37. [Google Scholar]
- Sun, Z.H.; Zhao, S.Z. Occurrence of clover cutworm and its control. J. Neimenggu Sci. Technol. 1995, 2, 23–24. [Google Scholar]
- Yu, J.N.; Bao, Y.Q. The occurrence of clover cutworm in cotton region of Xinjiang. Xinjiang Agric. Sci. 1996, 1, 34. [Google Scholar]
- Zhao, Z.J.; Chen, E.X.; Zhang, Y. Studies on the biological characteristic of Scotogramma trifolii (Rott) and its control. Zhongguo Tiancai 1992, 25–28. [Google Scholar]
- Li, Q.R.; Li, F.G.; Wei, Y.H.; Zhu, H.X.; Lai, Y.P.; Hou, L.; Guo, Q.Y. Checklist of insect pests, their enemies and description of five species of pests in Chenopodium quinoa Willd. in arid area of Qinghai Plateau. Plant Prot. 2019, 45, 190–198. [Google Scholar]
- Lehnert, M.S.; Monaenkova, D.; Andrukh, T.; Beard, C.E.; Adler, P.H.; Kornev, K.G. Hydrophobic-hydrophilic dichotomy of the butterfly proboscis. J. R. Soc. Interface 2013, 10, 20130336. [Google Scholar] [CrossRef]
- Krenn, H.W. Functional morphology and movements of the proboscis of Lepidoptera (Insecta). Zoomorphology 1990, 110, 105–114. [Google Scholar] [CrossRef]
- Bauder, J.A.S.; Karolyi, F. Superlong proboscises as co-adaptations to flowers. In Insect Mouthparts: Form, Function, Development and Performance; Krenn, H.W., Ed.; Springer Nature: Gewerbestrasse, Cham, Switzerland, 2019; pp. 479–527. [Google Scholar]
- Krenn, H.W.; Kristensen, N.P. Evolution of proboscis musculature in Lepidoptera. Eur. J. Entomol. 2004, 101, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Wannenmacher, G.; Wasserthal, L.T. Contribution of the maxillary muscles to proboscis movement in hawkmoths (Lepidoptera: Sphingidae)-an electrophysiological study. J. Insect Physiol. 2003, 49, 765–776. [Google Scholar] [CrossRef]
- Hepburn, H.R. Proboscis extension and recoil in Lepidoptera. J. Insect Physiol. 1971, 17, 637–656. [Google Scholar] [CrossRef]
- Krenn, H.W. Proboscis musculature in the butterfly Vanessa cardui (Nymphalidae, Lepidoptera): Settling the proboscis recoiling controversy. Acta Zool. (Stockh.) 2001, 81, 259–266. [Google Scholar] [CrossRef]
- Zhang, C.; Beard, C.E.; Adler, P.H.; Kornev, K.G. Effect of curvature on wetting and dewetting of proboscises of butterflies and moths. R. Soc. Open Sci. 2018, 5, 171241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehnert, M.S.; Wei, Q.H. Hierarchical microstructures and functions of the Lepidopteran proboscis cuticle. In Insect Mouthparts: Form, Function, Development and Performance; Krenn, H.W., Ed.; Springer Nature: Gewerbestrasse, Cham, Switzerland, 2019; pp. 315–334. [Google Scholar]
- Wang, N.; Wei, J.S.; Dang, L.; Wang, J.B.; Hua, B.Z. Ultramorphology of sensilla on the proboscis in three noctuid moths (Lepidoptera:Noctuidae). Acta Entomol. Sin. 2012, 55, 877–884. [Google Scholar] [CrossRef]
- Lee, S.C.; Kim, J.H.; Lee, S.J. Adhesion and suction functions of the tip region of a nectar-drinking butterfly proboscis. J. Bionic Eng. 2017, 14, 600–606. [Google Scholar] [CrossRef]
- Tsai, C.C.; Monaenkova, D.; Beard, C.E.; Adler, P.H.; Kornev, K.G. Paradox of the drinking-straw model of the butterfly proboscis. J. Exp. Biol. 2014, 217, 2130–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwauk, K.J.; Hasegawa, D.K.; Lehnert, M.S.; Beard, C.E.; Gerard, P.D.; Kornev, K.G.; Adler, P.H. Drinking with an unsealed tube: Fluid uptake along the butterfly proboscis. Ann. Entomol. Soc. Am. 2014, 107, 886–892. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Daniel, T.L. Mechanics of food handling by fluid-feeding insects. In Regulatory Mechanisms in Insect Feeding; Chapman, R.F., Ed.; Springer: Boston, MA, USA, 1995; pp. 32–73. [Google Scholar]
- Kornev, K.G.; Salamatin, A.A.; Adler, P.H.; Beard, C.E. Structural and physical determinants of the proboscis-sucking pump complex in the evolution of fluid-feeding insects. Sci. Rep. 2017, 7, 6582. [Google Scholar] [CrossRef] [Green Version]
- Salamatin, A.A.; Adler, P.H.; Kornev, K.G. Lepidopteran mouthpart architecture suggests a new mechanism of fluid uptake by insects with long proboscises. J. Theor. Biol. 2021, 510, 110525. [Google Scholar] [CrossRef]
- Monaenkova, D.; Lehnert, M.S.; Andrukh, T.; Beard, C.E.; Rubin, B.; Tokarev, A.; Lee, W.K.; Adler, P.H.; Kornev, K.G. Butterfly proboscis: Combining a drinking straw with a nanosponge facilitated diversification of feeding habits. J. R. Soc. Interface 2012, 9, 720–726. [Google Scholar] [CrossRef] [Green Version]
- Kornev, K.G.; Adler, P.H. Physical determinants of fluid-feeding in insects. In Insect Mouthparts: Form, Function, Development and Performance; Krenn, H.W., Ed.; Springer Nature: Gewerbestrasse, Cham, Switzerland, 2019; pp. 263–314. [Google Scholar]
- Krenn, H.W. Proboscis sensilla in Vanessa cardui (Nymphalidae, Lepidoptera): Functional morphology and significance in flower–probing. Zoomorphology 1998, 118, 23–30. [Google Scholar] [CrossRef]
- Song, Y.Q.; Sun, H.Z.; Wu, J.X. Ultrastructural characteristics of the proboscis and the labial palp pit organ in the oriental fruit moth, Grapholita Molesta. Bull. Insectol. 2016, 69, 59–66. [Google Scholar]
- Walters, B.D.; Albert, P.J.; Zacharuk, R.Y. Morphology and ultrastructure of sensilla on the proboscis of the adult spruce budworm, Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae). Can. J. Zool. 1998, 76, 466–479. [Google Scholar] [CrossRef]
- Inoue, T.A.; Asaoka, K.; Seta, K.; Imaeda, D.; Ozaki, M. Sugar receptor response of the food-canal taste sensilla in a nectar-feeding swallowtail butterfly, Papilio xuthus. Sci. Nat. 2009, 96, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Zaspel, J.M.; Weller, S.J.; Branham, M.A. A comparative survey of proboscis morphology and associated structures in fruit-piercing, tear-feeding, and blood-feeding moths in Calpinae (Lepidoptera: Erebidae). Zoomorphology 2011, 130, 203–225. [Google Scholar] [CrossRef]
- Goldware, M.A.; Barnes, M.M. Mouthparts of the adult codling moth, Laspeyresia pomonella (Lepidoptera: Olethreutidae). Ann. Entomol. Soc. Am. 1973, 66, 349–351. [Google Scholar] [CrossRef]
- Städler, E.; Seabrook, W.D. Chemoreceptors on the proboscis of the female eastern spruce budworm: Electrophysiological study. Entomol. Exp. Appl. 1975, 18, 153–160. [Google Scholar] [CrossRef]
- Jorgensen, K.; Kvello, P.; Almaas, T.J.; Mustaparta, H. Two closely located areas in the suboesophageal ganglion and the tritocerebrum receive projections of gustatory receptor neurons located on the antennae and the proboscis in the moth Heliothis virescens. J. Comp. Neurol. 2006, 496, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Nagnan-Le Meillour, P.; Cain, A.H.; Jacquin-Joly, E.; Francois, M.C.; Ramachandran, S.; Maida, R.; Steinbrecht, R.A. Chemosensory proteins from the proboscis of Mamestra brassicae. Chem. Senses 2000, 25, 541–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaney, W.M.; Simmonds, M.S.J. Food selection in adults and larvae of three species of Lepidoptera: A behavioural and electrophysiological study. Entomol. Exp. Appl. 1988, 49, 111–121. [Google Scholar] [CrossRef]
- Shields, V.D. Fine structure of the galeal styloconic sensilla of larval Lymantria dispar (Lepidoptera: Lymantriidae). Ann. Entomol. Soc. Am. 2009, 102, 1116–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakagami, K.; Matsubara, S.; Sugiura, S. Nocturnal moths as potential pollinators of Marsdenia formosana (Apocynaceae: Asclepiadoideae). Taiwania 2019, 64, 195–197. [Google Scholar] [CrossRef]
- Guo, P.; Wang, G.P.; Jin, L.J.; Fan, X.Q.; He, H.L.; Zhou, P.W.; Guo, X.R.; Li, W.Z.; Yuan, G.H. Identification of summer nectar plants contributing to outbreaks of Mythimna separata (Walker) (Lepidoptera: Noctuidae) in North China. J. Integr. Agric. 2018, 17, 1516–1526. [Google Scholar] [CrossRef] [Green Version]
- Sakagami, K.; Sugiura, S. Noctuid moths as pollinators of Habenaria sagittifera (Orchidaceae): Floral adaptations for the transfer of pollinaria on the thoraxes of moths. Sci. Nat. 2019, 106, 58. [Google Scholar] [CrossRef]
- Yu, J.N.; Zhou, X.H.; Ma, Y.P.; Bao, Y.Q. A study of Scotogramma trifolii and its control. J. Xinjiang Agric. Univ. 1997, 20, 70–73. [Google Scholar]
- Büttiker, W.; Krenn, H.W.; Putterill, J.F. The proboscis of eye-frequenting and piercing Lepidoptera (Insecta). Zoomorphology 1996, 116, 77–83. [Google Scholar] [CrossRef]
- Hilgartner, R.; Raoilison, M.; Büttiker, W.; Lees, D.C.; Krenn, H.W. Malagasy birds as hosts for eye-frequenting moths. Biol. Lett. 2007, 3, 117–120. [Google Scholar] [CrossRef] [Green Version]
- Knopp, M.C.N.; Krenn, H.W. Efficiency of fruit juice feeding in Morpho peleides (Nymphalidae, Lepidoptera). J. Insect Behav. 2003, 16, 67–77. [Google Scholar] [CrossRef]
- Danaher, M.W.; Ward, C.; Zettler, L.W.; Covell, C.V. Pollinia removal and suspected pollination of the endangered ghost orchid, Dendrophylax lindenii (Orchidaceae) by various hawk moths (Lepidoptera: Sphingidae): Another mystery dispelled. Fla. Entomol. 2020, 102, 671. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Dai, W. Fine morphology of the mouthparts in Cheilocapsus nigrescens (Hemiptera: Heteroptera: Miridae) reflects adaptation for phytophagous habits. Insects 2019, 10, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Length/Width (μm) | t-Test | |
|---|---|---|---|
| S. trifolii | P. scutosa | ||
| Proboscis | 7478.07 ± 159.76 (14) | 7858.15 ± 157.38 (14) | NS |
| Food canal | 61.58 ± 0.92 (28) | 59.42 ± 0.66 (28) | * |
| Zone 1 | 6426.14 ± 165.60 (14) | 7157.51 ± 156.61 (14) | * |
| Zone 2 | 1049.68 ± 36.54 (14) | 816.44 ± 25.68 (14) | * |
| Zone 3 | 35.03 ± 1.49 (8) | 39.03 ± 0.65 (14) | * |
| Type | Length (μm) | t-Test | Basal Width (μm) | t-Test | ||
|---|---|---|---|---|---|---|
| S. trifolii | P. scutosa | S. trifolii | P. scutosa | |||
| SC | 46.41 ± 1.93 (140) | 60.20 ± 2.13 (140) | * | 3.23 ± 0.06 (140) | 3.11 ± 0.06 (140) | NS |
| SST | 37.28 ± 0.52 (280) | 30.90 ± 0.48 (280) | * | 8.26 ± 0.09 (280) | 7.50 ± 0.09 (280) | * |
| ESB (Zone 1) | 9.67 ± 0.28 (140) | 6.81 ± 0.21 (140) | * | 2.37 ± 0.04 (140) | 2.15 ± 0.05 (140) | * |
| ESB (Zone 2) | 6.67 ± 0.43 (28) | 4.41 ± 0.30 (28) | * | 2.14 ± 0.07 (28) | 1.87 ± 0.08 (28) | * |
| ISB | 8.01 ± 0.30 (50) | 6.82 ± 0.29 (50) | * | 2.46 ± 0.05 (50) | 2.49 ± 0.05 (50) | NS |
| Types | Length (μm) | t-Test | Basal Width (μm) | t-Test | ||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | |||
| Proboscis | 7583.81 ± 640.84 (7) | 7372.33 ± 580.87 (7) | NS | – | – | – |
| Zone 1 | 6570.53 ± 253.31 (7) | 6281.75 ± 218.44 (7) | NS | – | – | – |
| Zone 2 | 1008.78 ± 46.84 (7) | 1090.58 ± 55.07 (7) | NS | – | – | – |
| Zone 3 | 33.67 ± 2.01 (4) | 36.39 ± 2.24 (4) | NS | – | – | – |
| SC | 44.24 ± 3.33 (70) | 48.58 ± 2.82 (70) | NS | 3.44 ± 0.09 (70) | 3.03 ± 0.06 (70) | * |
| SST | 36.43 ± 0.63 (140) | 38.13 ± 0.82 (140) | NS | 9.10 ± 0.13 (140) | 7.43 ± 0.08 (140) | * |
| ESB (Zone 1) | 8.99 ± 0.24 (70) | 10.36 ± 0.49 (70) | * | 2.50 ± 0.06 (70) | 2.24 ± 0.05 (70) | * |
| ESB (Zone 2) | 6.39 ± 0.39 (14) | 6.94 ± 0.78 (14) | NS | 2.27 ± 0.08 (14) | 2.01 ± 0.10 (14) | NS |
| ISB | 7.54 ± 0.46 (25) | 8.49 ± 0.36 (25) | NS | 2.64 ± 0.07 (25) | 2.28 ± 0.06 (25) | * |
| Types | Length (μm) | t-Test | Basal Width (μm) | t-Test | ||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | |||
| Proboscis | 8065.79 ± 195.46 (7) | 7650.50 ± 234.00 (7) | NS | – | – | – |
| Zone 1 | 7362.79 ± 186.98 (7) | 6952.23 ± 239.31 (7) | NS | – | – | – |
| Zone 2 | 865.20 ± 39.11 (7) | 767.67 ± 23.16 (7) | NS | – | – | – |
| Zone 3 | 38.83 ± 0.64 (7) | 39.24 ± 1.18 (7) | NS | – | – | – |
| SC | 59.02 ± 3.19 (70) | 61.38 ± 2.83 (70) | NS | 3.27 ± 0.09 (70) | 2.94 ± 0.09 (70) | * |
| SST | 31.16 ± 0.65 (140) | 30.64 ± 0.72 (140) | NS | 7.82 ± 0.13 (140) | 7.18 ± 0.13 (140) | * |
| ESB (Zone 1) | 6.54 ± 0.27 (70) | 7.16 ± 0.31 (70) | NS | 2.22 ± 0.07 (70) | 2.10 ± 0.07 (70) | NS |
| ESB (Zone 2) | 4.28 ± 0.40 (14) | 4.54 ± 0.46 (14) | NS | 1.89 ± 0.1 (14) | 1.86 ± 0.09 (14) | NS |
| ISB | 6.80 ± 0.45 (25) | 6.85 ± 0.39 (25) | NS | 2.48 ± 0.09 (25) | 2.49 ± 0.07 (25) | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.-M.; Niu, Y.; Hu, G.-L.; Lu, J.-Q. Ultramorphological Comparison of Proboscis and Associated Sensilla of Scotogramma trifolii and Protoschinia scutosa (Lepidoptera: Noctuidae). Insects 2021, 12, 992. https://doi.org/10.3390/insects12110992
Zhang C-M, Niu Y, Hu G-L, Lu J-Q. Ultramorphological Comparison of Proboscis and Associated Sensilla of Scotogramma trifolii and Protoschinia scutosa (Lepidoptera: Noctuidae). Insects. 2021; 12(11):992. https://doi.org/10.3390/insects12110992
Chicago/Turabian StyleZhang, Chuan-Min, Yue Niu, Gui-Lin Hu, and Ji-Qi Lu. 2021. "Ultramorphological Comparison of Proboscis and Associated Sensilla of Scotogramma trifolii and Protoschinia scutosa (Lepidoptera: Noctuidae)" Insects 12, no. 11: 992. https://doi.org/10.3390/insects12110992
APA StyleZhang, C.-M., Niu, Y., Hu, G.-L., & Lu, J.-Q. (2021). Ultramorphological Comparison of Proboscis and Associated Sensilla of Scotogramma trifolii and Protoschinia scutosa (Lepidoptera: Noctuidae). Insects, 12(11), 992. https://doi.org/10.3390/insects12110992