
Insect Herbivore Populations and Plant Damage Increase at Higher Elevations

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
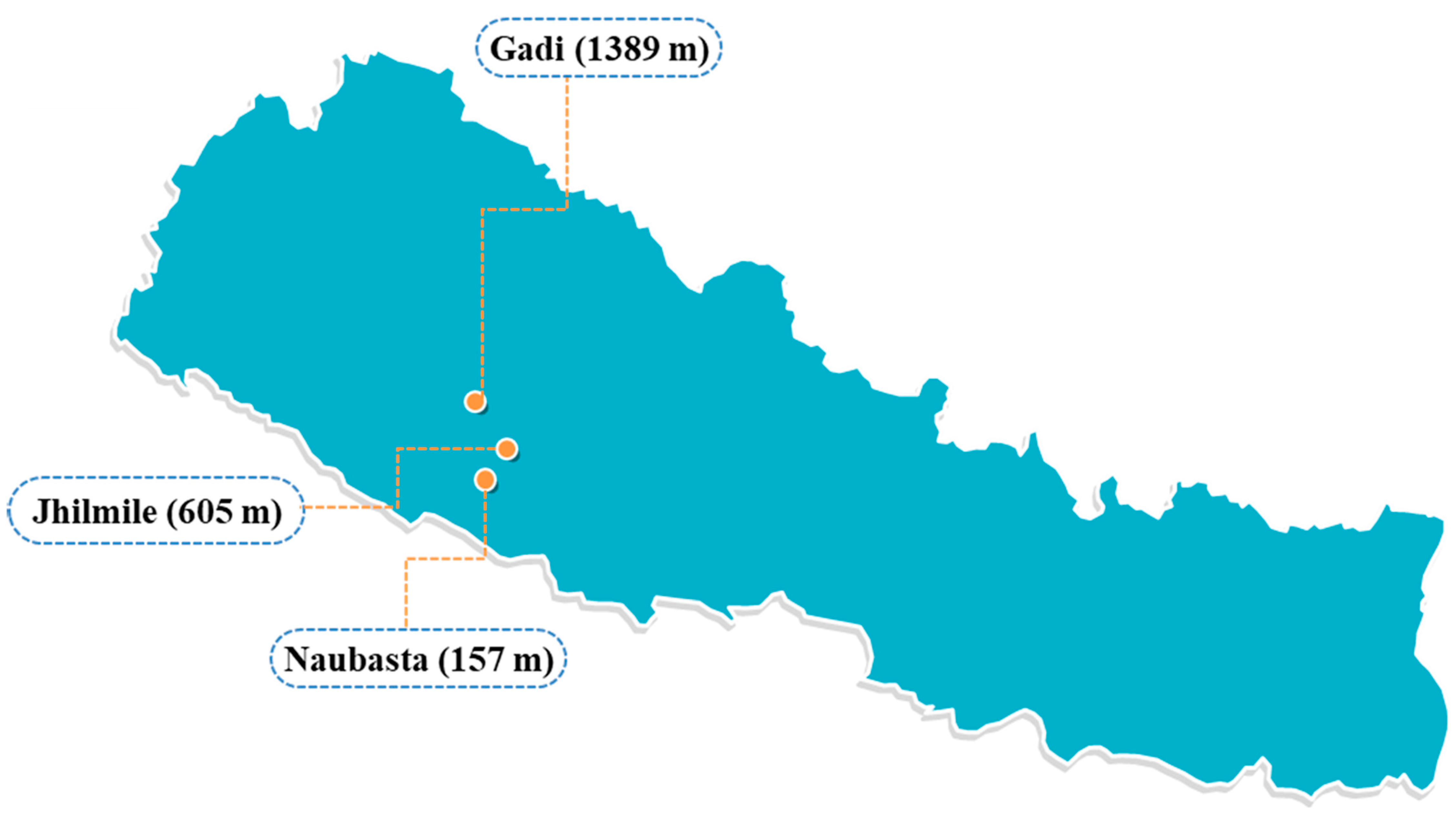
2. Materials and Methods
3. Results
3.1. Temperature Variation with Elevation
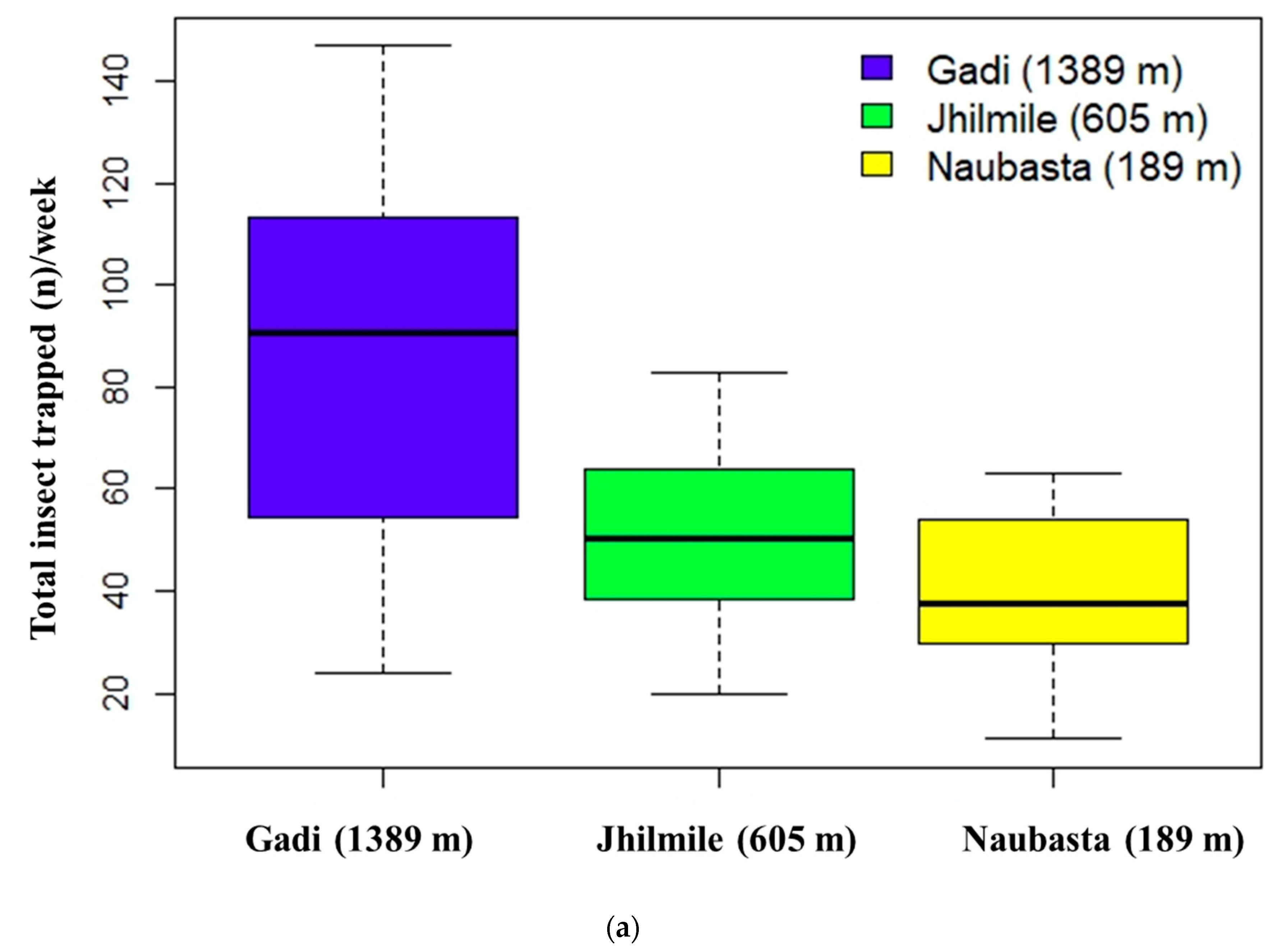
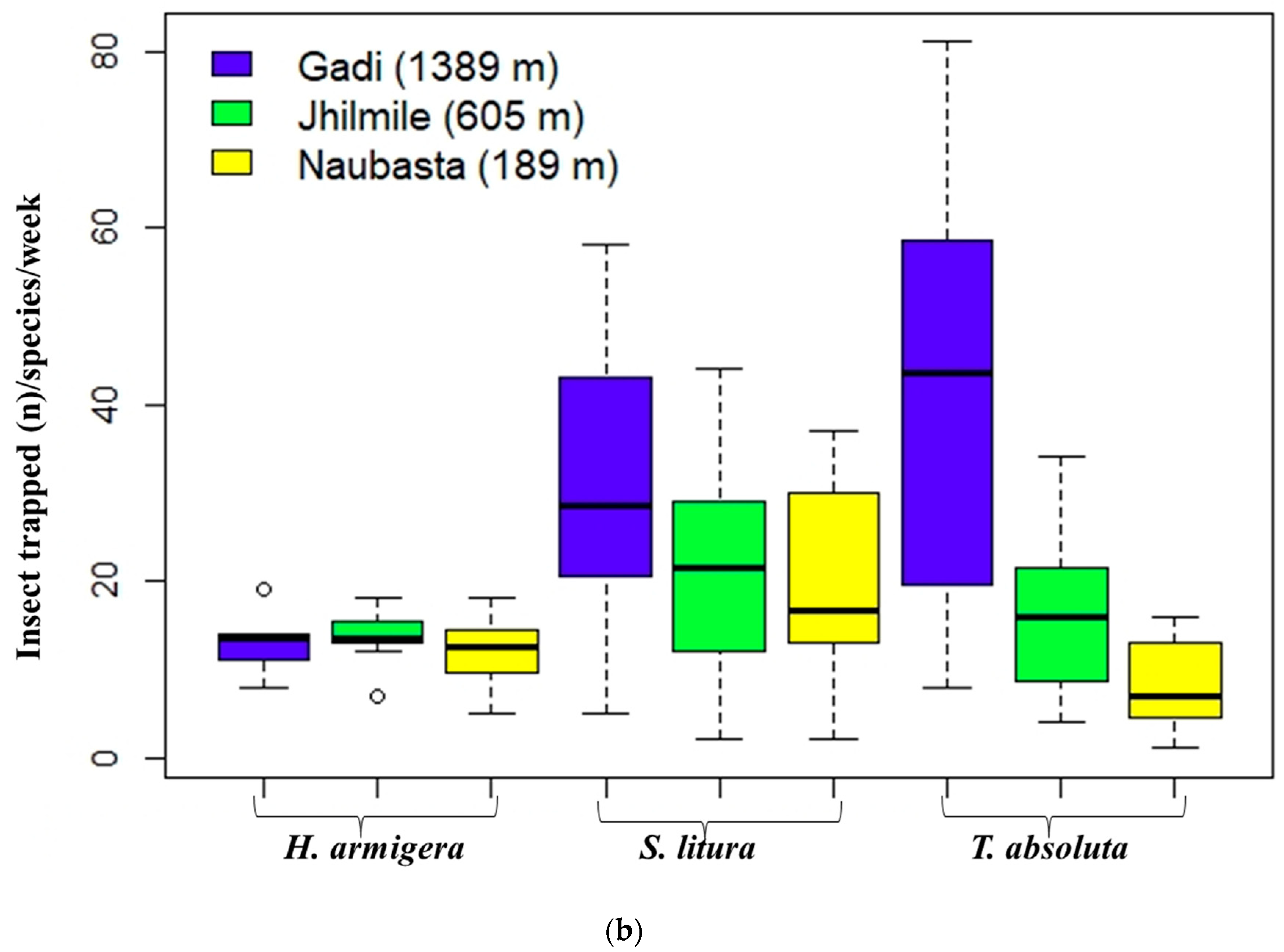
3.2. Insect Herbivore Populations
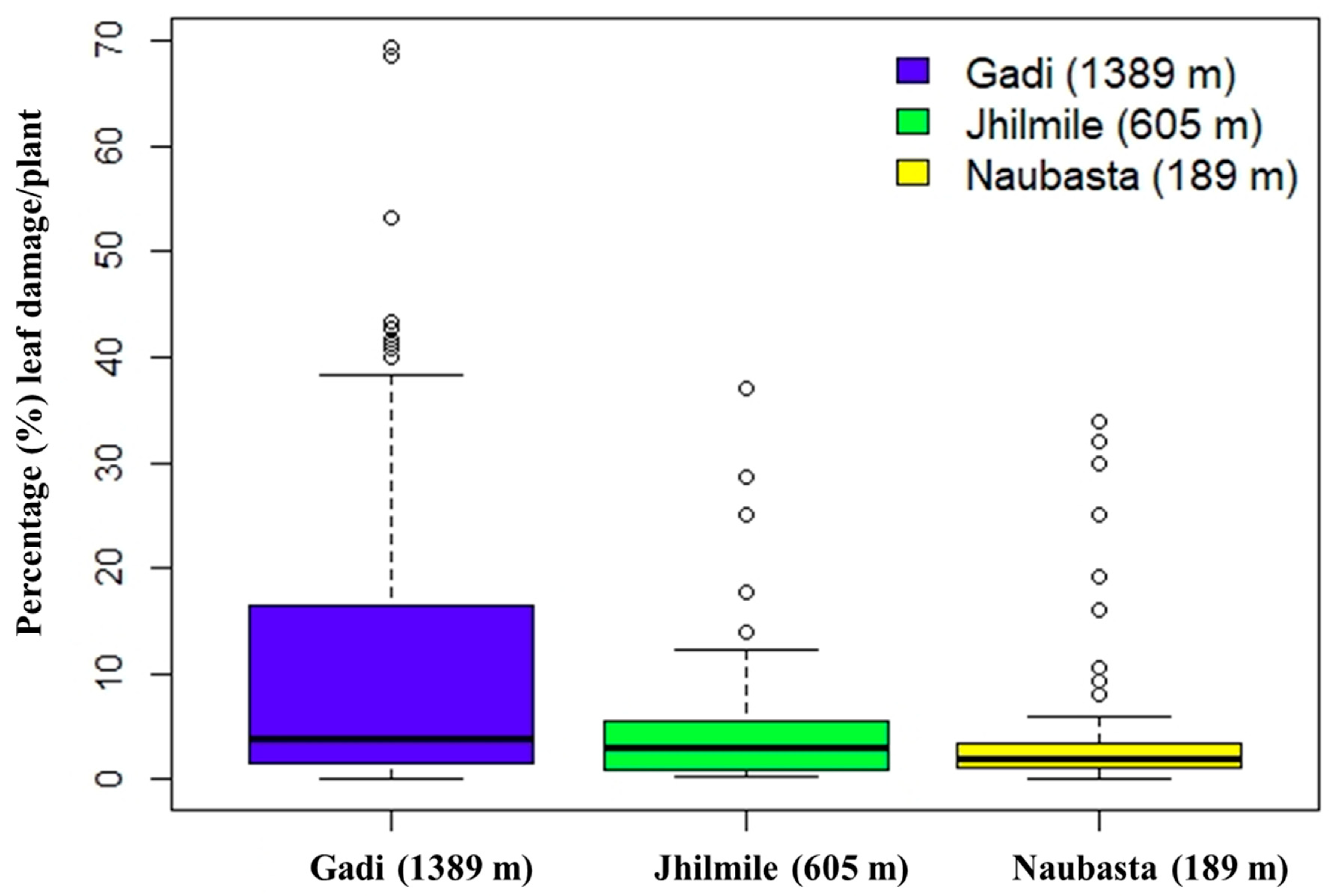
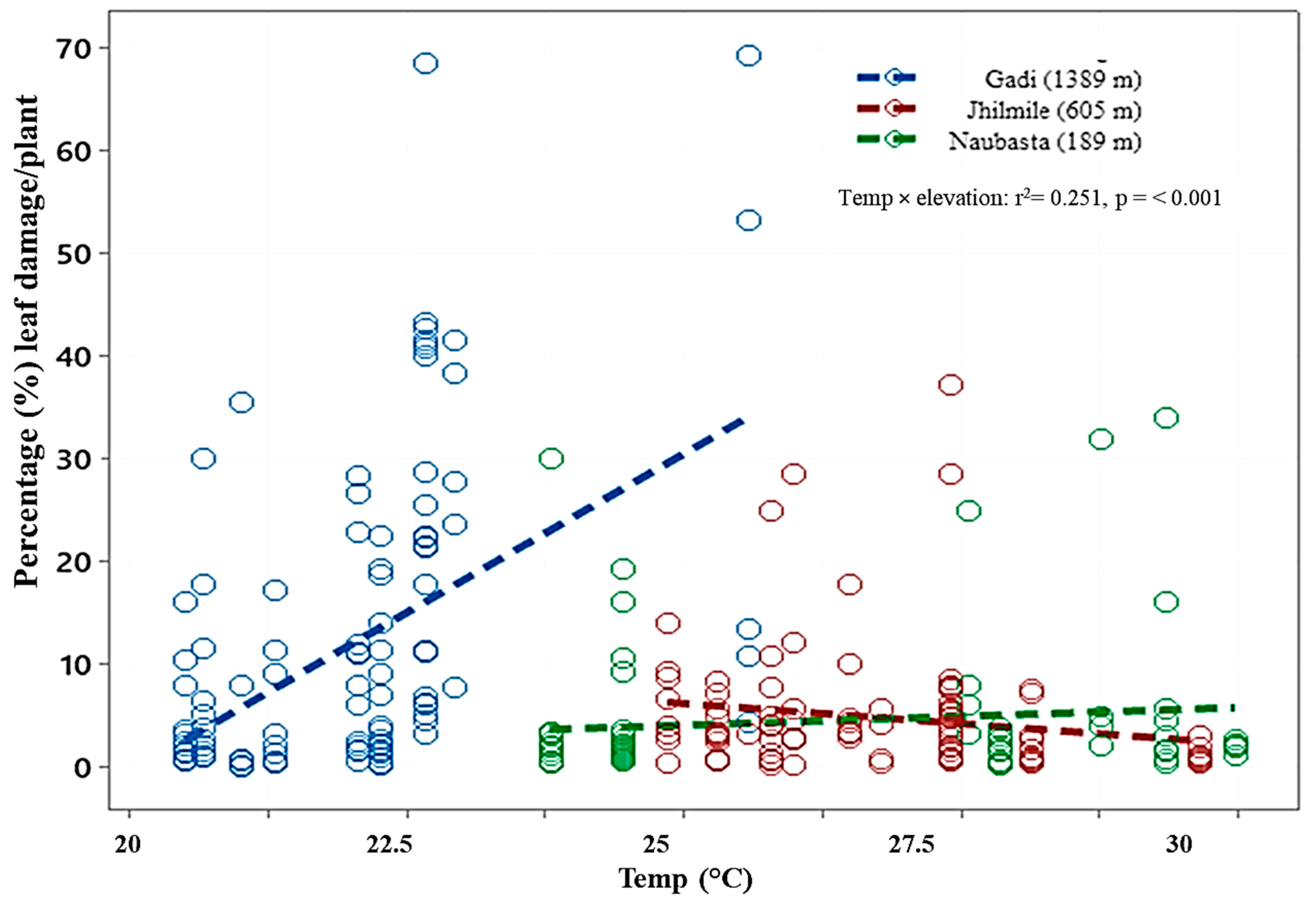
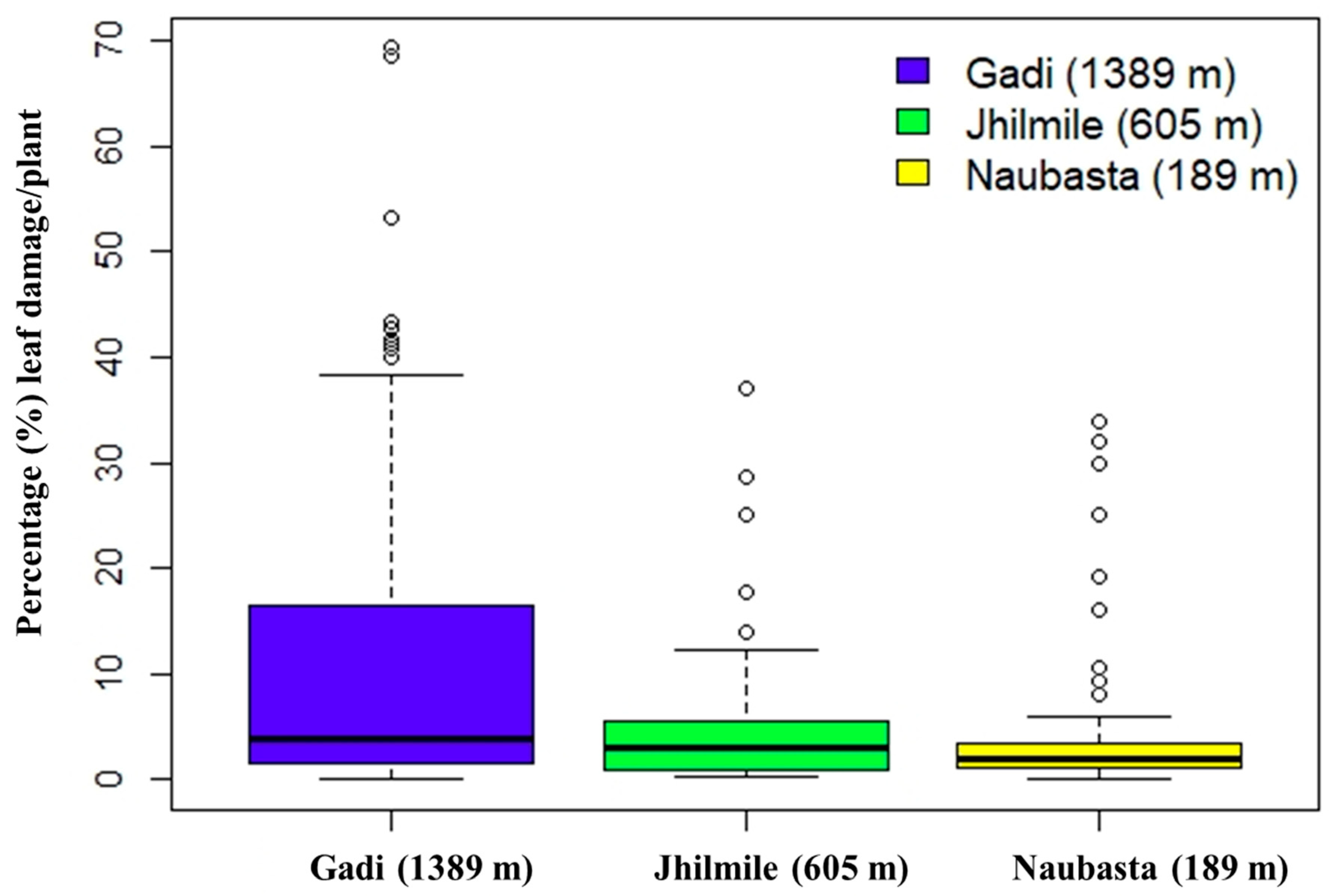
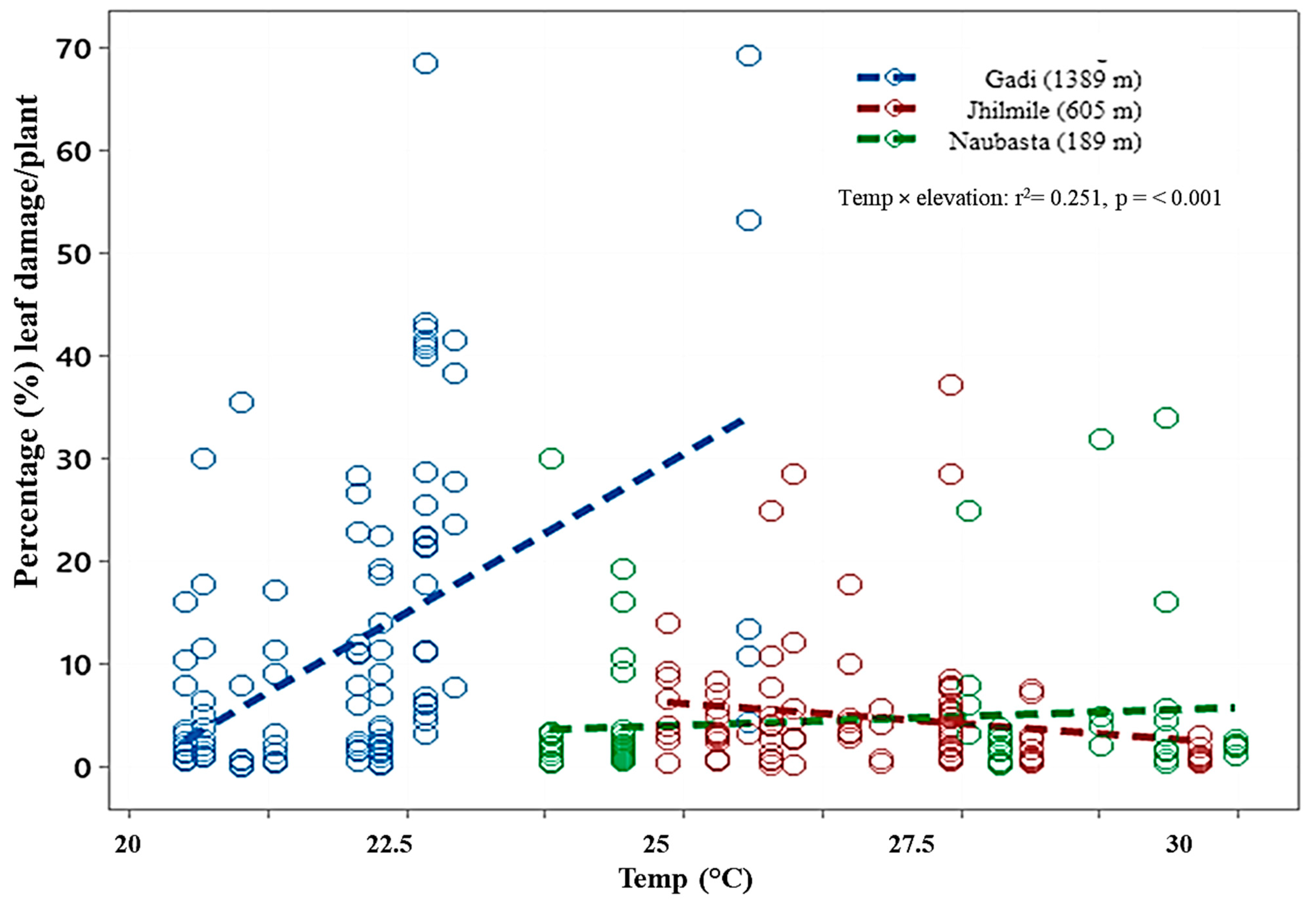
3.3. Leaf Damage from Herbivory
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Körner, C. The use of ‘altitude’in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Buckley, J.; Widmer, A.; Mescher, M.C.; De Moraes, C.M. Variation in growth and defence traits among plant populations at different elevations: Implications for adaptation to climate change. J. Ecol. 2019, 107, 2478–2492. [Google Scholar] [CrossRef]
- Moreira, X.; Petry, W.K.; Mooney, K.A.; Rasmann, S.; Abdala-Roberts, L. Elevational gradients in plant defences and insect herbivory: Recent advances in the field and prospects for future research. Ecography 2018, 41, 1485–1496. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.A.; Beaulieu, J.M. Life history influences rates of climatic niche evolution in flowering plants. Proc. R. Soc. B Biol. Sci. 2009, 276, 4345–4352. [Google Scholar] [CrossRef] [Green Version]
- Smilanich, A.M.; Fincher, R.M.; Dyer, L.A. Does plant apparency matter? Thirty years of data provide limited support but reveal clear patterns of the effects of plant chemistry on herbivores. New Phytol. 2016, 210, 1044–1057. [Google Scholar] [CrossRef] [Green Version]
- Pellissier, L.; Fiedler, K.; Ndribe, C.; Dubuis, A.; Pradervand, J.-N.; Guisan, A.; Rasmann, S. Shifts in species richness, herbivore specialization, and plant resistance along elevation gradients. Ecol. Evol. 2012, 2, 1818–1825. [Google Scholar] [CrossRef]
- Salgado, A.L.; Suchan, T.; Pellissier, L.; Rasmann, S.; Ducrest, A.-L.; Alvarez, N. Differential phenotypic and genetic expression of defence compounds in a plant–herbivore interaction along elevation. R. Soc. Open Sci. 2016, 3, 160226. [Google Scholar] [CrossRef] [Green Version]
- Dostdlek, T.; Rokaya, M.B.; Marsik, P.; Rezek, J.; Skuhrovec, J.; Pavela, R.; Munzbergova, Z. Trade-off among different anti-herbivore defence strategies along an altitudinal gradient. AoB Plants 2016, 8, plw026. [Google Scholar] [CrossRef] [Green Version]
- Talebi, S.M.; Nohooji, M.G.; Yarmohammadi, M.; Khani, M.; Matsyura, A. Effect of altitude on essential oil composition and on glandular trichome density in three Nepeta species (N. sessilifolia, N. heliotropifolia and N. fissa). Mediterr. Bot. 2019, 40, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Pratt, J.D.; Mooney, K.A. Clinal adaptation and adaptive plasticity in Artemisia californica: Implications for the response of a foundation species to predicted climate change. Glob. Chang. Biol. 2013, 19, 2454–2466. [Google Scholar] [CrossRef]
- Wang, Q.; Fan, X.; Wang, M. Recent warming amplification over high elevation regions across the globe. Clim. Dyn. 2014, 43, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Galmán, A.; Petry, W.K.; Abdala-Roberts, L.; Butron, A.; De La Fuente, M.; Francisco, M.; Kergunteuil, A.; Rasmann, S.; Moreira, X. Inducibility of chemical defences in young oak trees is stronger in species with high elevational ranges. Tree Physiol. 2018, 39, 606–614. [Google Scholar] [CrossRef]
- Dyoulgerov, M.; Bucher, A.; Zermoglio, F.; Forner, C. Climate Risk and Adaptation Country Profile. Nepal: Vulnerability, Risk Reduction and Adaptation to Climate Change. Global Facility for Disaster Reduction and Recovery; World Bank: Washington, DC, USA, 2011; Available online: https://www.gfdrr.org/en/publication/climate-risk-and-adaptation-country-profile-nepal (accessed on 15 January 2018).
- Ghimire, D.; Lamsal, G.; Paudel, B.; Khatri, S.; Bhusal, B. Analysis of trend in area, production and yield of major vegetables of Nepal. Trends Hortic. 2018. [Google Scholar] [CrossRef]
- Acharya, R.; Paudel, S.; Rajbhandari, B.P. Growth, yield formation, fruit quality and economics of tomato production under IPM package vs. conventional practice in Surkhet district. Nepal. J. Agric. Sci. 2015, 13, 96–102. [Google Scholar]
- Paudel, S.; Sah, L.P.; Pradhan, K.; Colavito, L.A.; Upadhyay, B.P.; Rajotte, E.G.; Muniappan, R. Development and Dissemination of Vegetable IPM Practices and Packages in Nepal. In Integrated Pest Management of Tropical Vegetable Crops; Springer: Dordrecht, The Netherlands, 2016; pp. 251–269. [Google Scholar]
- Venkatramanan, S.; Wu, S.; Shi, B.; Marathe, A.; Eubank, S.; Sah, L.; Giri, A.; Colavito, L.; Nitin, K.; Sridhar, V.; et al. Modeling commodity flow in the context of invasive species spread: Study of Tuta absoluta in Nepal. Crop. Prot. 2020, 135, 104736. [Google Scholar] [CrossRef] [Green Version]
- Kanhar, K.A.; Sahito, H.A.; Kanher, F.M.; Tunio, S.A.; Awan, R.R.H. Damage percent and biological parameters of leaf miner Acrocercops syngramma (Meyrick) on different mango varieties. J. Entomol. Zool. Stud. 2016, 4, 541–546. [Google Scholar]
- Paudel, S.; Sah, L.; Devkota, M.; Poudyal, V.; Prasad, P.; Reyes, M. Conservation Agriculture and Integrated Pest Management Practices Improve Yield and Income while Reducing Labor, Pests, Diseases and Chemical Pesticide Use in Smallholder Vegetable Farms in Nepal. Sustainability 2020, 12, 6418. [Google Scholar] [CrossRef]
- Demétrio, C.G.; Hinde, J.; Moral, R.A. Models for overdispersed data in entomology. In Ecological Modelling Applied to Entomology; Springer: Cham, Germany, 2014; pp. 219–259. [Google Scholar]
- Minitab (Version 18). Available online: http://www.minitab.com/en-US/products/minitab/default.aspx (accessed on 22 June 2020).
- Azrag, A.G.A.; Pirk, C.W.W.; Yusuf, A.A.; Pinard, F.; Niassy, S.; Mosomtai, G.; Babin, R. Prediction of insect pest distribution as influenced by elevation: Combining field observations and temperature-dependent development models for the coffee stink bug, Antestiopsis thunbergii (Gmelin). PLoS ONE 2018, 13, e0199569. [Google Scholar] [CrossRef]
- Sam, K.; Koane, B.; Sam, L.; Mrazova, A.; Segar, S.; Volf, M.; Moos, M.; Simek, P.; Sisol, M.; Novotny, V. Insect herbivory and herbivores of Ficus species along a rain forest elevational gradient in Papua New Guinea. Biotropica 2020, 52, 263–276. [Google Scholar] [CrossRef]
- Rasmann, S.; Pellissier, L.; Defossez, E.; Jactel, H.; Kunstler, G. Climate-driven change in plant-insect interactions along elevation gradients. Funct. Ecol. 2014, 28, 46–54. [Google Scholar] [CrossRef]
- Hodkinson, I.D. Terrestrial insects along elevation gradients: Species and community responses to altitude. Biol. Rev. 2005, 80, 489–513. [Google Scholar] [CrossRef] [Green Version]
- Paudel, S.; Lin, P.-A.; Hoover, K.; Felton, G.W.; Rajotte, E.G. Asymmetric Responses to Climate Change: Temperature Differentially Alters Herbivore Salivary Elicitor and Host Plant Responses to Herbivory. J. Chem. Ecol. 2020, 46, 891–905. [Google Scholar] [CrossRef]
- Pateman, R.M.; Hill, J.K.; Roy, D.B.; Fox, R.; Thomas, C.D. Temperature-Dependent Alterations in Host Use Drive Rapid Range Expansion in a Butterfly. Science 2012, 336, 1028–1030. [Google Scholar] [CrossRef]
- Johansson, F.; Orizaola, G.; Nilsson-Örtman, V. Temperate insects with narrow seasonal activity periods can be as vulnerable to climate change as tropical insect species. Sci. Rep. 2020, 1, 1–8. [Google Scholar] [CrossRef]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T.; et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nat. Cell Biol. 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Konvicka, M.; Maradova, M.; Benes, J.; Fric, Z.; Kepka, P. Uphill shifts in distribution of butterflies in the Czech Republic: Effects of changing climate detected on a regional scale. Glob. Ecol. Biogeogr. 2003, 12, 403–410. [Google Scholar] [CrossRef] [Green Version]
- Özgökçe, M.S.; Bayındır, A.; Karaca, I.; Çobanoğlu, S.; Kumral, N.A. Temperature-dependent development of the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) on tomato plant Lycopersicon esculentum Mill. (Solanaceae). Turk. J. Èntomol. 2016, 40, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Fand, B.B.; Sul, N.T.; Bal, S.K.; Minhas, P.S. Temperature Impacts the Development and Survival of Common Cutworm (Spodoptera litura): Simulation and Visualization of Potential Population Growth in India under Warmer Temperatures through Life Cycle Modelling and Spatial Mapping. PLoS ONE 2015, 10, e0124682. [Google Scholar] [CrossRef] [Green Version]
- Kriticos, D.J.; Ota, N.; Hutchison, W.D.; Beddow, J.; Walsh, T.; Tay, W.T.; Borchert, D.M.; Paula-Moraes, S.V.; Czepak, C.; Zalucki, M. Correction: The Potential Distribution of Invading Helicoverpa armigera in North America: Is It Just a Matter of Time? PLoS ONE 2015, 10, e0119618. [Google Scholar] [CrossRef]
- Pandey, Y.; Pun, A.; Upadhyuy, K. Participatory Varietal Evaluation of Rainy Season Tomato under Plastic House Condition. Nepal Agric. Res. J. 2006, 7, 11–15. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation Site | Insect Species | Time Series Regression Equation | Slope (Mean ± SE) | Intercept | r2 Value | F-Statistics | p-Value |
|---|---|---|---|---|---|---|---|
| Gadi (1389) | H. armigera + S. litura + T. absoluta | Trap catch number = −357.40 + 19.331 × Temperature | 19.331 ± 7.274 | −357.40 ± 167.10 | 0.414 | F (1,10) = 7.06 | 0.024 |
| Jhilmile (605 m) | H. armigera + S. litura + T. absoluta | Trap catch number = 190.90 − 5.105 × Temperature | −5.105 ± 6.453 | 190.90 ± 176.00 | 0.073 | F (1,8) = 0.63 | 0.452 |
| Naubasta (189 m) | H. armigera + S. litura + T. absoluta | Trap catch number = −63.88 + 3.621 × Temperature | 3.621 ± 1.730 | −63.88 ± 49.13 | 0.354 | F (1,8) = 4.38 | 0.070 |
| Gadi (1389) | H. armigera | Trap catch number = −9.36 + 0.982 × Temperature | 0.982 ± 0.818 | −9.36 ± 18.80 | 0.126 | F (1,10) = 1.44 | 0.258 |
| Jhilmile (605 m) | H. armigera | Trap catch number = 27.58 − 0.516 × Temperature | −0.516 ± 0.684 | 27.58 ± 18.48 | 0.054 | F (1,10) = 0.57 | 0.469 |
| Naubasta (189 m) | H. armigera | Trap catch number = 30.83 − 0.668 × Temperature | −0.668 ± 0.432 | 30.83 ± 12.33 | 0.193 | F (1,10) = 2.39 | 0.153 |
| Gadi (1389) | S. litura | Trap catch number = −167.86 + 8.681 × Temperature | 8.681 ± 3.063 | −167.86 ± 70.38 | 0.445 | F (1,10) = 8.03 | 0.0018 |
| Jhilmile (605 m) | S. litura | Trap catch number = −31.45 + 1.987 × Temperature | 1.987 ± 3.647 | −31.45 ± 98.86 | 0.032 | F (1,10) = 0.30 | 0.599 |
| Naubasta (189 m) | S. litura | Trap catch number = −45.45 + 2.262 × Temperature | 2.262 ± 1.082 | −45.45 ± 30.73 | 0.353 | F (1,8) = 4.37 | 0.070 |
| Gadi (1389) | T. absoluta | Trap catch number = −180.20 + 9.669 × Temperature | 9.669 ± 5.520 | −180.20 ± 126.80 | 0.235 | F (1,9) = 3.07 | 0.110 |
| Jhilmile (605 m) | T. absoluta | Trap catch number = 98.44 − 3.040 × Temperature | −3.040 ± 2.751 | 98.44 ± 75.01 | 0.132 | F (1,10) = 0.30 | 0.599 |
| Naubasta (189 m) | T. absoluta | Trap catch number = −20.10 + 1.004 × Temperature | 1.004 ± 0.533 | −20.10 ± 15.12 | 0.308 | F (1,8) = 3.56 | 0.096 |
| Gadi (1389 m) | |
|---|---|
| Equation | Arcsine(Damage) = − 120.323 + 6.005 × Temperature |
| Slope | 6.005 ± 0.769 (mean ± SE); F = 60.91, df = 1, 293, p < 0.001 |
| Intercept | −120.323 ± 17.532 (mean ± SE) |
| Jhilmile (605 m) | |
| Equation | Arcsine(Damage) = 47.173 − 1.330 × Temperature |
| Slope | −1.330 ± 0.675 (mean ± SE); F = 3.88, df = 1, 293, p = 0.050 |
| Intercept | 47.173 ± 18.399 (mean ± SE) |
| Naubasta (189 m) | |
| Equation | Arcsine(Damage) = −1.956 + 0.457 × Temperature |
| Slope | 0.457 ± 0.489 (mean ± SE); F = 0.87, df = 1, 293, p = 0.351 |
| Intercept | −1.956 ± 13.148 (mean ± SE) |
| r2 = 0.251 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paudel, S.; Kandel, P.; Bhatta, D.; Pandit, V.; Felton, G.W.; Rajotte, E.G. Insect Herbivore Populations and Plant Damage Increase at Higher Elevations. Insects 2021, 12, 1129. https://doi.org/10.3390/insects12121129
Paudel S, Kandel P, Bhatta D, Pandit V, Felton GW, Rajotte EG. Insect Herbivore Populations and Plant Damage Increase at Higher Elevations. Insects. 2021; 12(12):1129. https://doi.org/10.3390/insects12121129
Chicago/Turabian StylePaudel, Sulav, Pragya Kandel, Dependra Bhatta, Vinod Pandit, Gary W. Felton, and Edwin G. Rajotte. 2021. "Insect Herbivore Populations and Plant Damage Increase at Higher Elevations" Insects 12, no. 12: 1129. https://doi.org/10.3390/insects12121129
APA StylePaudel, S., Kandel, P., Bhatta, D., Pandit, V., Felton, G. W., & Rajotte, E. G. (2021). Insect Herbivore Populations and Plant Damage Increase at Higher Elevations. Insects, 12(12), 1129. https://doi.org/10.3390/insects12121129