
Evaluation of Eleven Plant Species as Potential Banker Plants to Support Predatory Orius sauteri in Tea Plant Systems

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants
2.2. Insect Colonies and Food Sources
2.3. Development of O. sauteri Nymph (Eggs to Adult Stage) in Selected Plant Species
2.4. Evaluation on Reproductive Capacity of O. sauteri Adults on Different Host Plants
2.5. O. sauteri Growth in Tea Plants Intercropped with 11 Tested Plants
2.6. Data Analysis and Calculations
3. Results
3.1. Development of Orius sauteri Nymph (1st Instar to Adult)
3.2. Reproductive Capacity of Orius sauteri Adults
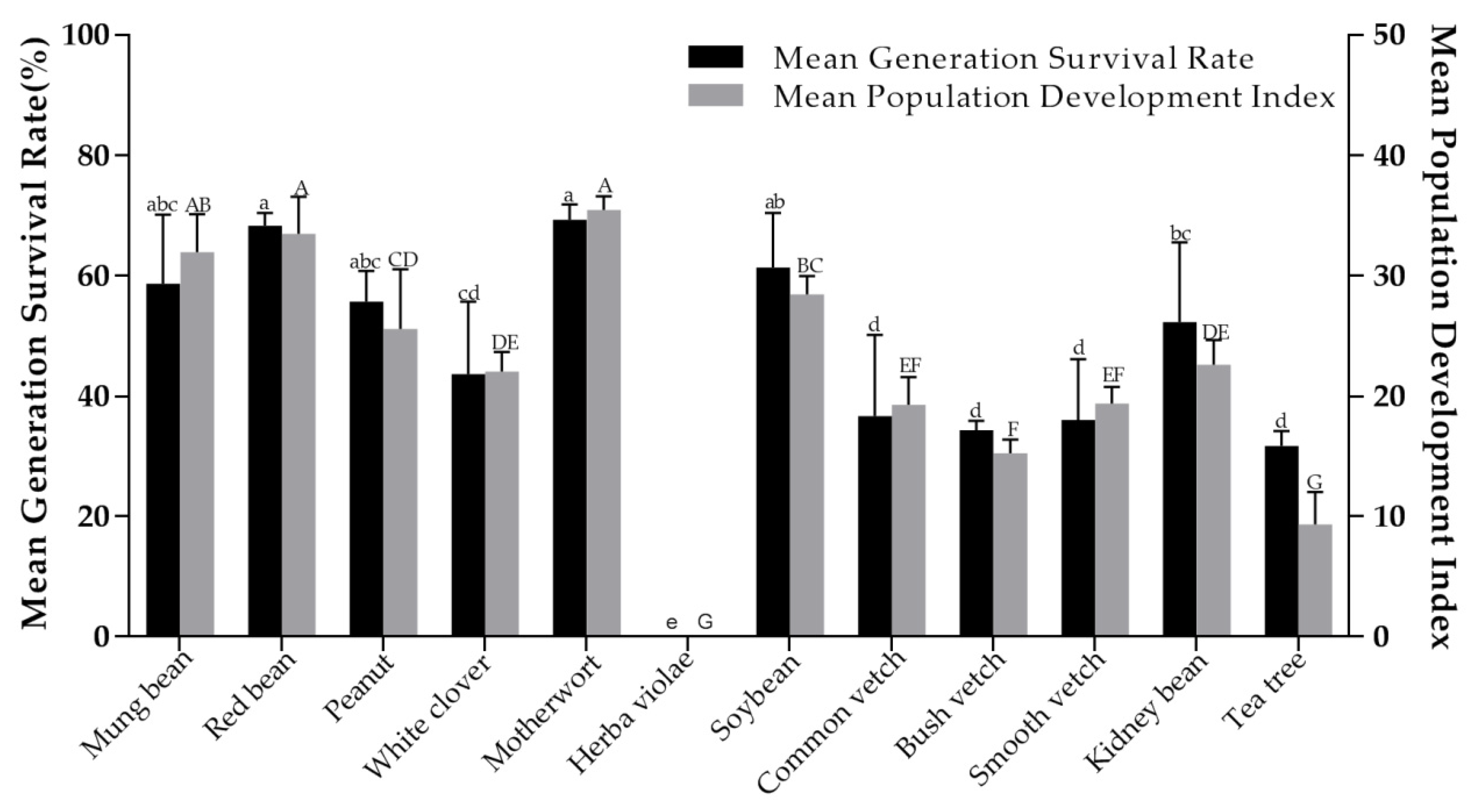
3.3. Population Life Parameters of Orius sauteri on Different Host Plants
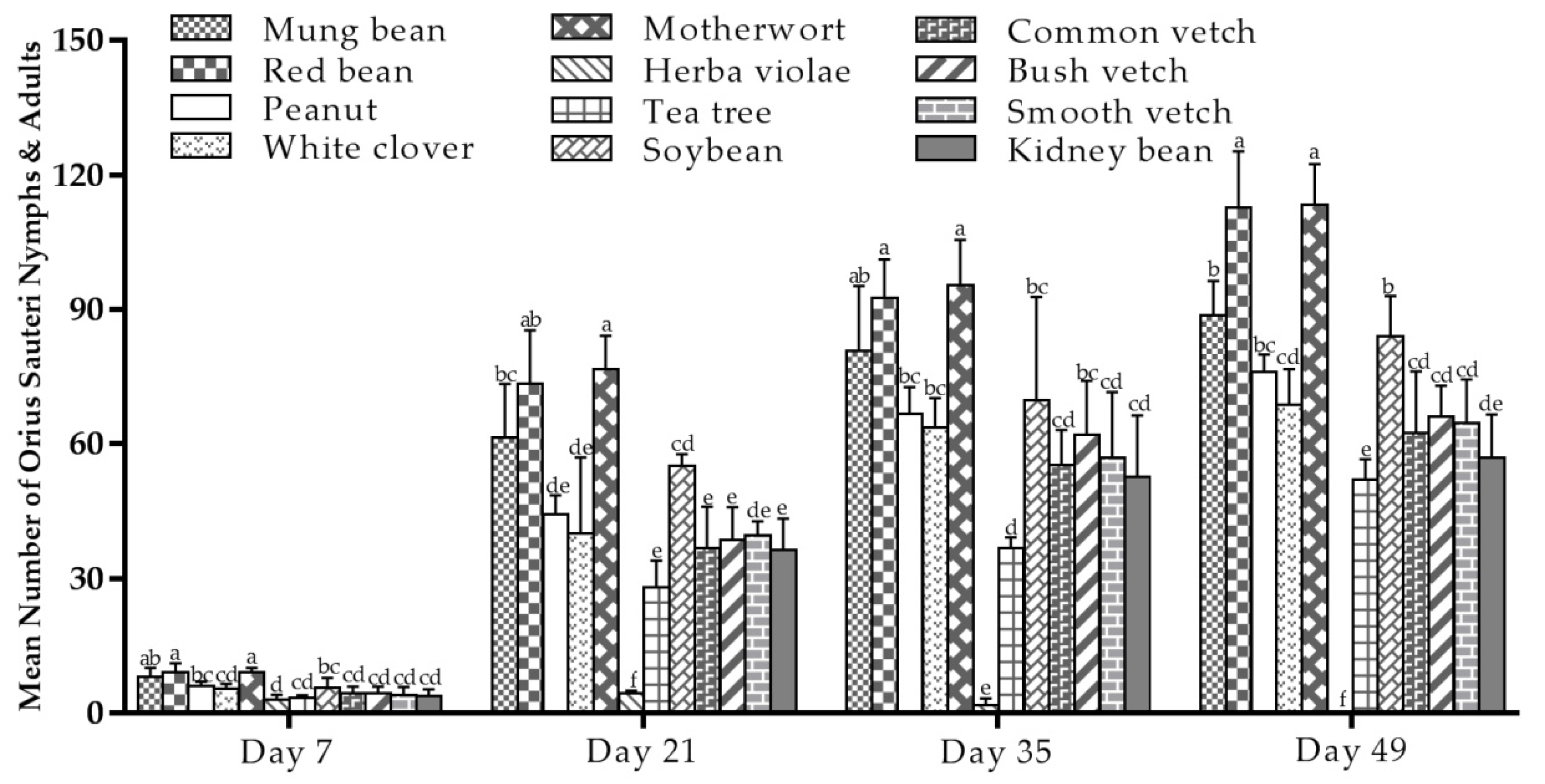
3.4. O. sauteri Growth in Tea Plants Intercropped with 11 Tested Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, H.R.; Hazarika, P. Biotechnological Approaches for Tea Improvement in Biotechnological Progress and Beverage Consumption; Academic Press: Cambridge, MA, USA, 2020; pp. 111–148. [Google Scholar]
- Zhou, Z.Y.; Hu, B.J.; Xu, L.; Hu, F.; Li, C.G.; Gao, T.C.; Su, W.H. Selection of high-efficient safe pesticides for controlling tea lesser leafhopper (Empoasca vitis). Agric. Sci. Technol. 2016, 17, 1651–1653. [Google Scholar]
- Settele, J.; Biesmeijer, J.; Bommarco, R. Switch to ecological engineering would aid independence. Nat. Cell Biol. 2008, 456, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurr, G.M.; Wratten, S.D.; Snyder, W.E.; Read, D.M.Y. Biodiversity and Insect Pests: Key Issues for Sustainable Management; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 214–229. [Google Scholar]
- Chailleux, A.; Mohl, E.K.; Alves, M.T.; Messelink, G.J.; Desneux, N. Natural enemy-mediated indirect interactions among prey species: Potential for enhancing biocontrol services in agroecosystems. Pest Manag. Sci. 2014, 70, 1769–1779. [Google Scholar] [CrossRef]
- Frank, S.D. Biological control of arthropod pests using banker plant systems: Past progress and future directions. Biol. Control. 2010, 52, 8–16. [Google Scholar] [CrossRef]
- Chen, X.X.; Liu, Y.Q.; Ren, S.X.; Zhang, F.; Zhang, W.Q.; Ge, F. Plant-mediated support system for natural enemies of insect pests. Chin. J. Appl. Entomol. 2014, 51, 1–12. [Google Scholar]
- Jaworski, C.C.; Chailleux, A.; Bearez, P.; Desneux, N. Predator-mediated apparent competition between pests fails to prevent yield loss despite actual pest population decrease. J. Pest Sci. 2015, 88, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Stacey, D.L. ’Banker’ Plant Production of Encarsia formosa Gahan and its Use in the Control of Glasshouse Whitefly on Tomatoes. Plant Pathol. 1977, 26, 63–66. [Google Scholar] [CrossRef]
- Collier, T.; Steenwyk, R.V. A critical evaluation of augmentative biological control. Biol. Control. 2004, 31, 245–256. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, K.; Jiang, Y.; Guo, Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nat. Cell Biol. 2012, 487, 362–365. [Google Scholar] [CrossRef]
- Waite, M.O.; Scott-Dupree, C.D.; Brownbridge, M.; Buitenhuis, R.; Murphy, G. Evaluation of seven plant species/cultivars for their suitability as banker plants for Orius insidiosus (Say). BioControl 2013, 59, 79–87. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, X.; Tan, X. Using Calendula officinalis as a floral resource to enhance aphid and thrips suppression by the flower bug Orius sauteri (Hemiptera: Anthocoridae). Pest Manag. Sci. 2017, 73, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wratten, S.D.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: Benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- Wäckers, F.L.; Van Rijn, P.C.J. Pick and Mix: Selecting Flowering Plants to Meet the Requirements of Target Biological Control Insects. In Biodiversity and Insect Pests; Wiley: Hoboken, NJ, USA, 2012; pp. 139–165. [Google Scholar]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Èntomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Li, H.L.; Li, P.; Zhang, H.; Wang, D.F.; Li, L.D.; Zeng, M.S.; Wu, G.Y.; Wang, Q.S. Predation of Orius sauterion pest insects of pea pushes. Acta Teas Sin. 2019, 60, 38–40. [Google Scholar]
- Tan, X.L.; Wang, S.; Liu, T.X. Acceptance and suitability of four plant substrates for rearing Orius sauteri (Hemip.tera: An-thocoridae). Biocontrol Sci. Techn. 2014, 24, 291–302. [Google Scholar] [CrossRef]
- Nagai, K.; Yano, E. Predation by Orius sauteri (Poppius) (Heteroptera: Anthocoridae) on Thrips palmi Karny (Thysanoptera: Thripidae). Functional response and selective predation. Appl. Èntomol. Zool. 2000, 35, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Gao, X.G.; Wu, Y.Q. Thrips control on the greenhouse eggplant by releasing Orius sauteri (Heteroptera: Anthocoridae). Chin. J. Biol. Control 2013, 29, 459–462. [Google Scholar]
- Kiman., Z.B.; Yearga., K.V. Development and reproduction of the predator Orius insidiosus (Hemiptera: Anthocoridae) reared on diets of selected plant material and arthropod prey. Ann Entomol Soc Am. 1985, 4, 464–467. [Google Scholar] [CrossRef]
- Yoshitaka, N.; Yoshimi, H. Effects of Prey Availability on Longevity, Prey Consumption, and Egg Production of the Insect Predators Orius sauteri and O. tantillus (Hemiptera: Anthocoridae). Ann. Entomol. Soc. Am. 1999, 92, 537–541. [Google Scholar]
- Slater, J.A. Order hemiptera: True bugs, cicadas, hoppers, psyllids, whiteflies, aphids, and scale insects. In Borror and Delong’s Introduction to the Study of Insects, 7th ed.; Triplehorn, C.A., Johnson, N.F., Eds.; Thomson Brooks/Cole: Belmont, CA, USA, 2005; pp. 268–332. [Google Scholar]
- Fan, J.S.; Zhang, L.X.; Wang, G.Q.; Ma, X.; Han, X. Life tables for experimental populations of Loxostege sticticalis (Lepidoptera: Crambidae) on five host plants. Plant Prot. 2016, 42, 104–109. [Google Scholar]
- Busvine, J.R. Revised method for spider mites and their eggs (e.g., Tetranychus sp. and Panonychus ulmi Koch). FAO method No. 10a. In Recommended Methods for Measurement of Pest Resistance to Pesticides; FAO Plant Production and Protection Paper; Food and Agriculture Organization of the United Nations: Rome, Italy, 1980; Volume 21, pp. 49–53. [Google Scholar]
- Shih, C.-I.T.; Poe, S.L.; Cromroy, H.L. Biology, Life Table, and Intrinsic Rate of Increase of Tetranychus urticae1. Ann. Èntomol. Soc. Am. 1976, 69, 362–364. [Google Scholar] [CrossRef]
- Ding, Y.Q. Population Mathematical Principles and Application of Insect Ecology; Science Press: Beijing, China, 1980. [Google Scholar]
- Gao, S.K.; Yang, Z.Q. Application of life table in pest biological control. Chin. J. Biol. Control 2015, 31, 256–263. [Google Scholar]
- Sun, R.Y. The Principle of Animal Ecology; Normal University Press: Beijing, China, 2001. [Google Scholar]
- Wong, S.K.; Frank, S.D. Pollen increases fitness and abundance of Orius insidiosus Say (Heteroptera: Anthocoridae) on banker plants. Biol. Control 2013, 64, 45–50. [Google Scholar] [CrossRef]
- Wu, H.; Zhan, Y.; Chen, H.F.; Li, J.; Jian, H.; Zhang, W.G.; Feng, Y.L.; Yang, S.L. Rapid identification of chemical constituents in violayedoensis by UHPLC-ESI-Q-TOF-MS/MS. China J. Exp. Tradit. Med. Formulae 2016, 22, 70–75. [Google Scholar]
- Oshima, N.; Narukawa, Y.; Takeda, T.; Kiuchi, F. Collagenase inhibitors from Viola yedoensis. J. Nat. Med. 2012, 67, 240–245. [Google Scholar] [CrossRef]
- Groenteman, R.; Guershon, M.; Coll, M. Effects of leaf nitrogen content on oviposition site selection, offspring performance, and intraspecific interactions in an omnivorous bug. Ecol. Èntomol. 2006, 31, 155–161. [Google Scholar] [CrossRef]
- Liman, A.-S.; Dalin, P.; Björkman, C. Enhanced leaf nitrogen status stabilizes omnivore population density. Oecologia 2017, 183, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.K.; Frank, S.D. Influence of banker plants and spiders on biological control by Orius insidiosus (Heteroptera: Anthocoridae). Biol. Control 2012, 63, 181–187. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. | Common Name | Latin Name/Family | Source * |
|---|---|---|---|
| 1 | Motherwort | Leonurus artemisia (Lour.) S. Y. Hu/Labiatae | Blue Sky Seed Industry |
| 2 | White clover | Trifolium repens./Leguminosae | Blue Sky Seed Industry |
| 3 | Bush vetch | Vicia villosa Roth./Leguminosae | Blue Sky Seed Industry |
| 4 | Common vetch | Vicia sativa L./Leguminosae | Blue Sky Seed Industry |
| 5 | Smooth vetch | Vicia glabrescens Koch./Leguminosae | Blue Sky Seed Industry |
| 6 | Herba violae | Viola philippica./Violaceae | Blue Sky Seed Industry |
| 7 | Soybean | Glycine javanica L./Leguminosae | Guizhou Dasheng Seed Industry Co., Ltd. |
| 8 | Mung bean | Vigna radiata (Linn.) Wilczek/Leguminosae | Guizhou Dasheng Seed Industry Co., Ltd. |
| 9 | Red bean | Vigna angularis (Willd.) Ohwiet Ohashi/Leguminosae | Guizhou Dasheng Seed Industry Co., Ltd. |
| 10 | Peanut | Arachis hypogaea Linn./Leguminosae | Guizhou Dasheng Seed Industry Co., Ltd. |
| 11 | Kidney bean | Phaseolus vulgaris Linn./Leguminosae | Dezhou De Vegetable Seed Industry Co., Ltd. |
| 12 | Tea plant | Camellia sinensis (L.) O. Ktze/Theaceae | Guizhou Tea plantation |
| Test Plants | Hatching Rate (%) | Nymph Development Time (d) | Emergence Rate (%) |
|---|---|---|---|
| Mung bean | 87.62 ± 3.06 a | 13.00 ± 0.58 a | 71.26 ± 1.90 a |
| Red bean | 87.62 ± 3.06 a | 12.33 ± 0.88 a | 76.67 ± 3.16 a |
| Peanut | 89.12 ± 1.82 a | 11.00 ± 1.45 a | 54.55 ± 3.54 bc |
| White clover | 93.33 ± 0.66 a | 14.00 ± 0.58 a | 54.69 ± 6.52 bc |
| Motherwort | 91.37 ± 2.39 a | 12.33 ± 0.88 a | 74.93 ± 3.35 a |
| Herba violae | 90.30 ± 2.36 a | 1.50 ± 0.50 b | 0.00 ± 0.00 f |
| Soybean | 86.44 ± 2.23 a | 12.33 ± 0.88 a | 67.24 ± 3.82 ab |
| Common vetch | 86.40 ± 2.36 a | 12.33 ± 1.45 a | 42.35 ± 4.27 cd |
| Bush vetch | 85.72 ± 2.15 a | 12.00 ± 1.15 a | 45.40 ± 6.57 cd |
| Smooth vetch | 86.41 ± 2.00 a | 12.33 ± 0.33 a | 51.94 ± 3.07 c |
| Kidney bean | 88.34 ± 2.64 a | 12.00 ± 1.73 a | 56.56 ± 7.70 bc |
| * Tea plant | 86.50 ± 2.13 a | 13.80 ± 0.42 a | 32.51 ± 3.71 d |
| Test Plants | Fecundity (Eggs per Female) | Oviposition Period (Day) |
|---|---|---|
| Mung bean | 124.75 ± 12.99 abc | 15.25 ± 1.31 abcd |
| Red bean | 148.75 ± 8.06 a | 20.50 ± 1.55 a |
| Peanut | 121.00 ± 9.46 abc | 16.75 ± 1.38 abcd |
| White clover | 132.25 ± 10.19 ab | 18.00 ± 1.22 abc |
| Motherwort | 148.25 ± 11.02 a | 19.25 ± 1.93 ab |
| Herba violae | 94.60 ± 5.40 bc | 11.60 ± 0.75 d |
| Soybean | 122.25 ± 5.82 abc | 16.00 ± 1.08 abcd |
| Common vetch | 103.50 ± 4.63 bc | 14.75 ± 0.85 bcd |
| Bush vetch | 119.75 ± 8.41 abc | 14.75 ± 1.03 bcd |
| Smooth vetch | 100.00 ± 6.72 bc | 13.25 ± 0.85 cd |
| Kidney bean | 117.75 ± 6.80 abc | 14.75 ± 0.85 bcd |
| * Tea plants | 90.20 ± 4.85 c | 12.00 ± 0.71 d |
| Test Plants | Net Reproductive Rate (R0) | Mean Generation Time (T)/d | Intrinsic Rate of Increase (rm) | Finite Increase Rate (λ) | Population Doubling Time (t)/d |
|---|---|---|---|---|---|
| Mung bean | 65.01 bc | 26.87 a | 0.15 cd | 1.17 cd | 4.47 bc |
| Red bean | 76.75 a | 26.26 ab | 0.16 abc | 1.18 abc | 4.20 c |
| Peanut | 44.58 de | 22.03 f | 0.17 ab | 1.19 ab | 4.02 d |
| White clover | 54.44 cd | 25.90 bc | 0.15 cd | 1.17 cd | 4.49 bc |
| Motherwort | 72.89 ab | 24.08 e | 0.18 a | 1.19 a | 3.90 d |
| Herba violae | / | / | / | / | / |
| Soybean | 64.96 bc | 25.41 bc | 0.16 bc | 1.18 bc | 4.23 c |
| Common vetch | 40.38 e | 23.70 e | 0.16 cd | 1.17 cd | 4.45 de |
| Bush vetch | 39.73 ef | 25.15 cd | 0.15 de | 1.16 de | 4.74 b |
| Smooth vetch | 38.46 ef | 25.15 cd | 0.14 de | 1.16 de | 4.79 b |
| Kidney bean | 35.86d ef | 24.44 de | 0.15 de | 1.16 de | 4.75b |
| * Tea plants | 29.35 f | 25.21 cd | 0.13 e | 1.14 e | 5.19 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Ji, D.; Zhang, Q.; Jin, L. Evaluation of Eleven Plant Species as Potential Banker Plants to Support Predatory Orius sauteri in Tea Plant Systems. Insects 2021, 12, 162. https://doi.org/10.3390/insects12020162
Zhang R, Ji D, Zhang Q, Jin L. Evaluation of Eleven Plant Species as Potential Banker Plants to Support Predatory Orius sauteri in Tea Plant Systems. Insects. 2021; 12(2):162. https://doi.org/10.3390/insects12020162
Chicago/Turabian StyleZhang, Ruifang, Dezhong Ji, Qiuqiu Zhang, and Linhong Jin. 2021. "Evaluation of Eleven Plant Species as Potential Banker Plants to Support Predatory Orius sauteri in Tea Plant Systems" Insects 12, no. 2: 162. https://doi.org/10.3390/insects12020162
APA StyleZhang, R., Ji, D., Zhang, Q., & Jin, L. (2021). Evaluation of Eleven Plant Species as Potential Banker Plants to Support Predatory Orius sauteri in Tea Plant Systems. Insects, 12(2), 162. https://doi.org/10.3390/insects12020162