Temperature Affects Biological Control Efficacy: A Microcosm Study of Trichogramma achaeae


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Laboratory Rearing
2.3. Microcosm Test of Biological Control Efficacy
2.4. Data Analysis
3. Results
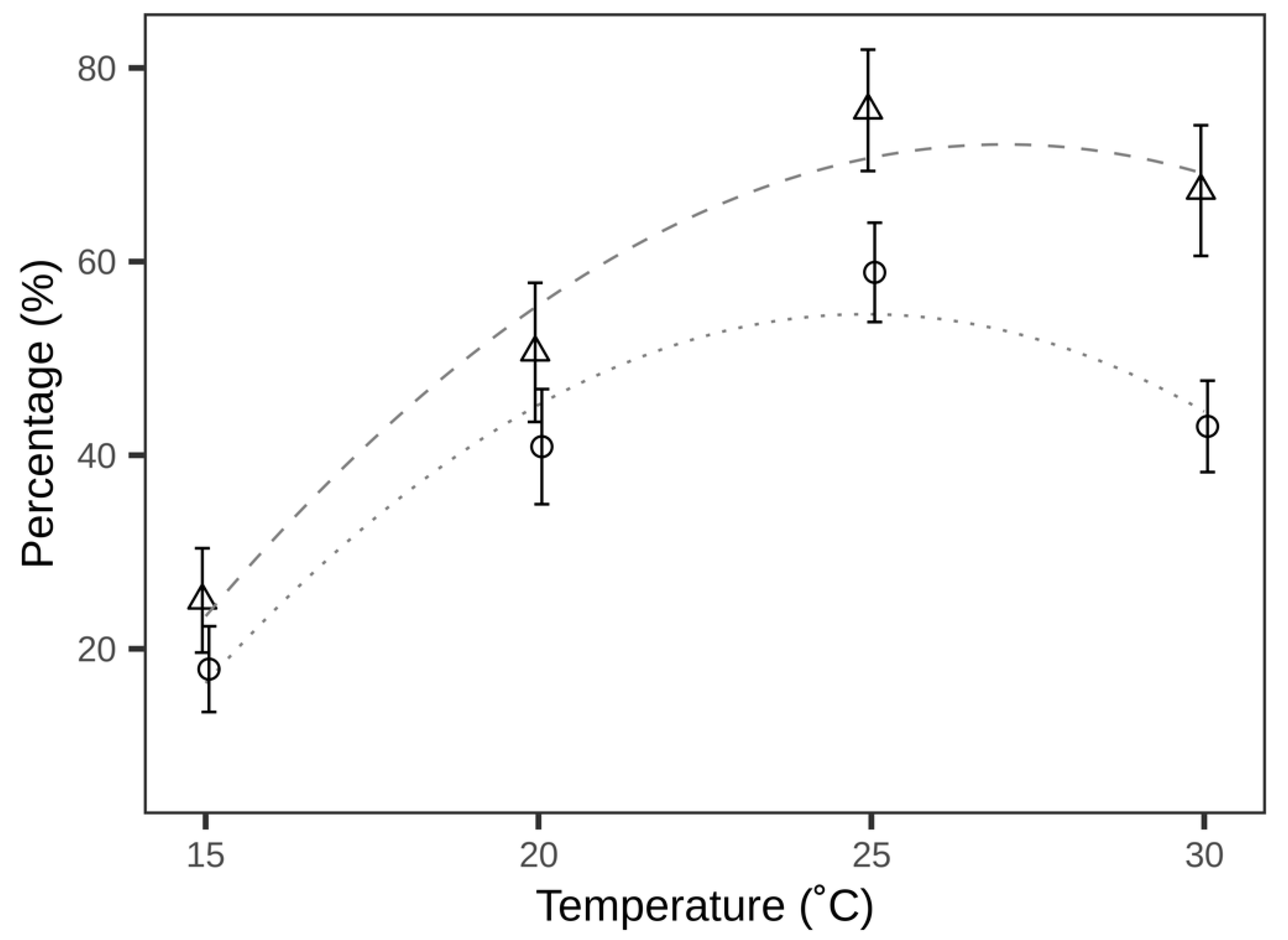
3.1. Realised Microcosm Parasitisation at Different Temperatures
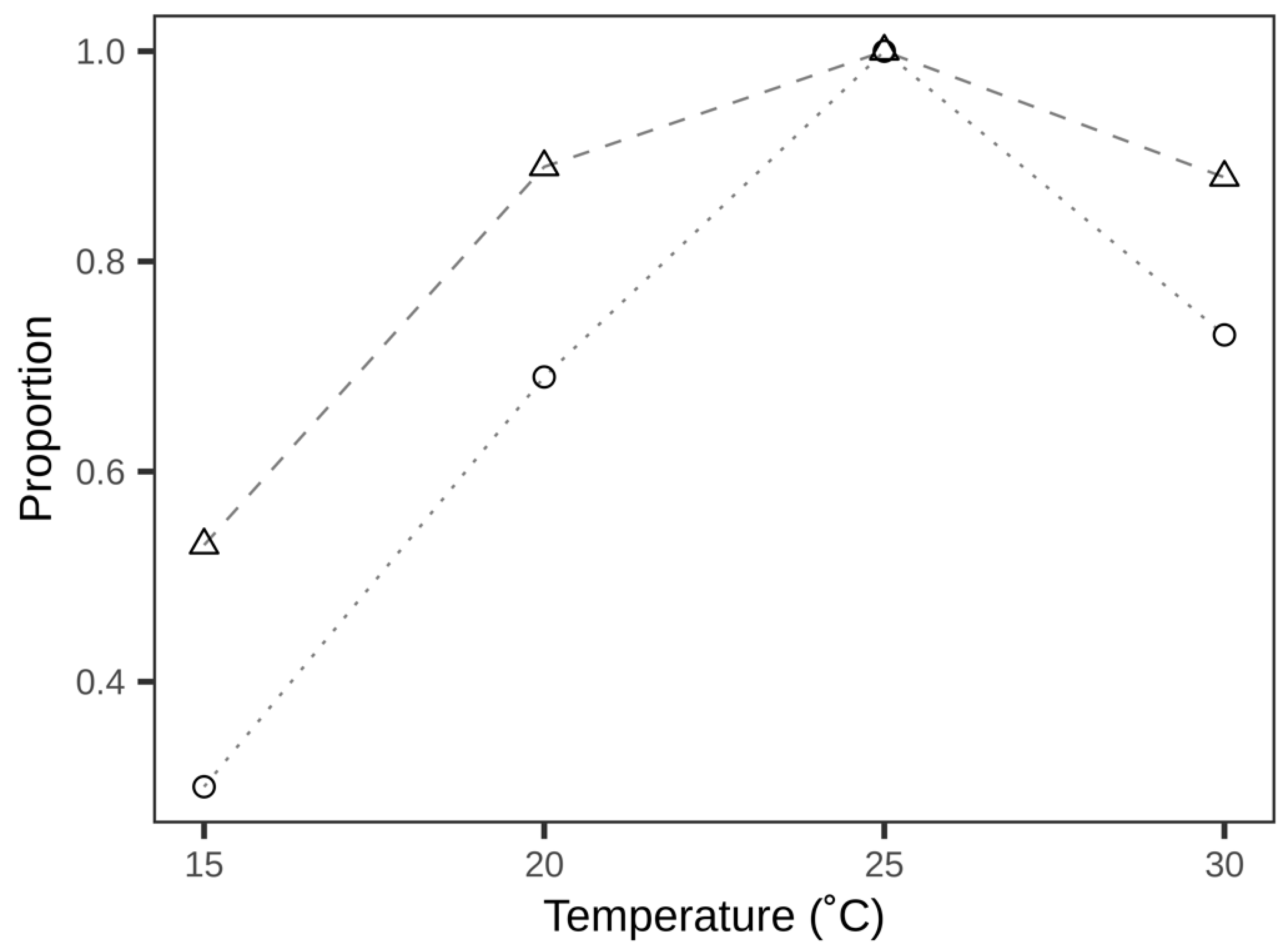
3.2. Distribution of Parasitised Eggs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mackauer, M. Genetic problems in the production of biological control agents. Annu. Rev. Entomol. 1976, 21, 369–385. [Google Scholar] [CrossRef]
- Cônsoli, F.L.; Parra, J.R.P. Biology of Trichogramma galloi and T. pretiosum (Hymenoptera: Trichogrammatidae) reared in vitro and in vivo. Ann. Entomol. Soc. Am. 1996, 89, 828–834. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Hale, A.; Klapwijk, J.N.; van Schelt, J.; Steinberg, S. Guidelines for quality control of commercially produced natural enemies. In Quality Control and Production of Biological Control Agents: Theory and Testing Procedures; van Lenteren, J.C., Ed.; CABI Publishing: Wallingford, UK, 2003; pp. 265–303. [Google Scholar]
- Gariepy, V.; Boivin, G.; Brodeur, J. Why two species of parasitoids showed promise in the laboratory but failed to control the soybean aphid under field conditions. Biol. Control 2014, 80, 1–7. [Google Scholar] [CrossRef]
- van Lenteren, J.C. (Ed.) Need for quality control of mass-produced biological control agents. In Quality Control and Production of Biological Control Agents: Theory and Testing Procedures; CABI Publishing: Wallingford, UK, 2003; pp. 1–18. [Google Scholar]
- Bigler, F. Quality assessment and control in entomophagous insects used for biological control. J. Appl. Entomol. 1989, 108, 390–400. [Google Scholar] [CrossRef]
- Lundgren, J.G.; Heimpel, G.E. Quality assessment of three species of commercially produced Trichogramma and the first report of thelytoky in commercially produced Trichogramma. Biol. Control 2003, 26, 68–73. [Google Scholar] [CrossRef]
- Cascone, P.; Carpenito, S.; Slotsbo, S.; Iodice, L.; Sørensen, J.G.; Holmstrup, M.; Guerrieri, E. Improving the efficiency of Trichogramma achaeae to control Tuta absoluta. BioControl 2015, 60, 761–771. [Google Scholar] [CrossRef]
- Höller, C.; Haardt, H. Low field performance of an aphid parasitoid, Aphelinus abdominalis, efficient in the laboratory (Hym., Aphelinidae). Entomophaga 1993, 38, 115–124. [Google Scholar] [CrossRef]
- Dutton, A.; Bigler, F. Flight activity assessment of the egg parasitoid Trichogramma brassicae (Hym.: Trichogrammatidae) in laboratory and field conditions. Entomophaga 1995, 40, 223–233. [Google Scholar] [CrossRef]
- Brown, R.L.; Stanbury, M.; El-Sayed, A.M.; Laban, J.; Butler, R.; Suckling, D.M. Locomotion activity meter for quality assessment of mass-reared sterile male moths (Lepidoptera). Fla. Entomol. 2016, 99, 131–137. [Google Scholar] [CrossRef]
- Thomson, L.J.; Hoffmann, A.A. Laboratory fecundity as predictor of field success in Trichogramma carverae (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2002, 95, 912–917. [Google Scholar] [CrossRef]
- Dutton, A.; Cerutti, F.; Bigler, F. Quality and environmental factors affecting Trichogramma brassicae efficiency under field conditions. Entomol. Exp. Appl. 1996, 81, 71–79. [Google Scholar] [CrossRef]
- Cônsoli, F.L.; Parra, J.R.P. Effects of constant and alternating temperatures on Trichogramma galloi Zucchi (Hym., Trichogrammatidae) biology. I. Development and thermal requirements. J. Appl. Entomol. 1995, 119, 415–418. [Google Scholar] [CrossRef]
- Cônsoli, F.L.; Parra, J.R.P. Effects of constant and alternating temperatures on Trichogramma galloi Zucchi (Hym., Trichogrammatidae) biology. II. Parasitism capacity and longevity. J. Appl. Entomol. 1995, 119, 667–670. [Google Scholar] [CrossRef]
- Castellanos, N.L.; Bueno, A.F.; Haddi, K.; Silveira, E.C.; Rodrigues, H.S.; Hirose, E.; Smagghe, G.; Oliveira, E.E. The fitness and economic benefits of rearing the parasitoid Telenomus podisi under fluctuating temperature regime. Neotrop. Entomol. 2019, 48, 934–948. [Google Scholar] [CrossRef] [PubMed]
- McCalla, K.A.; Keçeci, M.; Milosavljević, I.; Ratkowsky, D.A.; Hoddle, M.S. The Influence of temperature variation on life history parameters and thermal performance curves of Tamarixia radiata (Hymenoptera: Eulophidae), a parasitoid of the Asian citrus psyllid (Hemiptera: Liviidae). J. Econ. Entomol. 2019, 112, 1560–1574. [Google Scholar] [CrossRef]
- Bellefeuille, Y.; Fournier, M.; Lucas, E. Evaluation of two potential biological control agents against the foxglove aphid at low temperatures. J. Insect Sci. 2019, 19. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, J.G.; Addison, M.F.; Terblanche, J.S. Mass-rearing of insects for pest management: Challenges, synergies and advances from evolutionary physiology. Crop Prot. 2012, 38, 87–94. [Google Scholar] [CrossRef]
- Smith, S.M. Biological control with Trichogramma: Advances, successes, and potential of their use. Annu. Rev. Entomol. 1996, 41, 375–406. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.-S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status and perspectives. Annu. Rev. Entomol. 2021, 66. [Google Scholar] [CrossRef]
- Cabello, T.; Gámez, M.; Varga, Z.; Garay, J.; Carreño, R.; Gallego, J.R.; Fernández, F.J. Selection of Trichogramma spp. (Hym.: Trichogrammatidae) for the biological control of Tuta absoluta (Lep.: Gelechiidae) in greenhouses by an entomo-ecological simulation model. IOBC WPRS Bull. 2012, 80, 171–176. [Google Scholar]
- Oliveira, L.; Durão, A.C.; Fontes, J.; Roja, I.S.; Tavares, J. Potential of Trichogramma achaeae (Hymenoptera: Trichogrammatidae) in biological control of Tuta absoluta (Lepidoptera: Gelechiidae) in Azorean greenhouse tomato crops. J. Econ. Entomol. 2017, 110, 2010–2015. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Sørensen, J.G.; Enkegaard, A. Effects of temperature on development, reproduction and size of Trichogramma achaeae: Implications for biological control. bioRxiv 2020. [Google Scholar] [CrossRef]
- Prasad, R.P.; Roitberg, B.D.; Henderson, D. The effect of rearing temperature on flight initiation of Trichogramma sibericum Sorkina at ambient temperatures. Biol. Control 1999, 16, 291–298. [Google Scholar] [CrossRef]
- Suverkropp, B.P.; Bigler, F.; van Lenteren, J.C. Temperature influences walking speed and walking activity of Trichogramma brassicae (Hym., Trichogrammatidae). J. Appl. Entomol. 2001, 125, 303–307. [Google Scholar] [CrossRef]
- Singhamuni, S.A.A.; Hemachandra, K.S.; Sirisena, U.G.A.I. Potential for mass rearing of the egg parasitoids, Trichogramma chilonis and Trichogramma achaeae (Hymenoptera: Trichogrammatidae) on Corcyra cephalonica eggs. Trop. Agric. Res. 2016, 27, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Good, D.S. Evolution of behaviours in Drosophila melanogaster in high temperatures: Genetic and environmental effects. J. Insect Physiol. 1993, 39, 537–544. [Google Scholar] [CrossRef]
- Mills, N.J.; Kuhlmann, U. The relationship between egg load and fecundity among Trichogramma parasitoids. Ecol. Entomol. 2000, 25, 315–324. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org (accessed on 1 December 2020).
- Del Pino, M.; Gallego, J.R.; Hernández Suárez, E.; Cabello, T. Effect of temperature on life history and parasitization behavior of Trichogramma achaeae Nagaraja and Nagarkatti (Hym.: Trichogrammatidae). Insects 2020, 11, 482. [Google Scholar] [CrossRef]
- Bourchier, R.S.; Smith, S.M. Influence of environmental conditions and parasitoid quality on field performance of Trichogramma minutum. Entomol. Exp. Appl. 1996, 80, 461–468. [Google Scholar] [CrossRef]
- Bell, W.J. Searching behavior patterns in insects. Annu. Rev. Entomol. 1990, 35, 447–467. [Google Scholar] [CrossRef]
- Forsse, E.; Smith, S.M.; Bourchier, R.S. Flight initiation in the egg parasitoid Trichogramma minutum: Effects of ambient temperature, mates, food, and host eggs. Entomol. Exp. Appl. 1992, 62, 147–154. [Google Scholar] [CrossRef]
- Samietz, J.; Köhler, G. A fecundity cost of (walking) mobility in an insect. Ecol. Evol. 2012, 2, 2788–2793. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, K.; Wyckhuys, K.A.G.; Heimpel, G.E. Trade-offs between flight and fecundity in the soybean aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2009, 102, 133–138. [Google Scholar] [CrossRef]
- Milosavljević, I.; McCalla, K.A.; Ratkowsky, D.A.; Hoddle, M.S. Effects of constant and fluctuating temperatures on development rates and longevity of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae). J. Econ. Entomol. 2019, 112, 1062–1072. [Google Scholar] [CrossRef]
- Milosavljević, I.; McCalla, K.A.; Morgan, D.J.W.; Hoddle, M.S. The effects of constant and fluctuating temperatures on development of Diaphorina citri (Hemiptera: Liviidae), the Asian citrus psyllid. J. Econ. Entomol. 2020, 113, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Kogel, W.; Hoek, M.; Mollema, C. Oviposition preference of western flower thrips for cucumber leaves from different positions along the plant stem. Entomol. Exp. Appl. 1997, 82, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Suverkropp, B.P.; Dutton, A.; Bigler, F.; van Lenteren, J.C. Oviposition behaviour and egg distribution of the european corn borer, Ostrinia nubilalis, on maize, and its effect on host finding by Trichogramma egg parasitoids. Bull. Insectol. 2008, 61, 303–312. [Google Scholar]
- Salama, H.S.A.E.-R.; Ismail, I.A.-K.; Fouda, M.; Ebadah, I.; Shehata, I. Some ecological and behavioral aspects of the tomato leaf miner Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Ecol. Balk. 2015, 7, 35–44. [Google Scholar]

{kind=link}
{kind=link}
| Temperature (°C) | Number of Plants Used | Number (and %) of Plants with Parasitised Eggs | Leaves with Parasitised Eggs (%) | Parasitisation Efficiency (%) |
|---|---|---|---|---|
| 15 | 32 | 18 (56.25) | 25.00 ± 5.39 | 17.90 ± 4.43 |
| 20 | 32 | 22 (68.75) | 50.62 ± 7.18 | 40.88 ± 5.95 |
| 25 | 32 | 29 (90.62) | 75.62 ± 6.27 | 58.89 ± 5.13 |
| 30 | 30 | 26 (88.67) | 67.33 ± 6.75 | 42.98 ± 4.72 |
| Temperature (°C) | Number of Plants (Per Leaf Side) | Number of Plants with Parasitised Eggs | Leaves with Parasitised Eggs (%) | Parasitisation Efficiency (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | Upper | Lower | Upper | Lower | Upper | Lower | Top Tier | Middle Tier | Bottom Tier | |
| 15 | 16 | 10 | 8 | 42.00 ± 9.21 a | 47.50 ± 9.64 a | 32.46 ± 9.67 a | 31.03 ± 7.41 a | 51.40 ± 9.23 a | 35.45 ± 7.84 ab | 19.82 ± 6.83 b |
| 20 | 16 | 12 | 10 | 70.00 ± 7.18 a | 78.00 ± 8.67 a | 54.82 ± 5.79 a | 65.04 ± 8.01 a | 70.67 ± 8.34 a | 64.39 ± 5.46 a | 48.16 ± 7.94 a |
| 25 | 16 | 14 | 15 | 80.00 ± 6.95 a | 86.67 ± 7.22 a | 61.50 ± 5.39 a | 68.22 ± 6.54 a | 81.53 ± 3.84 a | 66.40 ± 5.05 b | 55.42 ± 6.64 b |
| 30 | 15 | 13 | 13 | 70.77 ± 8.36 a | 84.62 ± 6.47 a | 47.11 ± 6.08 a | 52.06 ± 5.63 a | 69.12 ± 4.92 a | 44.81 ± 5.72 b | 43.68 ± 4.70 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Enkegaard, A.; Sørensen, J.G. Temperature Affects Biological Control Efficacy: A Microcosm Study of Trichogramma achaeae. Insects 2021, 12, 95. https://doi.org/10.3390/insects12020095
Chen L, Enkegaard A, Sørensen JG. Temperature Affects Biological Control Efficacy: A Microcosm Study of Trichogramma achaeae. Insects. 2021; 12(2):95. https://doi.org/10.3390/insects12020095
Chicago/Turabian StyleChen, Long, Annie Enkegaard, and Jesper Givskov Sørensen. 2021. "Temperature Affects Biological Control Efficacy: A Microcosm Study of Trichogramma achaeae" Insects 12, no. 2: 95. https://doi.org/10.3390/insects12020095