
Detection of Phosphine Resistance in Field Populations of Four Key Stored-Grain Insect Pests in Pakistan

 ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Insect Populations
2.2. Phosphine Gas Generation
2.3. Bioassays
2.4. Statistical Analyses
3. Results
3.1. Resistance in R. dominica
3.2. Resistance in S. granarius
3.3. Resistance in T. castaneum
3.4. Resistance in T. granarium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arthur, F.H. Efficacy of combinations of methoprene and deltamethrin as long-term commodity protectants. Insects 2019, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Phillips, T.W.; Thoms, E.M.; DeMark, J.; Walse, S. Fumigation. In Stored Product Protection; Hagstrum, D.W., Phillips, T.W., Cuperus, G., Eds.; Kansas State University: Manhattan, KS, USA, 2012; pp. 157–177. [Google Scholar]
- Wijayaratne, L.K.W.; Rajapakse, R.H.S. Effects of spinosad on the heat tolerance and cold tolerance of Sitophilus oryzae L. (Coleoptera: Curculionidae) and Rhyzopertha dominica F. (Coleoptera: Bostrichidae). J. Stored Prod. Res. 2018, 77, 84–88. [Google Scholar] [CrossRef]
- Opit, G.; Collins, P.J.; Daglish, G.J. Resistance management. In Stored Product Protection; Hagstrum, D.W., Phillips, T.W., Cuperus, G., Eds.; Kansas State University: Manhattan, KS, USA, 2012; pp. 143–155. [Google Scholar]
- Pimentel, M.A.G.; Faroni, L.R.A.; Tótola, M.R.; Guedes, R.N.C. Phosphine resistance, respiration rate and fitness consequences in stored-product insects. Pest Manag. Sci. 2007, 63, 876–881. [Google Scholar] [CrossRef] [Green Version]
- Rossi, E.; Cosimi, S.; Loni, A. Insecticide resistance in Italian populations of Tribolium flour beetles. Bull. Insectol. 2010, 63, 251–285. [Google Scholar]
- Daglish, G.J.; Nayak, M.K. Prevalence of resistance to deltamethrin in Rhyzopertha dominica (F.) in eastern Australia. J. Stored Prod. Res. 2018, 78, 45–49. [Google Scholar] [CrossRef]
- Attia, M.A.; Wahba, T.F.; Shaarawy, N.; Moustafa, F.I.; Guedes, R.N.C.; Dewer, Y. Stored grain pest prevalence and insecticide resistance in Egyptian populations of the red flour beetle Tribolium castaneum (Herbst) and the rice weevil Sitophilus oryzae (L.). J. Stored Prod. Res. 2020, 87, 101611. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Lima, J.O.G.; Santos, J.P.; Cruz, C.D. Inheritance of deltamethrin resistance in a Brazilian strain of maize weevil (Sitophilus zeamais Mots.). Int. J. Pest Manag. 1994, 40, 103–106. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Lima, J.O.G.; Santos, J.P.; Cruz, C.D. Resistance to DDT and pyrethroids in Brazilian populations of Sitophilus zeamais Motsch. (Coleoptera: Curculionidae). J. Stored Prod. Res. 1995, 31, 145–150. [Google Scholar] [CrossRef]
- Lorini, I.; Galley, D.J. Deltamethrin resistance in Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae), a pest of stored grain in Brazil. J. Stored Prod. Res. 1999, 35, 37–45. [Google Scholar] [CrossRef]
- Fragoso, D.B.; Guedes, R.N.C.; Rezende, S.T. Glutathione S-transferase detoxification as a potential pyrethroid resistance mechanism in the maize weevil, Sitophilus zeamais. Entomol. Exp. Appl. 2003, 109, 21–29. [Google Scholar] [CrossRef]
- Ribeiro, B.M.; Guedes, R.N.C.; Oliveira, E.E.; Santos, J.P. Insecticide resistance and synergism in Brazilian populations of Sitophilus zeamais (Coleoptera: Curculionidae). J. Stored Prod. Res. 2003, 39, 21–31. [Google Scholar] [CrossRef]
- Fragoso, D.B.; Guedes, R.N.C.; Peternelli, L.A. Developmental rates and population growth of insecticide resistant and susceptible populations of Sitophilus zeamais. J. Stored Prod. Res. 2005, 41, 271–281. [Google Scholar] [CrossRef]
- Perez Mendoza, J. Survey of insecticide resistance in Mexican populations of maize weevil, Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). J. Stored Prod. Res. 1999, 35, 107–115. [Google Scholar] [CrossRef]
- Beckett, S.J.; Evans, D.E.; Morton, R. A comparison of the demographies of pesticide susceptible and resistant strains of Oryzaephilus surinamensis (L.) (Coleoptera: Silvanidae) on kibbled wheat. J. Stored Prod. Res. 1996, 32, 141–151. [Google Scholar] [CrossRef]
- Bell, C.H. Fumigation in the 21st century. Crop Prot. 2000, 19, 563–569. [Google Scholar] [CrossRef]
- Chaudhry, M.Q. Phoshine resistance: A growing threat to an ideal fumigant. Pestic. Outlook 2000, 11, 88–91. [Google Scholar] [CrossRef]
- Phillips, T.W.; Throne, J.E. Biorational approaches to managing stored-product insects. Annu. Rev. Entomol. 2010, 55, 375–397. [Google Scholar] [CrossRef]
- Kaur, R.; Nayak, M.K. Developing effective fumigation protocols to manage strongly phosphine-resistant Cryptolestes ferrugineus (Stephens) (Coleoptera: Laemophloeidae). Pest Manag. Sci. 2015, 71, 1297–1302. [Google Scholar] [CrossRef] [Green Version]
- Holloway, J.C.; Falk, M.G.; Emery, R.N.; Collins, P.J.; Nayak, M.K. Resistance to phosphine in Sitophilus oryzae in Australia: A national analysis of trends and frequencies over time and geographical spread. J. Stored Prod. Res. 2016, 69, 129–137. [Google Scholar] [CrossRef]
- Nayak, M.K.; Daglish, G.J.; Phillips, T.W. Managing resistance to chemical treatments in stored products pests. Stewart Postharvest Rev. 2015, 11, 3. [Google Scholar]
- Afful, E.; Elliott, B.; Nayak, M.K.; Phillips, T.W. Phosphine resistance in North American field populations of the lesser grain borer, Rhyzopertha dominica (Coleoptera: Bostrichidae). J. Econ. Entomol. 2018, 111, 463–469. [Google Scholar] [CrossRef]
- Champ, B.R.; Dyte, C.E. Report of the FAO Global Survey of Pesticide Susceptibility of Stored Grain Pests; Food and Agriculture Organization of the United Nations: Rome, Italy, 1976. [Google Scholar]
- Lorini, I.; Collins, P.J.; Daglish, G.J.; Nayak, M.K.; Pavic, H. Detection and characterisation of strong resistance to phosphine in Brazilian Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). Pest Manag. Sci. 2007, 63, 358–364. [Google Scholar] [CrossRef]
- Opit, G.P.; Phillips, T.W.; Aikins, M.J.; Hasan, M.M. Phosphine resistance in Tribolium castaneum and Rhyzopertha dominica from stored wheat in Oklahoma. J. Econ. Entomol. 2012, 105, 1107–1114. [Google Scholar] [CrossRef] [Green Version]
- Nayak, M.K.; Holloway, J.C.; Emery, R.N.; Pavic, H.; Bartlet, J.; Collins, P.J. Strong resistance to phosphine in the rusty grain beetle, Cryptolestes ferrugineus (Stephens) (Coleoptera: Laemophloeidae): Its characterisation, a rapid assay for diagnosis and its distribution in Australia. Pest Manag. Sci. 2013, 69, 48–53. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Collins, P.J.; Ebert, P.R. Inheritance and characterization of strong resistance to phosphine in Sitophilus oryzae (L.). PLoS ONE 2015, 10, e0124335. [Google Scholar] [CrossRef] [Green Version]
- Afful, E.; Tadesse, T.M.; Nayak, M.K.; Phillips, T.W. High-dose strategies for managing phosphine resistant populations of Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae). Pest Manag. Sci. 2020, 76, 1683–1690. [Google Scholar] [CrossRef]
- Afful, E.; Cato, A.; Nayak, M.K.; Phillips, T.W. A rapid assay for the detection of resistance to phosphine in the lesser grain borer, Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae). J. Stored Prod. Res. 2021, 91, 101776. [Google Scholar] [CrossRef]
- Chen, Z.; Schlipalius, D.; Opit, G.; Subramanyam, B.H.; Phillips, T.W. Diagnostic molecular markers for phosphine resistance in U.S. populations of Tribolium castaneum and Rhyzopertha dominica. PLoS ONE 2015, 10, e0121343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, R.; Subbarayalu, M.; Jagadeesan, R.; Daglish, G.J.; Nayak, M.K.; Naik, H.R.; Ramasamy, S.; Subramanian, C.; Ebert, P.R.; Schlipalius, D.I. Phosphine resistance in India is characterised by a dihydrolipoamide dehydrogenase variant that is otherwise unobserved in eukaryotes. Heredity 2015, 115, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S.G.; Opit, G.P.; Konemann, C.; Shakya, K.; Hosoda, E. Phosphine resistance in saw-toothed grain beetle, Oryzaephilus surinamensis in the United States. J. Stored Prod. Res. 2020, 89, 101690. [Google Scholar] [CrossRef]
- Zettler, J.L. Influence of resistance of future fumigation technology. In Proceedings of the International Conference on Controlled Atmosphere and Fumigation in Stored Products, Nicosia, Cyprus, 21–26 April 1996; Donahaye, E.J., Navarro, S., Varnava, A., Eds.; Printco Ltd.: Nicosia, Cyprus, 1997; pp. 445–454. [Google Scholar]
- Cato, A.J.; Elliott, B.; Nayak, M.K.; Phillips, T.W. Geographic variation in phosphine resistance among North American populations of the red flour beetle. J. Econ. Entomol. 2017, 110, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.S. Pests of Stored Foodstuffs and Their Control; Kluwer Academic Publishers: New York, NY, USA, 2003. [Google Scholar]
- Rees, D. Insects of Stored Products; Manson Publishing: London, UK, 2004. [Google Scholar]
- Mahroof, R.M.; Hagstrum, D.W. Biology, behavior, and ecology of insects in processed commodities. In Stored Product Protection; Hagstrum, D.W., Phillips, T.W., Cuperus, G., Eds.; Kansas State University: Manhattan, KS, USA, 2012; pp. 33–44. [Google Scholar]
- Mason, L.J.; McDonough, M. Biology, behavior, and ecology of stored grain and legume insects. In Stored Product Protection; Hagstrum, D.W., Phillips, T.W., Cuperus, G., Eds.; Kansas State University: Manhattan, KS, USA, 2012; pp. 7–20. [Google Scholar]
- Hagstrum, D.W.; Klejdysz, T.; Subramanyam, B.H.; Nawrot, J. Atlas of Stored-Product Insects and Mites; AACC International: St. Paul, MN, USA, 2013. [Google Scholar]
- Lowe, S.; Brone, M.; Boudjelas, S.; Poorter De, M. 100 of the World’s Worst Invasive Alien Species. A Selection from the Global Invasive Species Database; Hollands Printing Ltd.: Auckland, New Zealand, 2000. [Google Scholar]
- Kavallieratos, N.G.; Athanassiou, C.G.; Guedes, R.N.C.; Drempela, J.D.; Boukouvala, M.C. Invader competition with local competitors: Displacement or co-existence among the invasive khapra beetle, Trogoderma granarium Everts (Coleoptera: Dermestidae), and two other major stored-grain beetles? Front. Plant Sci. 2017, 8, 1837. [Google Scholar] [CrossRef] [Green Version]
- Kavallieratos, N.G.; Karagianni, E.S.; Papanikolaou, N.E. Life history of Trogoderma granarium Everts (Coleoptera: Dermestidae) on peeled barley, peeled oats and triticale. J. Stored Prod. Res. 2019, 84, 101515. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Athanassiou, C.G.; Boukouvala, M.C.; Tsekos, G.T. Influence of different non-grain commodities on the population growth of Trogoderma granarium Everts (Coleoptera: Dermestidae). J. Stored Prod. Res. 2019, 81, 31–39. [Google Scholar] [CrossRef]
- Myers, S.W.; Hagstrum, D.W. Quarantine. In Stored Product Protection; Hagstrum, D.W., Phillips, T.W., Cuperus, G., Eds.; Kansas State University: Manhattan, KS, USA, 2012; pp. 297–304. [Google Scholar]
- EPPO (European and Mediterranean Plant Protection Organization). EPPO Global Data Base. Trogoderma granarium. Available online: https://gd.eppo.int/taxon/TROGGA (accessed on 27 November 2020).
- Athanassiou, C.G.; Kavallieratos, N.G.; Boukouvala, M.C. Population growth of the khapra beetle, Trogoderma granarium Everts (Coleoptera: Dermestidae) on different commodities. J. Stored Prod. Res. 2016, 69, 72–77. [Google Scholar] [CrossRef]
- Islam, M.S.; Shahjahan, M.; Motaleb, M.A.; Hye, M.A. Laboratory evaluation of some indigenous plant extracts against granary weevil, Sitophilus granarius L. (Coleoptera: Curculionidae). Pak. J. Biol. Sci. 2002, 5, 763–766. [Google Scholar]
- Wakil, W.; Schmitt, T. Field trials on the efficacy of Beauveria bassiana, diatomaceous earth and imidacloprid for the protection of wheat grains from four major stored grain insect pests. J. Stored Prod. Res. 2015, 64, 160–167. [Google Scholar] [CrossRef]
- Honey, S.F.; Bajwa, B.; Mazhar, M.S.; Wakil, W. Trogoderma granarium (Everts) (Coleoptera: Dermestidae), an alarming threat to rice supply chain of Pakistan. Int. J. Entomol. Res. 2017, 5, 23–31. [Google Scholar]
- Ahmedani, M.S.; Shaheen, N.; Ahmedani, M.Y.; Aslam, M. Status of phosphine resistance in khapra beetle, Trogoderma granarium (Everts) strains collected from remote villages of Rawalpindi district. Pak. Entomol. 2007, 29, 95–102. [Google Scholar]
- Wakil, W.; Ghazanfar, M.U.; Yasin, M. Naturally occurring entomopathogenic fungi Infecting stored grain insect species in Punjab, Pakistan. J. Insect Sci 2014, 14, 182. [Google Scholar] [CrossRef] [Green Version]
- FAO (Food and Agriculture Organization of the United Nations). Recommended methods for the detection and measurement of resistance of agricultural pests to pesticides. Tentative method for adults of some major pest species of stored cereals, with methyl bromide and phosphine. FAO method no. 16. Plant Protect. Bull. 1975, 23, 12–25. [Google Scholar]
- Jagadeesan, R.; Collins, P.J.; Daglish, G.J.; Ebert, P.R.; Schlipalius, D.I. Phosphine resistance in the rust red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae): Inheritance, gene interactions and fitness costs. PLoS ONE 2012, 7, e31582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/ETS® 9.1 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2004. [Google Scholar]
- Alam, M.S.; Shaukat, S.S.; Ahmed, M.; Iqbal, S.; Ahmad, A. A survey of resistance to phosphine in some coleopterous pests of stored wheat and rice grain in Pakistan. Pak. J. Biol. Sci. 1999, 2, 623–626. [Google Scholar] [CrossRef] [Green Version]
- Ali, Q.M.; Abbas, M.; Arif, S. Monitoring of resistance against phosphine in stored grain insect pests in Sindh. Middle East J. Sci. Res. 2013, 16, 1501–1507. [Google Scholar]
- Gautam, S.G.; Opit, G.P.; Hosoda, E. Phosphine resistance in adult and immature life stages of Tribolium castaneum (Coleoptera: Tenebrionidae) and Plodia interpunctella (Lepidoptera: Pyralidae) populations in California. J. Econ. Entomol. 2016, 109, 2525–2533. [Google Scholar] [CrossRef]
- Pimentel, M.A.G.; Faroni, L.R.D.A.; Guedes, R.N.C.; Sousa, A.H.; Tótola, M.R. Phosphine resistance in Brazilian populations of Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). J. Stored Prod. Res. 2009, 45, 71–74. [Google Scholar] [CrossRef]
- Hernandez Nopsa, J.F.; Daglish, G.J.; Hagstrum, D.W.; Leslie, J.F.; Phillips, T.W.; Scoglio, C.; Thomas Sharma, S.; Walter, G.H.; Garrett, K.A. Ecological networks in stored grain: Key postharvest nodes for emerging pests, pathogens, and mycotoxins. BioScience 2015, 65, 985–1002. [Google Scholar] [CrossRef] [Green Version]
- Mills, K.A. Resistance to the fumigant hydrogen phosphide in some stored-product species associated with repeated inadequate treatments. Mitt. Dtsch. Ges. Allg. Angew. Entomol. 1983, 4, 98–101. [Google Scholar]
- Tyler, P.S.; Taylor, R.W.; Rees, D.P. Insect resistance to phosphine fumigation in food warehouses in Bangladesh. Int. Pest Control 1983, 25, 10–13. [Google Scholar]
- Taylor, R.W.D.; Halliday, D. The geographical spread of resistance to phosphine by coleopterous pests of stored products. In Proceedings of the British Crop Protection Conference, Pests and Diseases, Brighton Metropole, UK, 17–20 November 1986; British Crop Protection Council: Thornton Heath, UK, 1986; pp. 607–613. [Google Scholar]
- Taylor, R.W.D. Phosphine—A major fumigant at risk. Int. Pest Control 1989, 31, 10–14. [Google Scholar]
- Zettler, L.J.; Cuperus, G.W. Pesticide resistance in Tribolium castaneum (Coleoptera: Tenebrionidae) and Rhyzopertha dominica (Coleoptera: Bostrichidae) in wheat. J. Econ. Entomol. 1990, 83, 1677–1681. [Google Scholar] [CrossRef]
- Sayaboc, P.D.; Gibe, A.J.G.; Caliboso, F.M. Resistance of Rhizopertha dominica (F.) (Coleoptera: Bostrychidae) to phosphine in the Philippines. Philipp. Entomol. 1998, 12, 91–95. [Google Scholar]
- Collins, P.J.; Daglish, G.J.; Nayak, M.K.; Ebert, P.R.; Schlipalius, D.; Chen, W.; Pavic, H.; Lambkin, T.M.; Kopittke, R.; Bridgeman, B.W. Combating resistance to phosphine in Australia. In Proceedings of the International Conference on Controlled Atmosphere and Fumigation in Stored Products, Fresno, CA, USA, 29 October–3 November 2000; Donahaye, E.J., Navarro, S., Leesch, J.G., Eds.; Executive Printing Services: Clovis, CA, USA, 2001; pp. 593–607. [Google Scholar]
- Nayak, M.; Holloway, J.; Pavic, H.; Head, M.; Reid, R.; Collins, P. Developing strategies to manage highly phosphine resistant populations of flat grain beetles in large bulk storages in Australia. In Proceedings of the 10th International Working Conference on Stored-Product Protection, Estoril, Portugal, 27 June–2 July; Carvalho, M.O., Fields, P.G., Adler, C.S., Arthur, F.H., Athanassiou, C.G., Campbell, J.F., Fleurat Lessard, F., Flinn, P.W., Hodges, R.J., Isikber, A.A., et al., Eds.; Julius Kühn Institut: Berlin, Germany, 2010; pp. 396–401. [Google Scholar]
- Pimentel, M.A.G.; Farroni, L.R.D.A.; da Silva, F.H.; Batista, M.D.; Guedes, R.N.C. Spread of phosphine resistance among Brazilian populations of three species of stored product insects. Neotrop. Entomol. 2010, 39, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Emery, R.N.; Nayak, M.K.; Holloway, J.C. Lessons learned from phosphine resistance monitoring in Australia. Stewart Postharvest Rev. 2011, 3, 6. [Google Scholar]
- Schlipalius, D.I.; Valmas, N.; Tuck, A.G.; Jagadeesan, R.; Ma, L.; Kaur, R.; Goldinger, A.; Anderson, C.; Kuang, J.; Zuryn, S.; et al. A core metabolic enzyme mediates resistance to phosphine gas. Science 2012, 338, 807–810. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Collins, P.J.; Duong, T.M.; Schlipalius, D.I.; Ebert, P.R. Genetic conservation of phosphine resistance in the rice weevil Sitophilus oryzae (L.). J. Hered. 2016, 107, 228–237. [Google Scholar] [CrossRef] [Green Version]
- McCulloch, G.A.; Mohankumar, S.; Subramaniam, S.; Sonai Rajan, T.; Rahul, C.; Surendran, R.; Gaurav, R.; Chandrasekaran, S.; Daglish, G.J.; Walter, G.H. Contrasting patterns of phylogeographic structuring in two key beetle pests of stored grain in India and Australia. J. Pest Sci. 2019, 92, 1249–1259. [Google Scholar] [CrossRef]
- Thangaraj, S.R.; McCulloch, G.; Subtharishi, S.; Chandel, R.K.; Debnath, S.; Subramaniam, C.; Walter, G.H.; Subbarayalu, M. Genetic diversity and its geographic structure in Sitophilus oryzae (Coleoptera: Curculionidae) across India—Implications for managing phosphine resistance. J. Stored Prod. Res. 2019, 84, 101512. [Google Scholar] [CrossRef]
- Prikhodko, D.; Zrilyi, O. Pakistan. Review of the Wheat Sector and Grain Storage Issues; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Nayak, M.K.; Falk, M.G.; Emery, R.N.; Collins, P.J.; Holloway, J.C. An analysis of trends, frequencies and factors influencing the development of resistance to phoshine in the red flour beetle Tribolium castaneum (Herbst) in Australia. J. Stored Prod. Res. 2017, 72, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Benhalima, H.; Chaudhry, M.Q.; Mills, K.A.; Price, N.R. Phosphine resistance in stored-product insects collected from various grain storage facilities in Morocco. J. Stored Prod. Res. 2004, 40, 241–249. [Google Scholar] [CrossRef]
- Price, N.R. A comparison of the uptake and metabolism of 32P-radiolabelled phosphine in susceptible and resistant strains of the lesser grain borer (Rhyzopertha dominica). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 1981, 69, 129–131. [Google Scholar] [CrossRef]
- Price, N.R. Active exclusion of phosphine as a mechanism of resistance in Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). J. Stored Prod. Res. 1984, 20, 163–168. [Google Scholar] [CrossRef]
- Chaudhry, M.Q.; Price, N.R. Insect mortality at doses of phosphine which produce equal uptake in susceptible and resistant strains of Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). J. Stored Prod. Res. 1990, 26, 101–107. [Google Scholar] [CrossRef]
- Chaudhry, M.Q. A review of the mechanisms involved in the action of phosphine as an insecticide and phosphine resistance in stored-product insects. Pestic. Sci. 1997, 49, 213–228. [Google Scholar] [CrossRef]
- Collins, P.J.; Emery, R.N.; Wallbank, B.E. Two decades of monitoring and managing phosphine resistance in Australia. In Proceedings of the 8th International Working Conference on Stored-Product Protection, York, UK, 22–26 July 2002; Credland, P.F., Armitage, D.M., Bell, C.H., Cogan, P.M., Highley, E., Eds.; CAB International: Wallingford, UK, 2003; pp. 570–575. [Google Scholar]
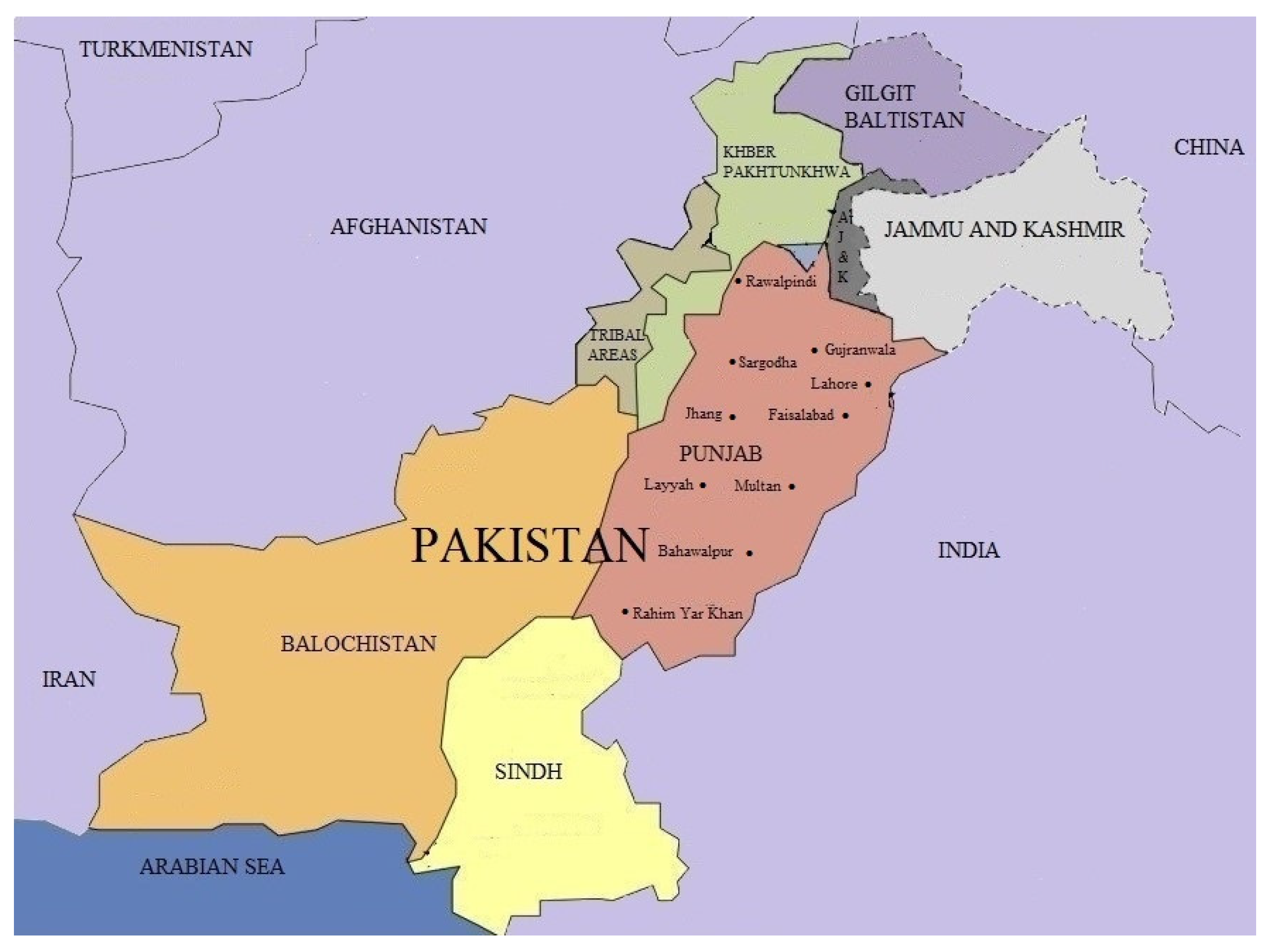

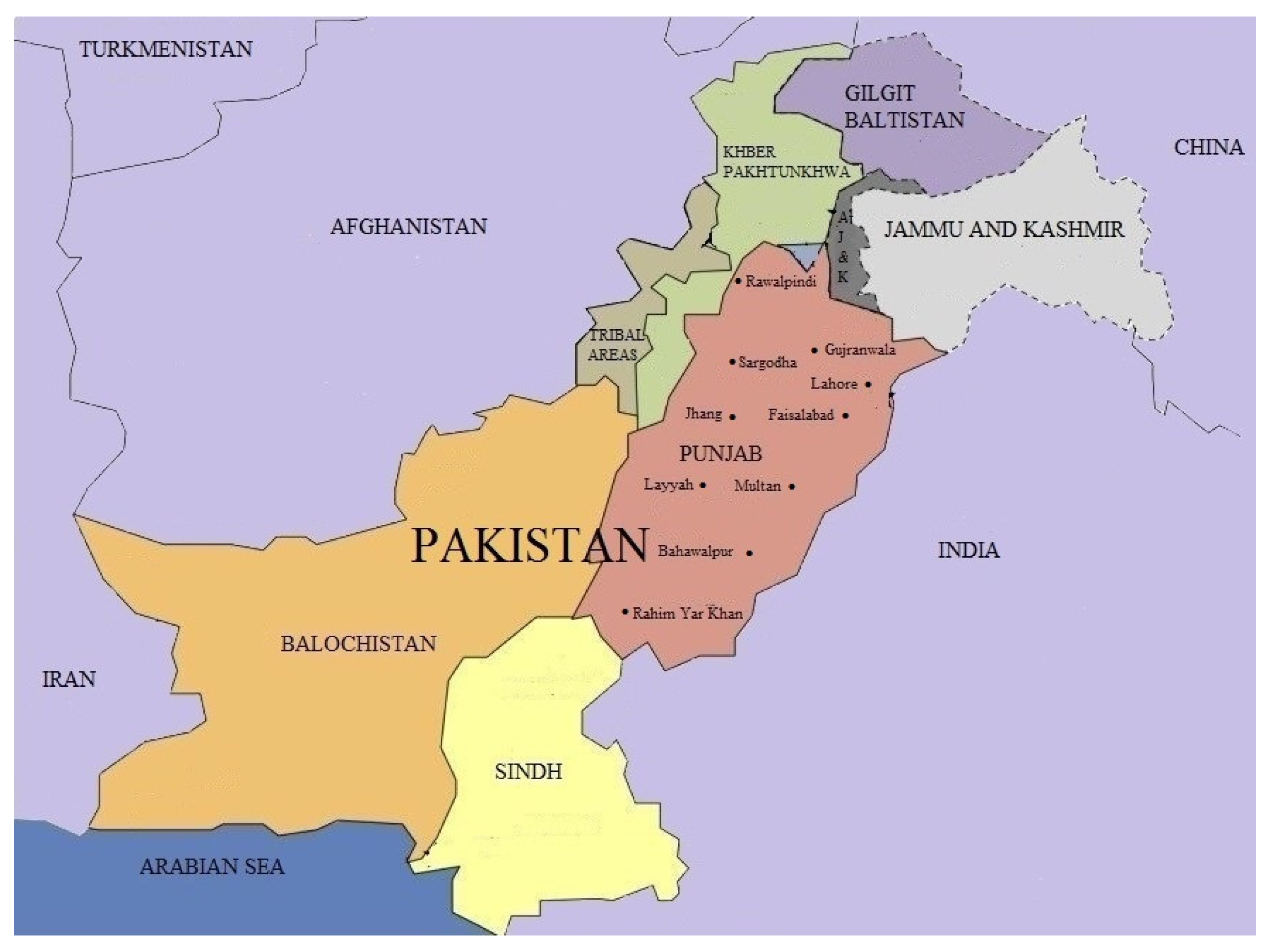{kind=link}
| Region | LC50 (ppm) 95% FL | Slope | Chi-Square | p | Intercept | RR a |
|---|---|---|---|---|---|---|
| Sargodha | 236.32 (163.06–259.16) | 2.35 ± 0.99 | 78.38 | <0.01 | −6.79 ± 1.58 | 82.91 |
| Multan | 333.37 (112.01–415.37) | 3.02 ± 0.92 | 36.57 | <0.01 | −7.63 ± 1.45 | 116.97 |
| Faisalabad | 257.99 (23.21–342.38) | 2.42 ± 0.76 | 71.85 | <0.01 | −21.37 ± 1.98 | 90.52 |
| Bahawalpur | 295.83 (215.23–376.28) | 2.89 ± 1.47 | 53.86 | <0.01 | −7.15 ± 1.90 | 103.80 |
| Rawalpindi | 210.98 (163.06–259.16) | 2.28 ± 0.25 | 26.76 | <0.01 | −5.31 ± 0.62 | 74.02 |
| Layyah | 287.72 (173.37–357.61) | 2.73 ± 0.86 | 62.93 | <0.01 | −6.72 ± 1.82 | 100.95 |
| Lahore | 234.05 (186.14–309.39) | 2.96 ± 0.13 | 57.91 | <0.01 | −7.01 ± 1.96 | 82.12 |
| Rahim Yar Khan | 360.90 (98.92–441.17) | 4.19 ± 0.58 | 46.22 | <0.01 | −11.09 ± 1.12 | 126.67 |
| Jhang | 315.36 (206.20–381.26) | 2.90 ± 1.19 | 47.00 | <0.01 | −7.25 ± 1.16 | 110.65 |
| Gujranwala | 279.88 (171.28–416.52) | 3.31 ± 0.56 | 78.31 | <0.01 | −8.10 ± 1.21 | 98.20 |
| Laboratory | 2.85 (2.62–3.07) | 2.48 ± 0.10 | 7.44 | 0.18 | −1.13 ± 0.07 | - |
| Region | LC50 (ppm) 95% FL | Slope | Chi-Square | p | Intercept | RR a |
|---|---|---|---|---|---|---|
| Sargodha | 86.60 (69.64–103.03) | 2.20 ± 0.15 | 11.05 | 0.05 | −4.26 ± 0.34 | 45.57 |
| Multan | 122.81 (89.15–155.13) | 2.31 ± 0.24 | 26.55 | <0.01 | −4.82 ± 0.56 | 64.63 |
| Faisalabad | 71.26 (54.62–87.26) | 2.10 ± 0.15 | 11.85 | 0.03 | −3.90 ± 0.35 | 37.50 |
| Bahawalpur | 93.72 (75.20–111.68) | 2.15 ± 0.14 | 11.29 | 0.04 | −4.25 ± 0.34 | 49.32 |
| Rawalpindi | 48.58 (27.28–68.61) | 2.03 ± 0.25 | 25.18 | <0.01 | −3.43 ± 0.54 | 25.56 |
| Layyah | 107.29 (81.23–132.49) | 2.33 ± 0.21 | 21.74 | <0.01 | −4.73 ± 0.49 | 56.46 |
| Lahore | 45.96 (28.55–62.43) | 2.06 ± 0.21 | 18.07 | <0.01 | −3.42 ± 0.47 | 24.18 |
| Rahim Yar Khan | 63.17 (44.80–80.62) | 2.19 ± 0.20 | 16.79 | <0.01 | −3.95 ± 0.442 | 33.24 |
| Jhang | 119.37 (84.69–152.65) | 2.25 ± 0.24 | 28.55 | <0.01 | −4.68 ± 0.57 | 62.82 |
| Gujranwala | 66.75 (60.94–80.96) | 2.11 ± 0.14 | 9.92 | 0.07 | −3.85 ± 0.32 | 35.13 |
| Laboratory | 1.90 (1.51–2.28) | 2.21 ± 0.16 | 12.23 | 0.03 | −0.61 ± 0.10 | - |
| Region | LC50 (ppm) 95% FL | Slope | Chi-Square | p | Intercept | RR a |
|---|---|---|---|---|---|---|
| Sargodha | 223.06 (156.78–298.67) | 3.79 ± 0.54 | 79.47 | <0.01 | −8.91 ± 1.97 | 87.81 |
| Multan | 261.00 (24.62–343.96) | 3.38 ± 1.10 | 54.30 | <0.01 | −8.16 ± 1.88 | 102.75 |
| Faisalabad | 202.99 (113.75–260.90) | 2.75 ± 0.73 | 89.89 | <0.01 | −6.36 ±1.51 | 79.91 |
| Bahawalpur | 245.43 (66.41–341.11) | 3.17 ± 0.99 | 48.54 | <0.01 | −7.59 ± 1.71 | 96.62 |
| Rawalpindi | 234.63 (141.27–289.47) | 3.26 ± 1.29 | 67.19 | <0.01 | −7.41± 1.36 | 92.37 |
| Layyah | 305.89 (131.04–374.82) | 3.73 ± 1.04 | 41.34 | <0.01 | −9.27 ± 1.72 | 120.42 |
| Lahore | 186.52 (110.43–243.79) | 2.64 ± 0.43 | 32.91 | <0.01 | −6.00 ± 1.10 | 73.43 |
| Rahim Yar Khan | 256.60 (172.34–345.16) | 3.50 ± 1.48 | 67.50 | <0.01 | −8.45 ± 2.87 | 101.02 |
| Jhang | 275.91 (74.74–355.02) | 3.24 ± 0.95 | 47.26 | <0.01 | −7.91± 2.50 | 108.62 |
| Gujranwala | 213.24 (189.24–265.19) | 2.83 ± 0.50 | 86.26 | <0.01 | −6.61 ± 1.91 | 83.92 |
| Laboratory | 2.54 (2.34–2.75) | 2.41 ± 0.10 | 6.35 | 0.27 | −0.98 ± 0.07 | - |
| Region | LC50 (ppm) 95% FL | Slope | Chi-Square | p | Intercept | RR a |
|---|---|---|---|---|---|---|
| Sargodha | 86.29 (77.48–94.96) | 2.15 ± 0.09 | 6.39 | 0.26 | −4.16 ± 0.22 | 42.93 |
| Multan | 127.21 (115.25–139.05) | 1.95 ± 0.09 | 4.48 | 0.48 | −4.11 ± 0.22 | 63.28 |
| Faisalabad | 100.35 (69.68–129.76) | 1.75 ± 0.18 | 20.01 | <0.01 | −3.51 ± 0.42 | 49.92 |
| Bahawalpur | 131.11 (100.44–161.00) | 2.06 ± 0.18 | 18.18 | <0.01 | −4.36 ± 0.44 | 65.22 |
| Rawalpindi | 75.47 (66.65–84.12) | 1.96 ± 0.09 | 4.17 | 0.52 | −3.69 ± 0.22 | 37.54 |
| Layyah | 169.99 (127.74–212.78) | 1.98 ± 0.21 | 25.57 | <0.01 | −4.42 ± 0.51 | 84.57 |
| Lahore | 74.50 (19.81–126.43) | 1.38 ± 0.28 | 53.47 | <0.01 | −2.59 ± 0.67 | 37.06 |
| Rahim Yar Khan | 117.76 (99.48–135.73) | 2.40 ± 0.14 | 11.00 | 0.05 | −4.97 ± 0.34 | 58.58 |
| Jhang | 145.23 (103.79–185.91) | 1.89 ± 0.21 | 25.01 | <0.01 | −4.10 ± 0.50 | 72.25 |
| Gujranwala | 109.19 (100.36–117.92) | 2.48 ± 0.10 | 5.71 | 0.33 | −5.05 ± 0.23 | 54.32 |
| Laboratory | 2.01 (1.82–2.21) | 2.20 ± 0.18 | 7.81 | 0.16 | −0.67 ± 0.17 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wakil, W.; Kavallieratos, N.G.; Usman, M.; Gulzar, S.; El-Shafie, H.A.F. Detection of Phosphine Resistance in Field Populations of Four Key Stored-Grain Insect Pests in Pakistan. Insects 2021, 12, 288. https://doi.org/10.3390/insects12040288
Wakil W, Kavallieratos NG, Usman M, Gulzar S, El-Shafie HAF. Detection of Phosphine Resistance in Field Populations of Four Key Stored-Grain Insect Pests in Pakistan. Insects. 2021; 12(4):288. https://doi.org/10.3390/insects12040288
Chicago/Turabian StyleWakil, Waqas, Nickolas G. Kavallieratos, Muhammad Usman, Sehrish Gulzar, and Hamadttu A. F. El-Shafie. 2021. "Detection of Phosphine Resistance in Field Populations of Four Key Stored-Grain Insect Pests in Pakistan" Insects 12, no. 4: 288. https://doi.org/10.3390/insects12040288
APA StyleWakil, W., Kavallieratos, N. G., Usman, M., Gulzar, S., & El-Shafie, H. A. F. (2021). Detection of Phosphine Resistance in Field Populations of Four Key Stored-Grain Insect Pests in Pakistan. Insects, 12(4), 288. https://doi.org/10.3390/insects12040288