
Investigating the Parasitoid Community Associated with the Invasive Mealybug Phenacoccus solenopsis in Southern China



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasitoid Collecting and Sampling
DNA Extraction, Amplification, and Sequencing
2.2. Sequence Analysis and Molecular Species Delimitation
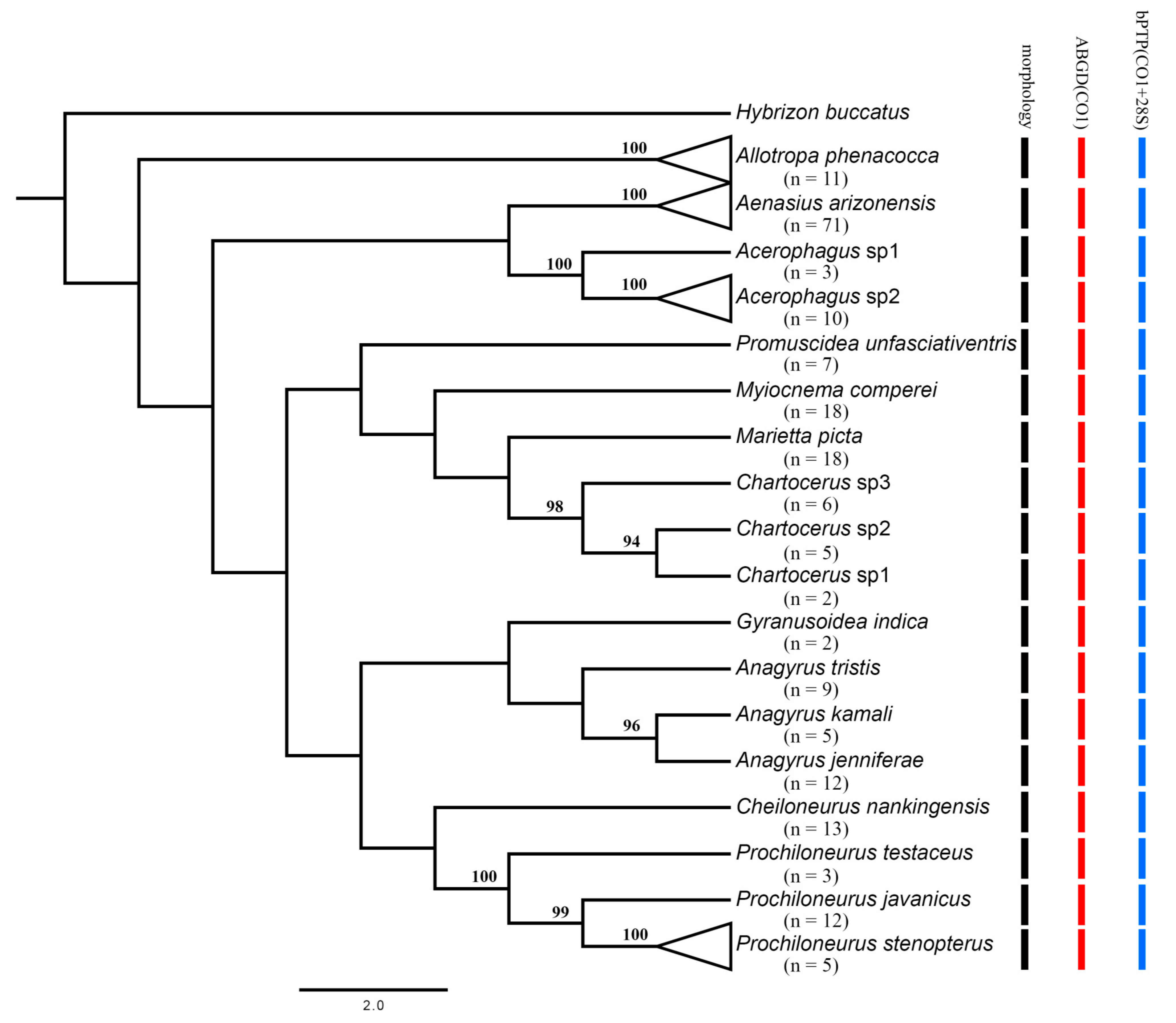
3. Results and Discussion
3.1. BLAST in the NCBI and the Barcode of Life Database (BOLD)
3.2. Genetic Distances
3.3. The Automatic Barcode Gap Discovery (ABGD) Analysis and Bayesian Poisson Tree Processes Model (bPTP)
3.3.1. Aphelinidae
Genus Marietta
Genus Myiocnema
Genus Promuscidea
3.3.2. Encyrtidae
Genus Acerophagus
Genus Aenasius
Genus Anagyrus
Genus Cheiloneurus
Genus Gyranusoidea
Genus Prochiloneurus
3.3.3. Platygasteridae
Genus Allotropa
3.3.4. Signiphoridae
Genus Chartocerus

{kind=link}
{kind=link}
| Family | Species | Parasitoid or Hyperparasitoid | Reference |
|---|---|---|---|
| Aphelinidae | Marietta picta (André, 1878) | parasitoid | [27,28] |
| Myiocnema comperei Ashmead | parasitoid | [27] | |
| Promuscidea unfasciativentris Girault | hyperparasitoid | [19,27] | |
| Encyrtidae | Aenasius arizonensis (Girault) | parasitoid | [18] |
| Acerophagus coccois Smith | parasitoid | [20,27] | |
| Acerophagus sp1 | parasitoid | This study | |
| Acerophagus sp2 | parasitoid | This study | |
| Anagyrus kamali Moursi | parasitoid | [19,27] | |
| Anagyrus agraensis Sarawat | parasitoid | [28] | |
| Anagyrus aligarhensis Agarwal and Alam | parasitoid | [28] | |
| Anagyrus californicus (Compere) | parasitoid | [52] | |
| Anagyrus dactylopii (Howard) | parasitoid | [19] | |
| Anagyrus jenniferae Noyes & Hayat | parasitoid | This study | |
| Anagyrus kamali Moursi | parasitoid | This study | |
| Anagyrus mirzai Agarwal & Alam | parasitoid | [19] | |
| Anagyrus osmoi Guerrieri & Ghahri | parasitoid | [62] | |
| Anagyrus tristis Noyes & Hayat | parasitoid | This study | |
| Bothriothorax serratellus (Dalman) | hyperparasitoid | [28] | |
| Cheiloneurus sp. | parasitoid | [17] | |
| Cheiloneurus nankingensis Li & Xu | hyperparasitoid | [27,29] | |
| Encyrtus aurantii (Geoffroy) | parasitoid | [19] | |
| Gyranusoidea indica Shafee, Alam & Agarwal | parasitoid | This study | |
| Homalotylus albiclavatus (Agarwal) | parasitoid | [19] | |
| Leptomastix algirica Trjapitzin | parasitoid | [52] | |
| Leptomastix dactylopii Howard | parasitoid | [28] | |
| Leptomastix mayri Ózdikmen | parasitoid | [28] | |
| Leptomastix epona (Walker) | parasitoid | [26] | |
| Metaphycus sp. | parasitoid | [22] | |
| Prochiloneurus aegyptiacus (Mercet) | hyperparasitoid | [28] | |
| Prochiloneurus javanicus (Ferriere) | hyperparasitoid | [63]; This study | |
| Prochiloneurus nagasakiensis (Ishii) | hyperparasitoid | [20] | |
| Prochiloneurus nigricornis (Girault) | hyperparasitoid | [27] | |
| Prochiloneurus pulchellus Silvestri | hyperparasitoid | [19,58] | |
| Prochiloneurus rex (Girault) | hyperparasitoid | [28] | |
| Prochiloneurus stenopterus Wang, Huang & Xu | hyperparasitoid | [58]; This study | |
| Prochiloneurus testaceus (Agarwal) | hyperparasitoid | This study | |
| Prochiloneurus uyguni Hayat | hyperparasitoid | [26] | |
| Prochiloneurus sp. | ? hyperparasitoid | This study | |
| Pseudleptomastix squammulata Girault | parasitoid | [52] | |
| Eulophidae | Aprostocetus bangaloricus Narendran | parasitoid | [19] |
| Aprostocetus minutus (Howard) | parasitoid | [17] | |
| Platygastridae | Allotropa phenacocca Chen, Liu & Xu | parasitoid | [21,27] |
| Pteromalidae | Pachyneuron leucopiscida Mani | parasitoid | [19] |
| Signiphoridae | Chartocerus kerrichi (Agarwal) | parasitoid | [19] |
| Chartocerus kurdjumovi (Nikolskaya) | hyperparasitoid | [28] | |
| Chartocerus sp1 | ?hyperparasitoid | This study | |
| Chartocerus sp2 | ?hyperparasitoid | This study | |
| Chartocerus sp3 | ?hyperparasitoid | This study |
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, D.R.; Miller, G.L.; Watson, G.W. Invasive species of mealybugs (Hemiptera: Pseudococcidae) and their threat to US agriculture. Proc. Entomol. Soc. Wash. 2002, 104, 825–836. [Google Scholar]
- Ben-Dov, Y. Note: The solanum mealybug, Phenacoccus solani ferris (Hemiptera: Coccoidea: Pseudococcidae), extends its distribution range in the mediterranean basin. Phytoparasitica 2005, 33, 15–16. [Google Scholar] [CrossRef]
- Wang, Y.P.; Wu, S.A.; Zhang, R.Z. Pest risk analysis of a new invasive pest, Phenacoccus solenopsis, to China. Chin. J. Entomol. 2009, 46, 101–106. [Google Scholar]
- Joshi, M.D.; Butani, P.G.; Patel, V.N.; Jeyakumar, P. Cotton mealy bug, Phenacoccus solenopsis Tinsley—A review. Agric. Rev. 2010, 31, 113–119. [Google Scholar]
- Wang, Y.; Watson, G.W.; Zhang, R. The potential distribution of an invasive mealybug Phenacoccus solenopsis and its threat to cotton in Asia. Agric. For. Èntomol. 2010, 12, 403–416. [Google Scholar] [CrossRef]
- Tong, H.-J.; Ao, Y.; Li, Z.-H.; Wang, Y.; Jiang, M.-X. Invasion biology of the cotton mealybug, Phenacoccus solenopsis Tinsley: Current knowledge and future directions. J. Integr. Agric. 2019, 18, 758–770. [Google Scholar] [CrossRef] [Green Version]
- Abdulrassoul, M.S.; Almalo, I.M.; Hermiz, F.B. First record and host plants of Solenopsis Mealybug, Phenacoccus solenopsis Tinsley, 1898 (Hemiptera: Pseudococcidae) from Iraq. J. Biodivers. Environ. Sci. 2015, 7, 216–222. [Google Scholar]
- Hanchinal, S.G.; Patil, B.V.; Basavanagoud, K.; Nagangoud, A.; Biradar, D.P.; Janagoudar, B.S. Incidence of invasive mealybug (Phenacoccus solenopsis Tinsley) on cotton. Karnataka J. Agric. Sci. 2011, 24, 143–145. [Google Scholar]
- Hodgson, C.; Abbas, G.; Arif, M.J.; Saeed, S.; Karar, H. Phenacoccus solenopsis Tinsley (Sternorrhyncha: Coccoidea: Pseudococcidae), an invasive mealybug damaging cotton in Pakistan and India, with a discussion on seasonal morphological variation. Zootaxa 2008, 1913, 1–35. [Google Scholar] [CrossRef]
- Nagrare, V.; Kranthi, S.; Biradar, V.; Zade, N.; Sangode, V.; Kakde, G.; Shukla, R.; Shivare, D.; Khadi, B.; Kranthi, K. Widespread infestation of the exotic mealybug species, Phenacoccus solenopsis (Tinsley) (Hemiptera: Pseudococcidae), on cotton in India. Bull. Èntomol. Res. 2009, 99, 537–541. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Dai, T.-M.; Tian, H.; Wan, F.-H.; Zhang, G.-F. Range expansion of the invasive cotton mealybug, Phenacoccus solenopsis Tinsley: An increasing threat to agricultural and horticultural crops in China. J. Integr. Agric. 2020, 19, 881–885. [Google Scholar] [CrossRef]
- Fand, B.B.; Suroshe, S.S. The invasive mealybug Phenacoccus solenopsis Tinsley, a threat to tropical and subtropical agricultural and horticultural production systems—A review. Crop. Prot. 2015, 69, 34–43. [Google Scholar] [CrossRef]
- Kaur, H.; Virk, J.S. Feeding potential of Cryptolaemus montrouzieri against the mealybug Phenacoccus solenopsis. Phytoparasitica 2011, 40, 131–136. [Google Scholar] [CrossRef]
- Ejaz, M.; Ullah, S.; Shad, S.A.; Abbas, N.; Binyameen, M. Characterization of inheritance and preliminary biochemical mechanisms of spirotetramat resistance in Phenacoccus solenopsis Tinsley: An economic pest from Pakistan. Pestic. Biochem. Physiol. 2019, 156, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Nagrare, V.S.; Fand, B.B.; Naik, V.C.B.; Naikwadi, B.; Deshmukh, V.; Sinh, D. Resistance development in Cotton mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) to insecticides from Organophosphate, Thiadiazines and Thiourea derivatives. Int. J. Trop. Insect Sci. 2019, 40, 181–188. [Google Scholar] [CrossRef]
- Afzal, M.B.; Shad, S.; Ejaz, M.; Serrao, J. Laboratory selection, cross-resistance, and estimations of realized heritability of indoxacarb resistance in Phenacoccus solenopsis (Homoptera: Pseudococcidae). Pest Manag. Sci. 2020, 76, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, T.W.; Stewart, J.W.; Minzenmayer, R.; Rose, M. First record of Phenacoccus solenopsis Tinsley in cultivated cotton in the United States. Southwest. Entomol. 1991, 16, 215–221. [Google Scholar]
- Hayat, M. Description of a new species of Aenasius Walker (Hymenoptera: Encyrtidae), parasitoid of the mealybug, Phenacoccus solenopsis Tinsley (Homoptera: Pseudococcidae) in India. Biosystematica 2009, 3, 21–26. [Google Scholar]
- Vennila, S.; Ramamurthy, V.V.; Deshmukh, A.; Pinjarkar, D.B.; Agarwal, M.; Pagar, P.C.; Prasad, Y.G.; Prabhakar, M.; Kranthi, K.R.; Bambawale, O.M. A Treatise on Mealybugs of Central Indian Cotton Production System; Technical Bulletin no. 24; National Centre for Integrated Pest Management: New Delhi, India, 2010; 39p. [Google Scholar]
- Chen, H.Y.; He, L.F.; Zheng, C.H.; Li, P.; Yi, Q.H.; Xu, Z.F. Survey on the natural enemies of mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) from Guangdong and Hainan, China. J. Environ. Entomol. 2011, 33, 269–272. [Google Scholar]
- Chen, H.; Liu, J.; Xu, Z. Description of a new platygastrid parasitoidAllotropa phenacocca (Hymenoptera) on Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae). Orient. Insects 2011, 45, 275–280. [Google Scholar] [CrossRef]
- Nagrare, V.S.; Kranthi, S.; Kumar, R.; DharaJothi, B.; Amutha, M.; Deshmukh, A.J.; Bisane, K.D.; Kranthi, K.R. Compendium of Cotton Mealybug; Central Institute for Cotton Research: Nagpur, India, 2011; 42p. [Google Scholar]
- Prasad, Y.G.; Prabhakar, M.; Sreedevi, G.; Thirupathi, M. Spatio-temporal dynamics of the parasitoid, Aenasius bambawalei Hayat (Hymenoptera: Encyrtidae) on mealybug, Phenacoccus solenopsis Tinsley in cotton based cropping systems and associated weed flora. J. Biol. Control 2011, 25, 198–202. [Google Scholar]
- Tanwar, R.K.; Jeyakumar, P.; Singh, A.; Jafri, A.; Bambawale, O.M. Survey for cotton mealybug, Phenacoccus solenopsis (Tinsley) and its natural enemies. J. Environ. Biol. 2011, 32, 381–384. [Google Scholar]
- Arif, M.I.; Rafiq, M.; Wazir, S.; Mehmood, N.; Ghaffar, A. Studies on cotton mealybug, Phenacoccus solenopsis (Pseudococcidae: Homoptera), and its natural enemies in Punjab, Pakistan. Int. J. Agric. Biol. 2012, 14, 557–562. [Google Scholar]
- Çalişkan, A.F.; Hayat, M.; Ulusoy, M.R.; Kaydan, M.B. Parasitoids (Hymenoptera: Encyrtidae) of an invasive mealybug Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) in Turkey. Turk. J. Èntomol. 2016, 40, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Li, J.F.; Deng, J.; Chen, H.Y.; Yang, L.; Zhou, Z.S.; Jiang, J.J.; Huang, L.F.; Gui, F.R.; Chen, H.S. Investigation on the occurrence of parasitic wasps of Phenacoccus solenopsis Tinsley in Guangxi. J. South. Agric. 2020, 51, 853–861. [Google Scholar]
- Torfi, E.T.; Rasekh, A.; Moravvej, S.A.; Mossadegh, M.S.; Rajabpoor, A. Associated Chalcidoidea (Hymenoptera) with the cotton mealybug Phenacoccus solenopsis (Hemiptera: Pseudococcidae) in the southwestern Iran. J. Crop Prot. 2020, 29, 129–139. [Google Scholar]
- Li, Z.; Yao, T.; Xu, Z.; Meng, L.; Li, B. A new species of Cheiloneurus Westwood (Hymenoptera, Encyrtidae) as a hyperparasitoid of the invasive cotton mealybug, Phenacoccus solenopsis Tinsley, in China. ZooKeys 2020, 974, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; Dewaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Boil. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Boil. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [Green Version]
- Venkatesan, T.; Baby, N.L.; Jalali, S.K.; Shylesha, A.N.; Rabindra, R.J. Characterization and identification of Acerophagus papayae Noyes and Schauff (Hymenoptera: Encyrtidae), an introduced parasitoid of papaya mealybug, Paracoccus marginatus Williams and Granara de Willink through DNA barcode. J. Biol. Control 2011, 25, 11–13. [Google Scholar]
- Zhou, Q.-S.; Xi, Y.-Q.; Yu, F.; Zhang, X.; Li, X.-J.; Liu, C.-L.; Niu, Z.-Q.; Zhu, C.-D.; Qiao, G.-X.; Zhang, Y.-Z. Application of DNA barcoding to the identification of Hymenoptera parasitoids from the soybean aphid (Aphis glycines) in China. Insect Sci. 2013, 21, 363–373. [Google Scholar] [CrossRef]
- Malausa, T.; Delaunay, M.; Fleisch, A.; Groussier-Bout, G.; Warot, S.; Crochard, D.; Guerrieri, E.; Delvare, G.; Pellizzari, G.; Kaydan, M.B.; et al. Investigating Biological Control Agents for Controlling Invasive Populations of the Mealybug Pseudococcus comstocki in France. PLoS ONE 2016, 11, e0157965. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Liu, S.; Zhang, Z. Efficacy of using DNA barcoding to identify parasitoid wasps of the melon-cotton aphid (Aphis gossypii) in watermelon cropping system. BioControl 2018, 63, 677–685. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Quicke, D.L.J.; Belshaw, R. Incongruence between Morphological Data Sets: An Example from the Evolution of Endoparasitism Among Parasitic Wasps (Hymenoptera: Braconidae). Syst. Biol. 1999, 48, 436–454. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.C.; Steffen-Campbell, J.D.; Werren, J.H. Phylogeny of the Nasonia species complex (Hymenoptera: Pteromalidae) inferred from an internal transcribed spacer (ITS2) and 28S rDNA sequences. Insect Mol. Biol. 1994, 2, 225–237. [Google Scholar] [CrossRef]
- Gillespie, J.J.; Munro, J.B.; Heraty, J.M.; Yoder, M.J.; Owen, A.K.; Carmichael, A.E. A Secondary Structural Model of the 28S rRNA Expansion Segments D2 and D3 for Chalcidoid Wasps (Hymenoptera: Chalcidoidea). Mol. Biol. Evol. 2005, 22, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Belshaw, R.; Quicke, D.L. A Molecular Phylogeny of the Aphidiinae (Hymenoptera: Braconidae). Mol. Phylogene. Evol. 1997, 7, 281–293. [Google Scholar] [CrossRef]
- Belshaw, R.; Lopez-Vaamonde, C.; Degerli, N.; Quicke, D.L. Paraphyletic taxa and taxonomic chaining: Evaluating the classification of braconine wasps (Hymenoptera: Braconidae) using 28S D2–3 rDNA sequences and morphological characters. Biol. J. Linn. Soc. 2001, 73, 411–424. [Google Scholar] [CrossRef]
- Chesters, U.; Wang, Y.; Yu, F.; Bai, M.; Zhang, T.-X.; Hu, H.-Y.; Zhu, C.-D.; Li, C.-D.; Zhang, Y.-Z. The Integrative Taxonomic Approach Reveals Host Specific Species in an Encyrtid Parasitoid Species Complex. PLoS ONE 2012, 7, e37655. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2011, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.S.; Krogmann, L.; Mayer, C.; Donath, A.; Gunkel, S.; Meusemann, K.; Kozlov, A.; Podsiadlowski, L.; Petersen, M.; Lanfear, R.; et al. Evolutionary History of the Hymenoptera. Curr. Biol. 2017, 27, 1013–1018. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Y.Y.; Wang, Z.H.; Huang, J. A new record genus and species of Aphelinidae (Hymnoptera, Chalcidoidea) from China. Acta Zootaxon. Sin. 2012, 37, 456–459. [Google Scholar]
- Suroshe, S.S.; Gautam, R.D.; Fand, B.B. Natural enemy complex associated with the mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) infesting different host plants in India. J. Biol. Control 2013, 27, 204–210. [Google Scholar]
- Chen, H.Y.; Cao, R.X.; Xu, Z.F. First record of Aenasius bambawalei Hayat (Hymenoptera: Encyrtidae), a parasitoid of the mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) from China. J. Environ. Entomol. 2010, 32, 280–282. [Google Scholar]
- Yu, H.B.; Liang, W.S.; Fang, T.S.; Pan, Z.P. Investigation of damage and its natural enemies of Phenacoccus solenopsis Tinsley on garden plants in Guangdong Province. J. Environ. Entomol. 2015, 37, 1109–1112. [Google Scholar]
- Spodek, M.; Ben-Dov, Y.; Mondaca, L.; Protasov, A.; Erel, E.; Mendel, Z. The cotton mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) in Israel: Pest status, host plants and natural enemies. Phytoparasitica 2018, 46, 45–55. [Google Scholar] [CrossRef]
- Peronti, A.L.B.G.; Martinelli, N.M.; Alexandrino, J.G.; Júnior, A.L.M.; Penteado-Dias, A.M.; Almeida, L.M. Natural Enemies Associated with Maconellicoccus hirsutus (Hemiptera: Pseudococcidae) in the State of São Paulo, Brazil. Fla. Èntomol. 2016, 99, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Noyes, J.S.; Hayat, M. A review of the genera of Indo-Pacific Encyrtidae (Hymenoptera: Chalcidoidea). Bull. Br. Mus. Nat. Hist. Entomol. Ser. 1984, 48, 131–395. [Google Scholar]
- Xu, Z.H.; Huang, J. Chinese Fauna of Parasitic Wasps on Scale Insects; Shanghai Scientific & Technical Publishers: Shanghai, China, 2004; 524p. [Google Scholar]
- Zhang, Y.Z.; Huang, D.W. A Review and an Illustrated Key to Genera of Encyrtidae (Hymenoptera: Chalcidoidea) from China; Science Press: Beijing, China, 2004; 95p. [Google Scholar]
- Hayat, M. Indian Encyrtidae (Hymenoptera: Chalcidoidea); Department of Zoology, Aligarh Muslim University: Aligarh, India, 2006; 496p. [Google Scholar]
- Wang, Z.H.; Huang, J.; Xu, Z.H. Chinese species of the genus Prochiloneurus Silvestri with description of a new species (Hymenoptera: Encyrtidae). Zoolog. Syst. 2014, 39, 424–432. [Google Scholar]
- Sureshan, P.; Narendran, T. Description of a new species of Ophelosia Riley with synonymic and taxonomic notes on Indian Eunotinae (Hymenoptera: Chalcidoidea: Pteromalidae). Zoos Print J. 2005, 20, 1854–1855. [Google Scholar] [CrossRef]
- Pillai, K.G.; Krishnamoorthy, A.; Gangavisalakshy, P.N. First report of hyperparasitoids of Anagyrus dactylopii (Howard) from India. J. Biol. Control 2009, 23, 193–194. [Google Scholar]
- Noyes, J.S. Universal Chalcidoidea Database. Available online: http://www.nhm.ac.uk/research-curation/projects/chalcidoids (accessed on 30 August 2020).
- Guerrierri, E.; Ghahari, H. New records, descriptions and notes on Encyrtidae (Hymenoptera: Chalcidoidea) from Iran. Zootaxa 2018, 4444, 316–326. [Google Scholar] [CrossRef]
- Joshi, S.; Rameshkumar, A.; Mohanraj, P. New host-parasitoid associations for some coccids (Hemiptera: Coccoidea) from India. J. Èntomol. Res. 2017, 41, 177. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.-Y.; Li, H.-L.; Pang, H.; Zhu, C.-D.; Zhang, Y.-Z. Investigating the Parasitoid Community Associated with the Invasive Mealybug Phenacoccus solenopsis in Southern China. Insects 2021, 12, 290. https://doi.org/10.3390/insects12040290
Chen H-Y, Li H-L, Pang H, Zhu C-D, Zhang Y-Z. Investigating the Parasitoid Community Associated with the Invasive Mealybug Phenacoccus solenopsis in Southern China. Insects. 2021; 12(4):290. https://doi.org/10.3390/insects12040290
Chicago/Turabian StyleChen, Hua-Yan, Hong-Liang Li, Hong Pang, Chao-Dong Zhu, and Yan-Zhou Zhang. 2021. "Investigating the Parasitoid Community Associated with the Invasive Mealybug Phenacoccus solenopsis in Southern China" Insects 12, no. 4: 290. https://doi.org/10.3390/insects12040290
APA StyleChen, H. -Y., Li, H. -L., Pang, H., Zhu, C. -D., & Zhang, Y. -Z. (2021). Investigating the Parasitoid Community Associated with the Invasive Mealybug Phenacoccus solenopsis in Southern China. Insects, 12(4), 290. https://doi.org/10.3390/insects12040290