
Mating Disruption for Managing the Honeydew Moth, Cryptoblabes gnidiella (Millière), in Mediterranean Vineyards

 ,
, 
 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Adult Captures and Evaluation of Infestation Levels
2.3. Pheromone Release Rate Over the Season
2.4. Statistical Analysis
3. Results
3.1. Adult Flights
3.2. Pheromone Release Rate over the Season
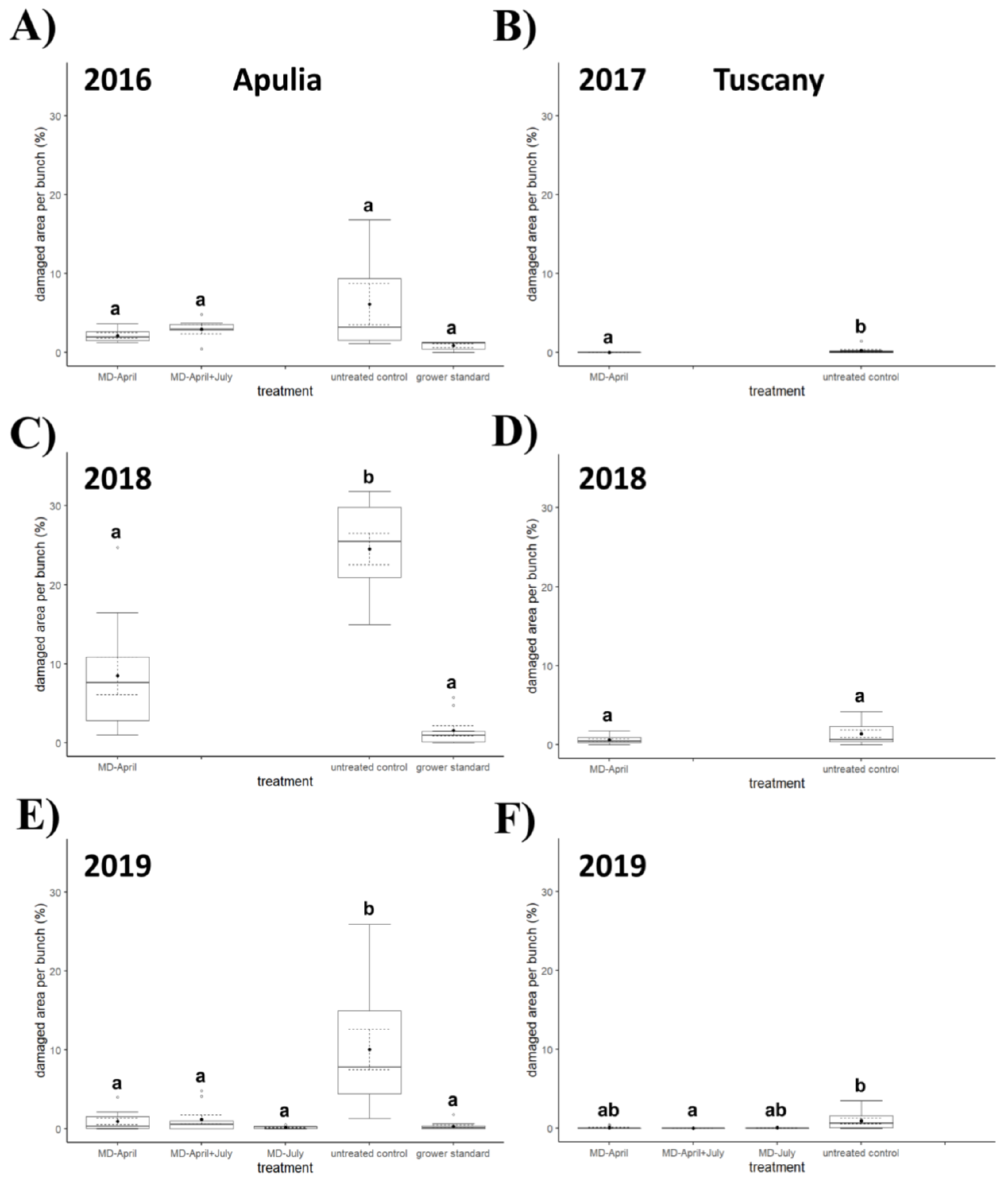
3.3. Field Experiments in Central Italy
3.4. Field Experiments in Southern Italy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bagnoli, B.; Lucchi, A. Bionomics of Cryptoblabes gnidiella (Millière) (Pyralidae Phycitinae) in Tuscan Vineyards. IOBC-WPRS Bull. 2001, 24, 79–84. [Google Scholar]
- Harari, A.R.; Zahavi, T.; Gordon, D.; Anshelevich, L.; Harel, M.; Ovadia, S.; Dunkelblum, E. Pest Management Programmes in Vineyards Using Male Mating Disruption. Pest Manag. Sci. 2007, 63, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, A.; Ricciardi, R.; Benelli, G.; Bagnoli, B. What Do We Really Know on the Harmfulness of Cryptoblabes gnidiella (Millière) to Grapevine? From Ecology to Pest Management. Phytoparasitica 2019, 47, 1–15. [Google Scholar] [CrossRef]
- CABI Cryptoblabes gnidiella (Citrus Pyralid). Available online: https://www.cabi.org/isc/datasheet/16381 (accessed on 22 January 2021).
- Ioriatti, C.; Lucchi, A. Semiochemical Strategies for Tortricid Moth Control in Apple Orchards and Vineyards in Italy. J. Chem. Ecol. 2016, 42, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, A.; Sambado, P.; Juan Royo, A.B.; Bagnoli, B.; Conte, G.; Benelli, G. Disrupting Mating of Lobesia botrana Using Sex Pheromone Aerosol Devices. Environ. Sci. Pollut. Res. Int. 2018, 25, 22196–22204. [Google Scholar] [CrossRef]
- Thiéry, D.; Louâpre, P.; Muneret, L.; Rusch, A.; Sentenac, G.; Vogelweith, F.; Iltis, C.; Moreau, J. Biological Protection against Grape Berry Moths. A Review. Agron. Sustain. Dev. 2018, 38, 15. [Google Scholar] [CrossRef] [Green Version]
- Lucchi, A.; Suma, P.; Ladurner, E.; Iodice, A.; Savino, F.; Ricciardi, R.; Cosci, F.; Marchesini, E.; Conte, G.; Benelli, G. Managing the Vine Mealybug, Planococcus ficus, through Pheromone-Mediated Mating Disruption. Environ. Sci. Pollut. Res. 2019, 26, 10708–10718. [Google Scholar] [CrossRef]
- Wysoki, M.; Izhar, Y.; Gurevitz, E.; Swirski, E.; Greenberg, S. Control of the Honeydew Moth, Cryptoblabes gnidiella Mill. (Lepidoptera: Phycitidae), with Bacillus thuringiensis Berliner in Avocado Plantations. Phytoparasitica 1975, 3, 103–111. [Google Scholar] [CrossRef]
- Lucchi, A.; Ladurner, E.; Iodice, A.; Savino, F.; Ricciardi, R.; Cosci, F.; Conte, G.; Benelli, G. Eco-Friendly Pheromone Dispensers—a Green Route to Manage the European Grapevine Moth? Environ. Sci. Pollut. Res. 2018, 25, 9426–9442. [Google Scholar] [CrossRef] [PubMed]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex Pheromones and Their Impact on Pest Management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef]
- Rodriguez-Saona, C.R.; Stelinski, L.L. Behavior-Modifying Strategies in IPM: Theory and Practice. In Integrated Pest Management: Innovation-Development Process; Peshin, R., Dhawan, A.K., Eds.; Springer: Dordrecht, The Netherlands, 2009; Volume 1, pp. 263–315. ISBN 978-1-4020-8992-3. [Google Scholar]
- Cardé, R.T.; Haynes, K.F. Structure of the pheromone communication channel in moths. In Advances in Insect Chemical Ecology; Millar, J.G., Cardé, R.T., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 283–332. ISBN 978-0-521-79275-2. [Google Scholar]
- Cardé, R.T.; Willis, M.A. Navigational Strategies Used by Insects to Find Distant, Wind-Borne Sources of Odor. J. Chem. Ecol. 2008, 34, 854–866. [Google Scholar] [CrossRef]
- Cardé, R.T.; Minks, A.K. Control of Moth Pests by Mating Disruption: Successes and Constraints. Annu. Rev. Entomol. 1995, 40, 559–585. [Google Scholar] [CrossRef]
- Suckling, D.M. Issues Affecting the Use of Pheromones and Other Semiochemicals in Orchards. Crop. Prot. 2000, 19, 677–683. [Google Scholar] [CrossRef]
- Lo Verde, G.; Guarino, S.; Barone, S.; Rizzo, R. Can Mating Disruption Be a Possible Route to Control Plum Fruit Moth in Mediterranean Environments? Insects 2020, 11, 589. [Google Scholar] [CrossRef]
- Mori, B.A.; Evenden, M.L. When Mating Disruption Does Not Disrupt Mating: Fitness Consequences of Delayed Mating in Moths. Entomol. Exp. Appl. 2013, 146, 50–65. [Google Scholar] [CrossRef]
- Onufrieva, K.S.; Hickman, A.D.; Leonard, D.S.; Tobin, P.C. Ground Application of Mating Disruption against the Gypsy Moth (Lepidoptera: Erebidae). J. Appl. Entomol. 2019, 143, 1154–1160. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.L.; Starcher, S.; Chandran, R.S. Comparison of Mating Disruption and Insecticide Application for Control of Peachtree Borer and Lesser Peachtree Borer (Lepidoptera: Sesiidae) in Peach. Insects 2020, 11, 658. [Google Scholar] [CrossRef] [PubMed]
- Trematerra, P.; Colacci, M.; Athanassiou, C.G.; Kavallieratos, N.G.; Rumbos, C.I.; Boukouvala, M.C.; Nikolaidou, A.J.; Kontodimas, D.C.; Benavent-Fernández, E.; Gálvez-Settier, S. Evaluation of Mating Disruption For the Control of Thaumetopoea pityocampa (Lepidoptera: Thaumetopoeidae) in Suburban Recreational Areas in Italy and Greece. J. Econ. Entomol. 2019, 112, 2229–2235. [Google Scholar] [CrossRef]
- Jallow, M.F.A.; Dahab, A.A.; Albaho, M.S.; Devi, V.Y.; Jacob, J.; Al-Saeed, O. Efficacy of Mating Disruption Compared with Chemical Insecticides for Controlling Tuta absoluta (Lepidoptera: Gelechiidae) in Kuwait. Appl. Entomol. Zool. 2020, 55, 213–221. [Google Scholar] [CrossRef]
- Kong, W.; Wang, Y.; Guo, Y.; Chai, X.; Niu, G.; Li, Y.; Ma, R.; Li, J. Effects of Disruption of Grapholita molesta (Lepidoptera: Tortricidae) Using Sex Pheromone on Moth Pests and Insect Communities in Orchards. Appl. Entomol. Zool. 2020, 55, 367–377. [Google Scholar] [CrossRef]
- Angeli, G.; Anfora, G.; Baldessari, M.; Germinara, G.S.; Rama, F.; De Cristofaro, A.; Ioriatti, C. Mating Disruption of Codling Moth Cydia pomonella with High Densities of Ecodian Sex Pheromone Dispensers. J. Appl. Entomol. 2007, 131, 311–318. [Google Scholar] [CrossRef]
- Cocco, A.; Muscas, E.; Mura, A.; Iodice, A.; Savino, F.; Lentini, A. Influence of Mating Disruption on the Reproductive Biology of the Vine Mealybug, Planococcus ficus (Hemiptera: Pseudococcidae), under Field Conditions. Pest Manag. Sci. 2018, 74, 2806–2816. [Google Scholar] [CrossRef]
- Anfora, G.; Baldessari, M.; De Cristofaro, A.; Germinara, G.S.; Ioriatti, C.; Reggiori, F.; Vitagliano, S.; Angeli, G. Control of Lobesia botrana (Lepidoptera: Tortricidae) by Biodegradable Ecodian Sex Pheromone Dispensers. J. Econ. Entomol. 2008, 101, 444–450. [Google Scholar] [CrossRef]
- Hummel, H.E. A Brief Review on Lobesia botrana Mating Disruption by Mechanically Distributing and Releasing Sex Pheromones from Biodegradable Mesofiber Dispensers. Biochem. Mol. Biol. J. 2017, 3. [Google Scholar] [CrossRef]
- Hashem, A.G.; Tadros, A.W.; Abo-Sheasha, M.A. (Ministry of A. Monitoring the Honeydew Moth, Cryptoblabes gnidiella Mill in Citrus, Mango and Grapevine Orchards, Lepidoptera: Pyralidae. Ann. Agric. Sci. Ain Shams Univ. Egypt 1997, 42, 335–343. [Google Scholar]
- Yehuda, S.B.; Wysoki, M.; Rosen, D. Phenology of the Honeydew Moth, Cryptoblabes gnidiella (Millière) (Lepidoptera: Pyralidae), on Avocado in Israel. Isr. J. Entomol. 1991, 45, 149–160. [Google Scholar]
- Abdel-Moaty, R.M.; Hashim, S.M.; Tadros, A.W. Monitoring the Honeydew Moth, Cryptoblabes gnidiella Millière (Lepidoptera: Pyralidae) in Pomegranate Orchards in the Northwestern Region of Egypt. J. Plant. Prot. Pathol. 2017, 8, 505–509. [Google Scholar] [CrossRef]
- Demirel, N. Seasonal Flight Patterns of the Honeydew Moth, Cryptoblabes gnidiella Millière (Lepidoptera: Pyralidae) in Pomegranate Orchards as Observed Using Pheromone Traps. Entomol. Appl. Sci. Lett. 2016, 3, 1–5. [Google Scholar]
- Arn, H.; Rauscher, S.; Schmid, A.; Jaccard, C.; Bierl-Leonhardt, B.A. Field Experiments to Develop Control of the Grape Moth, Eupoecilia ambiguella, by Communication Disruption. In Management of Insect Pests with Semiochemicals: Concepts and Practice; Mitchell, E.R., Ed.; Springer: Boston, MA, USA, 1981; pp. 327–338. ISBN 978-1-4613-3216-9. [Google Scholar]
- Franco, J.C.; Lucchi, A.; Mendel, Z.; Suma, P.; Vacas, S.; Mansour, R.; Navarro-Llopis, V. Scientific and Technological Developments in Mating Disruption of Scale Insects. Entomol. Gen. 2021. accepted. [Google Scholar] [CrossRef]
- Sellanes, C.; González, A. The Potential of Sex Pheromones Analogs for the Control of Cryptoblabes gnidiella (Lepidoptera: Pyralidae), an Exotic Pest in South America. IOBC WPRS Bull. 2014, 99, 55–60. [Google Scholar]
- Acín, P. Management of the honeydew moth by mating disruption in vineyard. In Proceedings of the Joint Meeting of the IOBC-WPRS Working Groups “Pheromones and Other Semiochemicals in Integrated Production” & “Integrated Protection of Fruit Crops”, Lisbon, Portugal, 20–25 January 2019. IOBC WPRS Bull. 2019, 146, 28–31. [Google Scholar]
- European and Mediterranean Plant Protection Organization Principles of Efficacy Evaluation for Mating Disruption Pheromones. EPPO Bull. 2019, 49, 426–430. [CrossRef]
- Ioriatti, C.; Bagnoli, B.; Lucchi, A.; Veronelli, V. Vine Moths Control by Mating Disruption in Italy: Results and Future Prospects. Redia 2004, 87, 117–128. [Google Scholar]
- Miller, J.R.; Gut, L.J. Mating Disruption for the 21st Century: Matching Technology with Mechanism. Environ. Entomol. 2015, 44, 427–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elnagar, H.M. Population Dynamic of Honeydew Moth, Cryptoblabes gnidiella Miller in Vineyards Orchards. Egypt. Acad. J. Biol. Sci. Entomol. 2018, 11, 73–78. [Google Scholar] [CrossRef]
- Barker, B.; Coop, L. Honeydew Moth Cryptoblabes gnidiella (Lepidoptera: Pyralidae). Phenology/Degree-Day and Climate Suitability Model Analysis; Prepared for USDA APHIS PPQ. Version 1.0, 12/09/2019; Department of Horticulture and Integrated Plant Protection Center, Oregon State University: Corvallis, OR, USA, 2019. [Google Scholar]
- Louis, F.; Schirra, K.-J. Mating Disruption of Lobesia botrana (Lepidoptera: Tortricidae) in Vineyards with Very High Population Densities. IOBC-WPRS Bull. 2001, 24, 75–79. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Variety | Harvesting Period | Row Spacing (m) | Space within Rows (m) |
|---|---|---|---|---|
| Tuscany (Capalbio) 42°25′51.78″ N 11°25′4.82″ E | Cabernet Sauvignon | End of September | 1.3 | 0.6 |
| Apulia (Minervino Murge) 41°8′56.72″ N 16°2′16.09″ E | Aglianico | Middle of October | 2.3 | 0.8 |
| Year | Site | Treatements | ||||
|---|---|---|---|---|---|---|
| 2016 | Minervino Murge | A | AJ | - | C | GS |
| 2017 | Capalbio | A | - | - | C | - |
| 2018 | Minervino Murge | A | - | - | C | GS |
| Capalbio | A | - | - | C | - | |
| 2019 | Minervino Murge | A | AJ | J | C | GS |
| Capalbio | A | AJ | J | C | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ricciardi, R.; Di Giovanni, F.; Cosci, F.; Ladurner, E.; Savino, F.; Iodice, A.; Benelli, G.; Lucchi, A. Mating Disruption for Managing the Honeydew Moth, Cryptoblabes gnidiella (Millière), in Mediterranean Vineyards. Insects 2021, 12, 390. https://doi.org/10.3390/insects12050390
Ricciardi R, Di Giovanni F, Cosci F, Ladurner E, Savino F, Iodice A, Benelli G, Lucchi A. Mating Disruption for Managing the Honeydew Moth, Cryptoblabes gnidiella (Millière), in Mediterranean Vineyards. Insects. 2021; 12(5):390. https://doi.org/10.3390/insects12050390
Chicago/Turabian StyleRicciardi, Renato, Filippo Di Giovanni, Francesca Cosci, Edith Ladurner, Francesco Savino, Andrea Iodice, Giovanni Benelli, and Andrea Lucchi. 2021. "Mating Disruption for Managing the Honeydew Moth, Cryptoblabes gnidiella (Millière), in Mediterranean Vineyards" Insects 12, no. 5: 390. https://doi.org/10.3390/insects12050390
APA StyleRicciardi, R., Di Giovanni, F., Cosci, F., Ladurner, E., Savino, F., Iodice, A., Benelli, G., & Lucchi, A. (2021). Mating Disruption for Managing the Honeydew Moth, Cryptoblabes gnidiella (Millière), in Mediterranean Vineyards. Insects, 12(5), 390. https://doi.org/10.3390/insects12050390