
Guarding Vibrations—Axestotrigona ferruginea Produces Vibrations When Encountering Non-Nestmates


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Species
2.2. Vibration Recordings
2.2.1. In the Nest
2.2.2. At the Entrance Tube
2.2.3. In the Arena
2.3. Vibration Analysis
3. Results
3.1. Production of Vibrations at the Nest
3.2. Arena Experiments
4. Discussion
Vibrational Signaling?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grüter, C. Evolution and diversity of stingless bees. In Stingless Bees. Their Behaviour, Ecology and Evolution, 1st ed.; Grüter, C., Ed.; Springer: Cham, Switzerland, 2020; pp. 43–86. [Google Scholar]
- Peters, R.S.; Krogmann, L.; Mayer, C.; Donath, A.; Gunkel, S.; Meusemann, K.; Kozlov, A.; Podsiadlowski, L.; Petersen, M.; Lanfear, R.; et al. Evolutionary history of the Hymenoptera. Curr. Biol. 2017, 27, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jarau, S. Chemical communication during food exploitation in stingless bees. In Food Exploitation by Social Insects. Ecological, Behavioral, and Theoretical Approaches, 1st ed.; Jarau, S., Hrncir, M., Eds.; CRC Press/Taylor and Francis Group: Bora Raton, FL, USA, 2009; pp. 223–249. [Google Scholar]
- Hill, P.S.M. Stretching the paradigm or building a new? Development of a cohesive language for vibrational communication. In Studying Vibrational Communication, 1st ed.; Cocroft, R.B., Gogala, M., Hill, P.S.M., Wessel, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 13–30. [Google Scholar]
- Hrncir, M.; Barth, F.G. Vibratory communication in stingless bees (Meliponini): The challenge of interpreting the signals. In Studying Vibrational Communication, 1st ed.; Cocroft, R.B., Gogala, M., Hill, P.S.M., Wessel, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 349–374. [Google Scholar]
- Krausa, K.; Hager, F.A.; Kircher, W.H. The effect of food profitability on foraging behaviors and vibrational signals in the African stingless bee Plebeina Hildebrandti. Insect. Soc. 2017, 64, 567–578. [Google Scholar] [CrossRef]
- Barth, F.G.; Hrncir, M.; Jarau, S. Signals and cues in the recruitment behaviour of stingless bees. J. Comp. Physiol. 2008, 194, 313–327. [Google Scholar] [CrossRef]
- Brewer, S.M.; McGrew, W.C. Chimpanzee use of a tool-set to get honey. Folia Primatol. 1990, 54, 100–104. [Google Scholar] [CrossRef]
- Kajobe, R.; Roubik, D.W. Honey-making bee colony abundance and predation by apes and humans in a Uganda forest reserve. Biotropica 2006, 38, 210–218. [Google Scholar] [CrossRef]
- Kiatoko, N. Distribution, Behavioural Biology, Rearing and Pollination Efficiency of Five Stingless Bee Species (Apidae: Meliponinae) in Kakamega Forest, Kenya. Ph.D. Thesis, Kenyatta University, Nairobi, Kenya, 2012. [Google Scholar]
- Grüter, C.; von Zuben, L.G.; Segers, F.H.I.D.; Cunningham, J.P. Warfare in stingless bees. Insect Soc. 2016, 63, 223–236. [Google Scholar] [CrossRef]
- van Zweden, J.; d’Ettorre, P. Nestmate recognition in social insects and the role of hydrocarbons. In Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology, 1st ed.; Blomquist, G.J., Bagnères, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 222–243. [Google Scholar]
- Leonhardt, S.D. Chemical Ecology of Stingless Bees. J. Chem. Ecol. 2017, 43, 385–402. [Google Scholar] [CrossRef]
- Grüter, C. Enemies, dangers and colony defence. In Stingless Bees. Their Behaviour, Ecology and Evolution, 1st ed.; Grüter, C., Ed.; Springer: Cham, Switzerland, 2020; pp. 233–272. [Google Scholar]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioural, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef]
- Monnin, T. Chemical recognition of reproductive status in social insects. Ann. Zool Fenn. 2006, 43, 515–530. [Google Scholar]
- Couvillon, M.; Ratnieks, F.L.W. Odour transfer in stingless bee marmelada (Frieseomelitta varia) demonstrates that entrance guards use an “undesirable-absent” recognition system. Behav. Ecol. Sociobiol. 2008, 62, 1099–1105. [Google Scholar] [CrossRef]
- Blum, M.S. Alarm pheromones. Annu. Rev. Entomol. 1969, 14, 57–80. [Google Scholar] [CrossRef]
- Smith, B.H.; Roubik, D.W. Mandibular glands of stingless bees (Hymenoptera: Apidae): Chemical analysis of their contents and biological function of two species of Melipona. J. Chem. Ecol. 1983, 9, 1465–1472. [Google Scholar] [CrossRef]
- Cruz-López, L.; Malo, E.A.; Morgan, E.D.; Rincon, M.; Guzmán, M.; Rojas, J.C. Mandibular gland secretion of Melipona beecheii: Chemistry and behavior. J. Chem. Ecol. 2005, 31, 1621–1632. [Google Scholar] [CrossRef]
- Schorkopf, D.L.P.; Hrncir, M.; Mateus, S.; Zucchi, R.; Schmidt, V.M.; Barth, F.G. Mandibular gland secretions of meliponine worker bees: Further evidence for their role in interspecific and intraspecific defence and aggression and against their role in food source signaling. J. Exp. Biol. 2009, 212, 1153–1162. [Google Scholar] [CrossRef]
- Hrncir, M.; Barth, F.G.; Tautz, J. Vibratory and airborne sound signals in bee communication (Hymenoptera). In Insect Sound and Communication—Physiology, Behaviour, Ecology and Evolution, 1st ed.; Drosopoulos, S., Claridge, M.F., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 421–436. [Google Scholar]
- Hager, F.A.; Krausa, K.; Kirchner, W.H. Vibrational behavior in termites (Isoptera). In Biotremology: Studying Vibrational Behavior, 1st ed.; Hill, P.S.M., Lakes-Harlan, R., Mazzoni, V., Narins, P.M., Virant-Doberlet, M., Wessel, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 309–327. [Google Scholar]
- Kirchner, W.H. Acoustical communication in social insects. In Orientation and Communication in Arthropods, 1st ed.; Lehrer, M., Ed.; Birkhäuser Verlag: Basel, Switzerland, 1997; pp. 273–300. [Google Scholar]
- Haas, A. Das Rätsel des Hummeltrompeters: Lichtalarm. Z. Tierpsychol. 1961, 18, 129–138. [Google Scholar]
- Schneider, P. Akustische Signale bei Hummeln. Naturwissenschaften 1972, 59, 168. [Google Scholar] [CrossRef]
- Schneider, P. Versuche zur Erzeugung des Verteidigungstones bei Hummeln. Zool. Jb Physiol. 1975, 79, 11–127. [Google Scholar]
- Kirchner, W.H.; Röschard, J. Hissing in bumblebees: An interspecific defence signal. Insect Soc. 1999, 46, 239–243. [Google Scholar] [CrossRef]
- Wagner, W. Psychobiologische Untersuchungen an Hummeln. Zoologica 1907, 19, 1–239. [Google Scholar]
- Lindauer, M. Über die Verständigung bei indischen Bienen. Z. Vergl. Physiol. 1956, 38, 521–557. [Google Scholar] [CrossRef]
- Sakagami, S.F. Preliminary report on the specific difference behavior and the other ecological characters between European and Japanese honeybee. Acta Hymenopterol. 1960, 1, 171–198. [Google Scholar]
- Koeniger, N.; Fuchs, S. Kommunikative Schallerzeugung von Apis cerana Fabr. im Bienenvolk. Naturwissenschaften 1972, 59, 169. [Google Scholar] [CrossRef]
- Fuchs, S.; Koeniger, N. Schallerzeugung im Dienst der Verteidigung des Bienenvolkes (Apis cerana Fabr.). Apidologie 1974, 5, 271–287. [Google Scholar] [CrossRef]
- Seeley, T.D.; Seeley, R.H.; Akratanakul, P. Colony defence strategies of the honeybees in Thailand. Ecol. Monog. 1982, 52, 43–63. [Google Scholar] [CrossRef]
- Spangler, H.G. High-frequency sound production by honeybees. J. Apic. Res. 1986, 25, 213–214. [Google Scholar] [CrossRef]
- Sen Sarma, M.S.; Fuchs, S.; Werber, C.; Tautz, J. Worker piping triggers hissing for coordinated colony defence in the dwarf honeybee Apis Florea. Zoology 2002, 105, 215–223. [Google Scholar] [CrossRef]
- Kastberger, G.; Weihmann, F.; Hoetzl, T. Social waves in giant honeybees (Apis dorsata) elicit nest vibrations. Naturwissenschaften 2013, 100, 595–609. [Google Scholar] [CrossRef]
- Kawakita., S.; Ichikawa, K.; Sakamoto, F.; Moriya, K. Hissing of A. cerana japonica is not only a direct aposematic response but also a frequent behavior during daytime. Insect Soc. 2018, 65, 331–337. [Google Scholar] [CrossRef]
- Kawakita, S.; Ichikawa, K.; Sakamoto, F.; Moriya, K. Sound recordings of Apis cerana japonica colonies over 24 h reveal unique daily hissing patterns. Apidologie 2019, 50, 204–214. [Google Scholar] [CrossRef]
- Kirchner, W.H.; Hager, F.A.; Krausa, K. Vibrational behavior in honeybees. In Biotremology: Physiology, Ecology, and Evolution, 1st ed.; Hill, P.S.M., Mazzoni, V., Stritih Peljhan, N., Virant-Doberlet, M., Wessel, A., Eds.; Springer: Berlin/Heidelberg, Germany. (in press)
- Cocroft, R.B.; Gogala, M.; Hill, P.S.M.; Wessel, A. Fostering research progress in a rapidly growing field. In Studying Vibrational Communication, 1st ed.; Cocroft, R.B., Gogala, M., Hill, P.S.M., Wessel, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 3–12. [Google Scholar]
- Hill, P.S.M.; Wessel, A. Biotremology. Curr. Biol. 2016, 26, R187–R191. [Google Scholar] [CrossRef]
- Johnson, L.K.; Haynes, L.W.; Carlson, M.A.; Fortnum, H.A.; Gorgas, D.L. Alarm substances of the stingless bee. Trigona silvestriana. J. Chem. Ecol. 1985, 11, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Nunes, T.M.; von Zuben, L.G.; Costa, L.; Venturieri, G.C. Defensive repertoire of the stingless bee Melipona flavolineata Friese (Hymenoptera: Apidae). Sociobiol 2014, 61, 541–546. [Google Scholar] [CrossRef]
- Esch, H.; Esch, I.; Kerr, W.E. Sound: An element common to communication of stingless bees and to dances of the honey bee. Science 1965, 149, 320–321. [Google Scholar] [CrossRef] [PubMed]
- Krausa, K.; Hager, F.A.; Kiatoko, N.; Kirchner, W.H. Vibrational signals of African stingless bees. Insect Soc. 2017, 64, 415–424. [Google Scholar] [CrossRef]
- Küper, W.; Sommer, J.H.; Lovett, J.C.; Mutke, J.; Linder, H.P.; Beentje, H.J.; Van Rompaey, R.; Chatelain, C.; Sosef, M.S.M.; Barthlott, W. Africa’s hotspots of biodiversity redefined. Ann. Mo. Bot. Gard. 2004, 91, 525–535. [Google Scholar]
- Eardley, C.D. Taxonomic revision of the African stingless bees (Apoidea: Apidae: Apinae: Meliponini). Afr. Plant Prot. 2004, 10, 63–96. [Google Scholar]
- Wittmann, D. Aerial defense of the nest by workers of the stingless bee Trigona (Tetragonisca) angustula (Latreille) (Hymenoptera: Apidae). Behav. Ecol. Sociobiol. 1985, 16, 111–114. [Google Scholar] [CrossRef]
- Grüter, C.; Kärcher, M.H.; Ratnieks, F.L.W. The natural history of nest defence in a stingless bee, Tetragonisca angustula (Latreille) (Hymenoptera: Apidae), with two distinct types of entrance guards. Neotrop. Entomol. 2011, 40, 55–61. [Google Scholar] [CrossRef]
- Harano, K.; Maya Silva, C.; Hrncir, M. Why do stingless bees (Melipona subnitida) leave their nest with resin loads? Insect Soc. 2020, 67, 195–200. [Google Scholar] [CrossRef]
- Kirchner, W.H.; Friebe, R. Nestmate discrimination in the African stingless bee Hypotrigona gribodoi Magretti (Hymenoptera: Apidae). Apidologie 1999, 30, 293–298. [Google Scholar] [CrossRef]
- Roubik, D.W. Ecology and Natural History of Tropical Bees; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Breed, M.D.; Moure, J. Animal Behavior; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Breed, M.D.; Page, R.E. Intra- and interspecific nestmate recognition in Melipona workers (Hymenoptera: Apidae). J. Insect. Behav. 1991, 4, 463–469. [Google Scholar] [CrossRef]
- Suka, T.; Inoue, T. Nestmate recognition of the stingless bee Trigona (Tetragonula) minangkabau (Apidae: Meliponinae). J. Ethol. 1993, 11, 141–147. [Google Scholar] [CrossRef]
- Bowden, R.M.; Garry, M.F.; Breed, M.D. Discrimination of con- and heterospecific bees by Trigona (Tetragonisca) angustula guards. J. Kans. Entomol. Soc. 1994, 67, 137–139. [Google Scholar]
- Inoue, T.; Roubik, D.W.; Sakagami, S.F. Nestmate recognition in the stingless bee Melipona panamica (Apidae, Meliponini). Insect Soc. 1999, 46, 208–218. [Google Scholar] [CrossRef]
- Buchwald, R.; Breed, M.D. Nestmate recognition cues in a stingless bee, Trigona fulviventris. Anim. Behav. 2005, 70, 1331–1337. [Google Scholar] [CrossRef]
- Nunes, T.M.; Nascimento, F.S.; Turatti, I.C.; Lopes, N.P.; Zucchi, R. Nestmate recognition in a stingless bee: Does the similarity of chemical cues determine guard acceptance? Anim. Behav. 2008, 75, 1165–1171. [Google Scholar] [CrossRef]
- Jones, S.M.; van Zweden, J.S.; Grüter, C.; Menezes, C.; Alves, D.A.; Nunes-Silva, P.; Czaczkes, T.; Imperatriz-Fonseca, V.L.; Ratnieks, F.L.W. The role of wax and resin in the nestmate recognition system of a stingless bee, Tetragonisca angustula. Behav. Ecol. Sociobiol. 2012, 66, 1–12. [Google Scholar] [CrossRef]
- Krausa, K. Populationsbiologische Untersuchungen an den afrikanischen Stachellosen Bienen Hypotrigona gribodoi und Liotrigona spec. Master’s Thesis, Ruhr-University Bochum, Bochum, Germany, 2012. [Google Scholar]
- Maynard Smith, J.; Harper, D.G.C. Animal Signals; University Press: Oxford, UK, 2003. [Google Scholar]
- Scott-Phillips, T.C. Defining biological communication. J. Evol. Biol. 2008, 21, 387–395. [Google Scholar] [CrossRef]
- Grüter, C. Recruitment and communication in foraging. In Stingless Bees. Their behaviour, Ecology and Evolution, 1st ed.; Grüter, C., Ed.; Springer: Cham, Switzerland, 2020; pp. 341–372. [Google Scholar]
- Ratnieks, F.L.S.; Anderson, C. Task partitioning in insect societies. Insect Soc. 1999, 46, 95–108. [Google Scholar] [CrossRef]
- Hager, F.A.; Kirchner, W.H. Vibrational long-distance communication in the termites Macrotermes natalensis and Odontotermes sp. J. Exp. Biol. 2013, 216, 3249–3256. [Google Scholar] [CrossRef]
- Hunt, J.H.; Richard, F.J. Intracolony vibroacoustic communication in social insects. Insect Soc. 2013, 60, 403–417. [Google Scholar] [CrossRef]




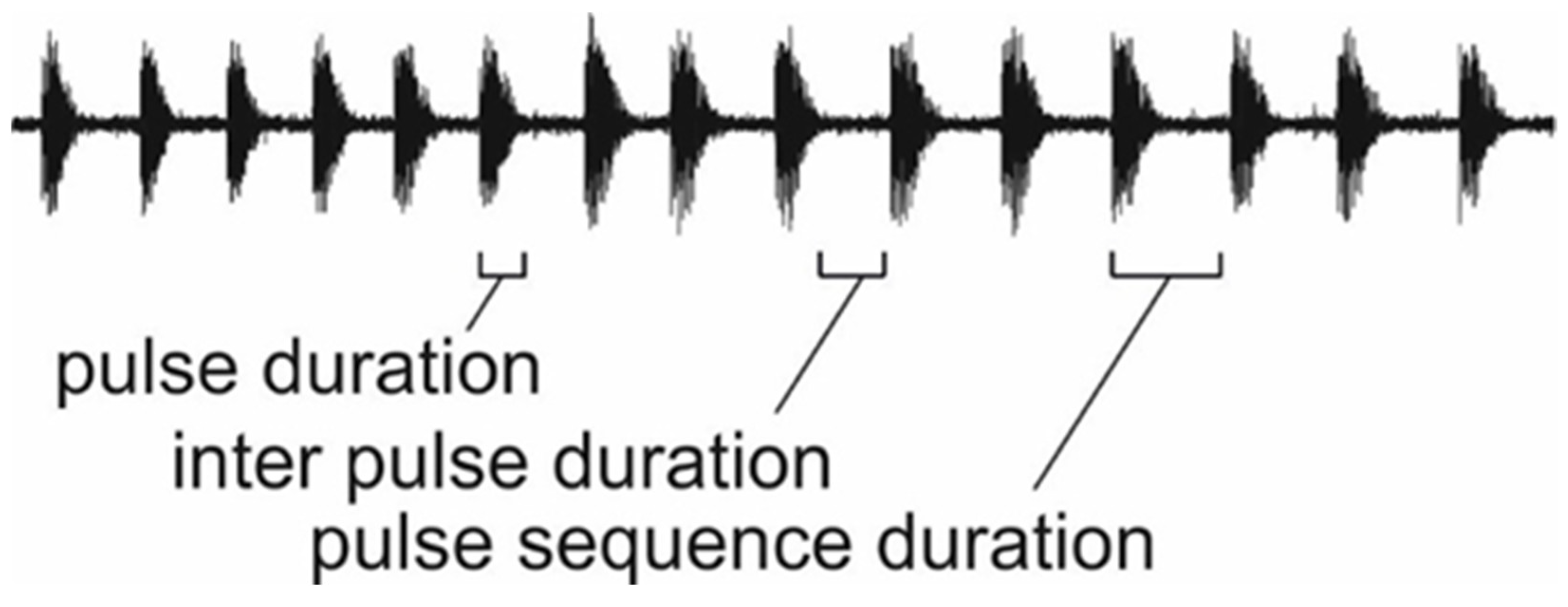{kind=link}
{kind=link}
{kind=link}
| Recording Location | PSD [ms] | PD [ms] | IPD [ms] | DC | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| m | sd | n | m | sd | n | m | sd | n | m | sd | n | N | |
| nest | 214 | ±38 | 45 | 63 | ±8 | 51 | 149 | ±42 | 45 | 0.32 | ±0.09 | 45 | 7 |
| entrance tube | 766 | ±275 | 67 | 182 | ±45 | 76 | 582 | ±278 | 67 | 0.34 | ±0.11 | 67 | 9 |
| arena | 683 | ±287 | 398 | 267 | ±116 | 406 | 416 | ±307 | 398 | 0.47 | ±0.15 | 398 | 8 |
| Recording Location | PSD | PD | IPD | DC | n | ||||
|---|---|---|---|---|---|---|---|---|---|
| CV | p | CV | p | CV | p | CV | p | ||
| (1) nest | 26.3 a | (1–2) 0.009 | 23.6 a | n.s. | 35.0 a | (1–2) 0.022 | 28.2 a | (1–2) 0.011 | 7 |
| (2) entrance tube | 67.4 b | (2–3) n.s. | 38.3 a | 96.3 b | (2–3) n.s. | 54.4 b | (2–3) n.s. | 9 | |
| (3) arena | 59.9 b | (3–1) 0.017 | 61.8 a | 90.9 b | (3–1) 0.009 | 39.6 a,b | (3–1) n.s. | 8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krausa, K.; Hager, F.A.; Kirchner, W.H. Guarding Vibrations—Axestotrigona ferruginea Produces Vibrations When Encountering Non-Nestmates. Insects 2021, 12, 395. https://doi.org/10.3390/insects12050395
Krausa K, Hager FA, Kirchner WH. Guarding Vibrations—Axestotrigona ferruginea Produces Vibrations When Encountering Non-Nestmates. Insects. 2021; 12(5):395. https://doi.org/10.3390/insects12050395
Chicago/Turabian StyleKrausa, Kathrin, Felix A. Hager, and Wolfgang H. Kirchner. 2021. "Guarding Vibrations—Axestotrigona ferruginea Produces Vibrations When Encountering Non-Nestmates" Insects 12, no. 5: 395. https://doi.org/10.3390/insects12050395
APA StyleKrausa, K., Hager, F. A., & Kirchner, W. H. (2021). Guarding Vibrations—Axestotrigona ferruginea Produces Vibrations When Encountering Non-Nestmates. Insects, 12(5), 395. https://doi.org/10.3390/insects12050395