
Contributions to the Taxonomy of Arlesminthurus Bretfeld and Calvatomina Yosii (Collembola, Symphypleona, Appendiciphora), with the Description of New Species from Northeastern Brazil †

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Taxonomic Summary of Arlesminthurus and Genus Diagnosis
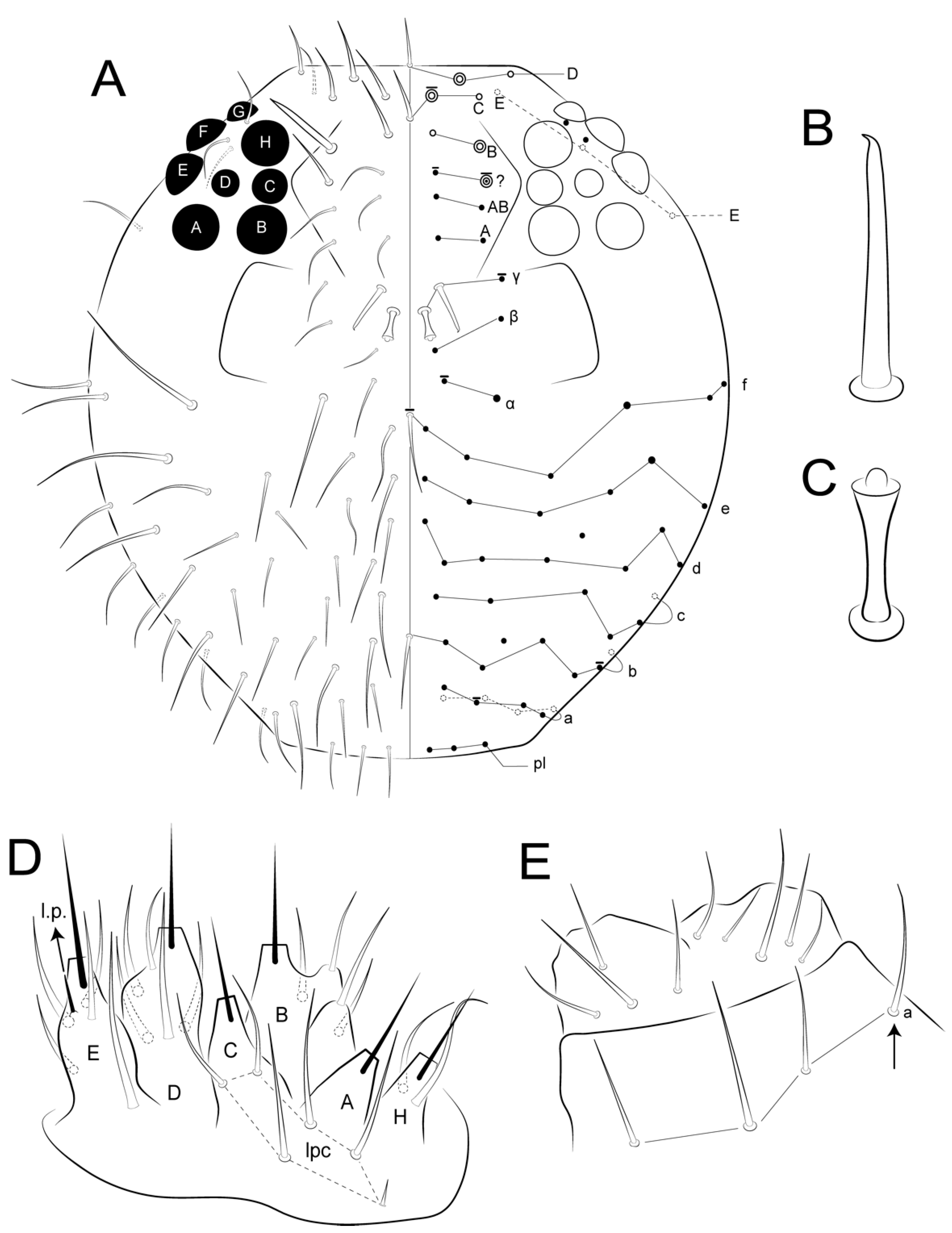
3.2. Arlesminthurus caatinguensis sp. nov. Cipola, Lycarião and Medeiros
3.3. Identification Key and Distribution of Arlesminthurus Species
- Color patterns sexually dimorphic; dens inner row with six chaetae slightly larger than the others … 2
- -
- 2.
- Male head with one postocullar and one lateral band of pigment, females with two small postocular spots; large abdomen with an evident W-shaped spot in males; Ant IV with 9–11 subsegments; male γ frontal chaetae apically fringed; unguis III pseudonychia present; mucro normal with smooth edges … A. franzkafkai Palacios-Vargas and Cabrera, 2015 [13] (Nicaragua)
- -
- Males with two longitudinal bands on lateral body, female head with one interantennal spot and with the large abdomen with three depigmented spots; Ant IV with eight subsegments; male γ frontal chaetae apically swollen; unguis III pseudonychia absent; mucro wide with irregular edges … A. richardsi (Arlé, 1971) [12] (Brazil)
- 3.
- Head color diffuse, with occipital region, as well as the body, depigmented; male head with inferior frontal chaetae (γ) apically hooked; mucro outer edge with four invaginations … A. salinensis (Arlé, 1971) [12] (Brazil)
- -
- 4.
- Ant IV with 6–7 subsegments; male with superior frontal chaetae (γ) apically swollen and f line unpaired chaeta larger than the others; unguis inner tooth absent; mucro inner edge irregular … A. aueti (Arlé, 1961) [11] (Brazil)
- -
- Ant IV with eight subsegments (Figure 7A); male with superior frontal chaetae (γ) apically hooked and f line unpaired chaeta subequal to the others (Figure 5A and Figure 8A,B); unguis inner tooth present (Figure 16B); mucro inner edge with two invaginations (Figure 17C) … A. caatinguensis sp. nov. (Brazil)
3.4. Taxonomic Summary of Calvatomina and Genus Diagnosis
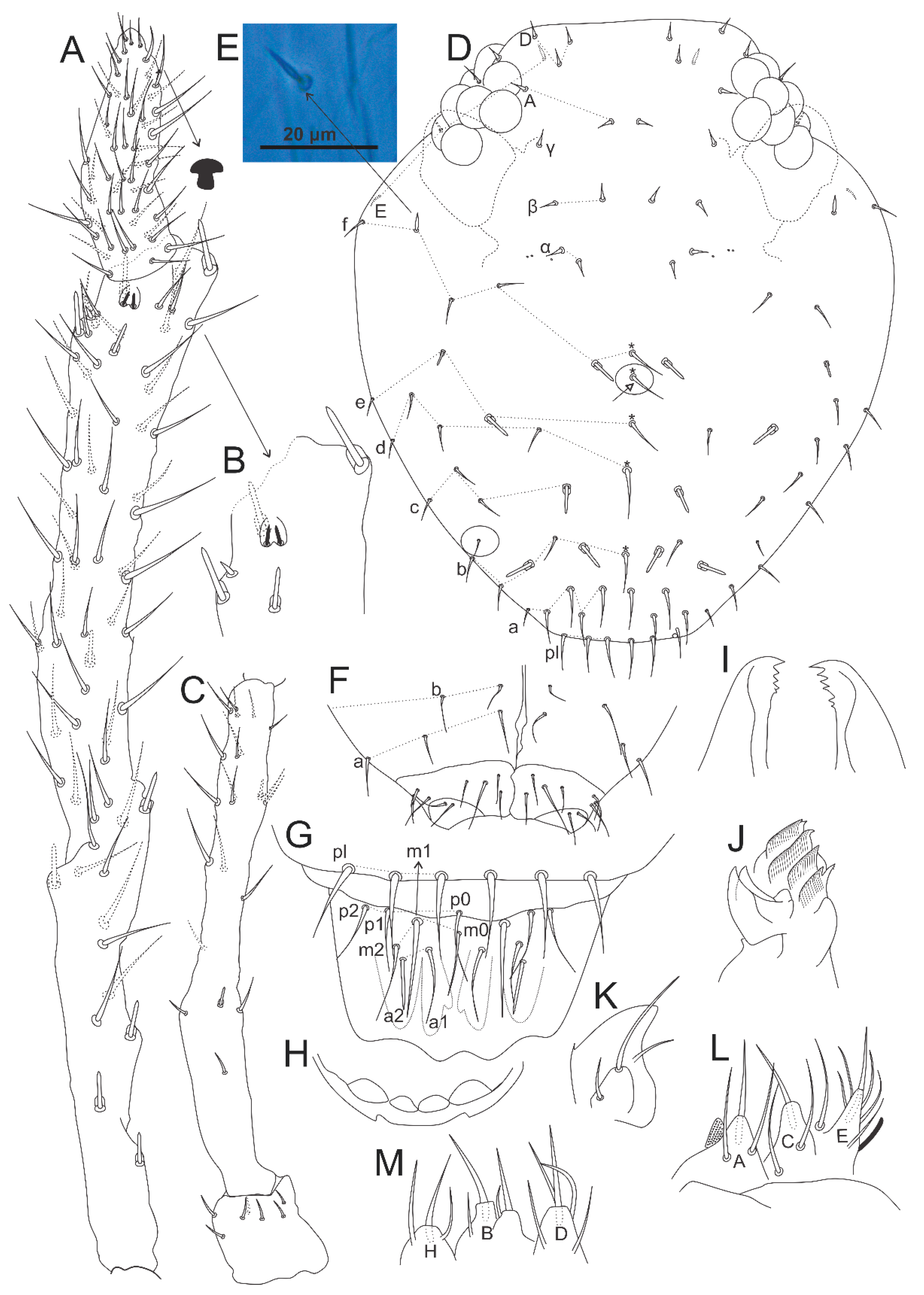
3.5. Calvatomina gladiata sp. nov. Medeiros, Bellini and Cipola
3.6. Identification Key and Distribution* of Neotropical Species of Calvatomina
- Dens ventral chaetae formula from the apex to the basis as 3–4,2,1,1,1,0,1, with four unpaired basal chaetae (bougainvilleae-group sensu Yosii, 1969) [38] … 2
- -
- Dens ventral chaetae formula from the apex to the basis as 3–4,2,1,1,0,0,1, with three unpaired basal chaetae … 3
- 2.
- Clypeal region lacking the lateral acanthoid chaeta on f line, dens dorsally with 23 chaetae, small abdomen of the female with as1(M), ms4(H) and mi1(L) as blunt acanthoid chaetae … C. christianseni (Delamare-Deboutteville and Massoud, 1964) [15] (Suriname)
- -
- 3.
- Small abdomen of the female with ms1(a0) as a regular pointed chaeta (formosana-group sensu Yosii, 1969) [37], chaetae ms2(N), ms4(H) and mi1(L) of the same region and gender also as regular pointed chaetae, dorsal color pattern of the large abdomen with two dark knife-shaped forms … C. nymphascopula Soto-Adames, 1988 [17] (Puerto Rico)
- -
- Small abdomen of the female with ms1(a0) as a blunt acanthoid chaeta (rufescens-group sensu Yosii, 1969) [37], chaetae ms2(N), ms4(H) and mi1(L) of the same region and gender also as blunt acanthoid chaetae, dorsal color pattern of the large abdomen otherwise … 4
- 4.
- Clypeal region with eight cup sensilla, collophore with one chaeta, tenaculum with one chaeta, manubrium with 10 chaetae … C. rufescens (Reuter, 1890) [40] (USA, Finland, Colombia, Cuba, Mexico, Puerto Rico)
- -
- Clypeus with 5–6 cup sensilla, collophore with two chaetae, tenaculum with two chaetae, manubrium with nine chaetae … 5
- 5.
- Ant III with nine and Ant II with three cup sensilla, respectively, clypeal region with five cup sensilla, dens dorsally with 24 chaetae, small abdomen of the female with as1(M) and as2(M’) as blunt acanthoid chaetae … C. guyanensis Nayrolles and Betsch, 1995 [18] (French Guiana)
- -
- Ant III with 10 and Ant II with two cup sensilla, respectively, clypeal region with six cup sensilla, dens dorsally with 25 chaetae, small abdomen of the female with as1(M) and as2(M’) as regular pointed chaetae … C. gladiata sp. nov. (Brazil)
4. Discussion
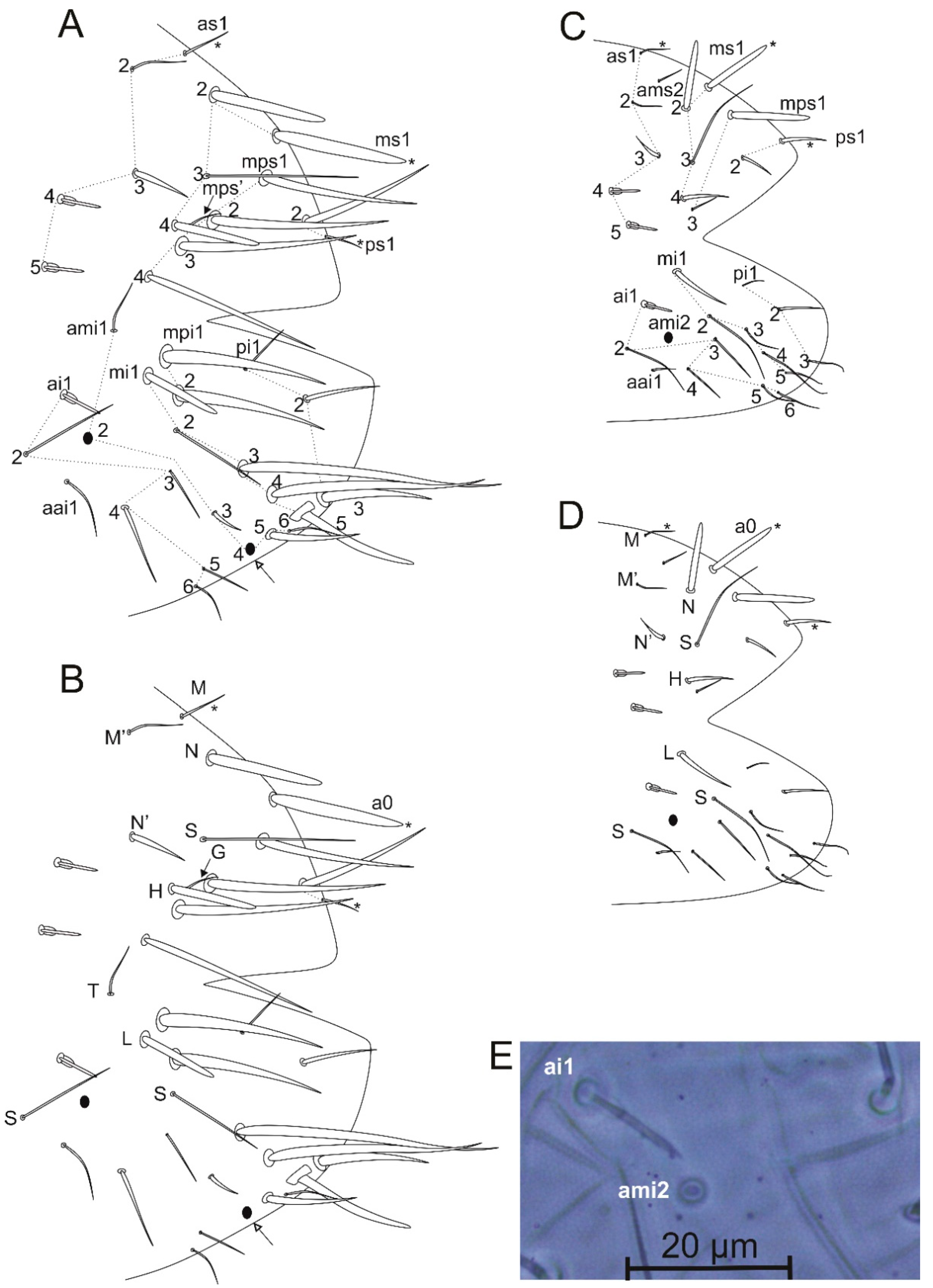
4.1. On the Bothriotrichum-Like Sens of Arlesminthurus and the Genus Affinities
4.2. On the Differential Chaetotaxy of Calvatomina Species
4.3. On the Validity of Calvatomina Internal Groups
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bretfeld, G. Sturmius epiphytus n. gen. n. spec. from Colombia, a taxon of the Symphypleona (Insecta, Colembola) with an unexpected character combination. Description and position in non-Linnean and Linnean classifications of the Symphypleona. Zool. Syst. Evol. Res. 1994, 32, 264–281. [Google Scholar] [CrossRef]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 25 February 2021).
- Bretfeld, G. Phylogenetic systematics of the higher taxa of Symphypleona Borner, 1901 (Insecta, Entognatha, Collembola). Proc. 2nd Intern. Sem. Apterygota 1986, 1, 302–311. [Google Scholar]
- Bretfeld, G. Synopses on Palaeartic Collembola, Volume 2. Symphypleona. Abh. Ber. Naturkundemus. Gorlitz 1999, 71, 1–318. [Google Scholar]
- Zeppelini, D.; Queiroz, G.C.; Bellini, B.C. Symphypleona in Catálogo Taxonômico da Fauna do Brasil. PNUD. Available online: http://fauna.jbrj.gov.br/fauna/faunadobrasil/470 (accessed on 1 February 2021).
- Bellini, B.C. Fauna de Collembola (Arthropoda) em áreas úmidas do semiárido. In Artrópodes do Semiárido, Biodiversidade e Conservação, 1st ed.; Bravo, F., Calor, A., Eds.; Feira de Santana Printmídia: Feira de Santana, Brazil, 2014; Volume 1, pp. 57–68. [Google Scholar]
- Bellini, B.C.; Godeiro, N.N. Novos registros de Collembola (Arthropoda, Hexapoda) para áreas úmidas do semiárido do Brasil. In Artrópodes do Semiárido II: Biodiversidade e Conservação, 1st ed.; Bravo, F., Ed.; Métis Produção Editorial: São Paulo, Brazil, 2017; Volume 1, pp. 28–53. [Google Scholar]
- Yosii, R. On some Collembola of Afghanistan, India and Ceylon, collected by the Kuphe-Expedition, 1960. R. Kyoto Univ. Sci. Exp. Karakoram Hindukush 1966, 8, 332–405. [Google Scholar]
- Santos-Rocha, I.M.; Andreazze, R.; Bellini, B.C. Registros de Collembola (Arthropoda, Hexapoda) no Estado do Rio Grande do Norte. Biota Neotrop. 2011, 11, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Cipola, N.G.; Zequi, J.A.C.; Bellini, B.C. Collembola (Hexapoda). In Fauna e flora do Parque Estadual Mata São Francisco—Norte do Paraná, 1st ed.; Zequi, J.A.C., Orsi, M.L., Shibatta, L.S., Eds.; Eduel: Londrina, Brazil, 2019; Volume 1, pp. 105–119. [Google Scholar]
- Arlé, R. Novas espécies de colêmbolas aquáticas (Nota preliminar). Atas Soc. Biol. Rio de Janeiro 1961, 5, 34–37. [Google Scholar]
- Arlé, R. Collemboles d´Amazonie, III. Quelques Symphypléones du Bas-Amazone et complément à la description de Deuterosminthurus aueti Arlé, 1961. Bol. Mus. Para. Emílio Goeldi 1971, 75, 1–11. [Google Scholar]
- Palacios-Vargas, J.G.; Cabrera, M. Arlesminthurus franzkafkai sp. nov. (Collembola; Borletiellidae) from Nicaragua. Brenesia 2015, 83–84, 62–67. [Google Scholar]
- Cipola, N.G.; Silva, D.D.; Bellini, B.C. Class Collembola. In Keys to Neotropical Hexapoda—Thorp and Cobish’s Freshwater Invertebrates, 4th ed.; Hamada, N., Thorp, J.H., Rogers, D.C., Eds.; Elsevier Academic Press: London, UK, 2018; Volume 3, pp. 11–55. [Google Scholar] [CrossRef]
- Delamare-Deboutteville, C.; Massoud, Z. Studies on the fauna of Suriname and other Guianas: No. 26. Collemboles Symphypléones du Surinam. Stud. Fauna Sur. Guy. 1964, 26, 56–81. [Google Scholar]
- Mari-Mutt, J.A. Collembola from two localities near Buenaventura, Colombia. J. Kans. Entomol. Soc. 1987, 60, 364–379. [Google Scholar]
- Soto-Adames, F.N. Nuevos dicirtómidos de Puerto Rico (Insecta: Collembola: Dycyrtomidae). Caribb. J. Sci. 1988, 14, 60–70. [Google Scholar]
- Nayrolles, P.; Betsch, J.M. Discussion on the genus Calvatomina Yosii, 1966 (Collembola, Symphypleona) with description of a new species, C. guyanenses, from French Guiana. Zool. Anz. 1995, 234, 281–292. [Google Scholar]
- Börner, C. Das System der Collembolen nebst Beschreibung neuer Collembolen des Hamburger Naturhistorischen Museums. Mit. Natur. Mus. Hambg. 1906, 23, 147–188. [Google Scholar]
- Shorthouse, D. SimpleMappr, an Online Tool to Produce Publication-Quality Point Maps. Available online: http://www.simplemappr.net (accessed on 5 September 2020).
- Nayrolles, P. La chetotaxie antennaire des Collemboles Symphypleones. Trav. Lab. Ecobiol. Arthr. Édaph. Toulouse 1991, 6, 1–94. [Google Scholar]
- Nayrolles, P. Aspects structuraux de la chétotaxie appendiculaire des Collemboles Symphypléones. Nouv. Rev. Entomol. 1992, 9, 345–356. [Google Scholar]
- Nayrolles, P. La biométrie des caractères discontinus d’après le revêtement appendiculaire des Collemboles Symphypléones. I. Sur quelques concepts nouveaux dans l’analyse numérique de la chétotaxia. Bull. Mus. Natl. Hist. Nat. ParisSérie 4 1993, 15, 79–93. [Google Scholar]
- Cipola, N.G.; Morais, J.W.; Bellini, B.C. A new species of Seira (Collembola: Entomobryidae: Seirini) from Northern Brazil, with the addition of new chaetotaxic characters. Zoologia 2014, 31, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Fjellberg, A. The Labial Palp in Collembola. Zool. Anz. 1999, 237, 309–330. [Google Scholar]
- Nayrolles, P. Chetotaxie tibiotarsale des collemboles symphyplèones. Trav. Lab. Ecobiol. Arthr. Édaph. Toulouse 1988, 5, 1–19. [Google Scholar]
- Hüther, W. New aspects in taxonomy of Lepidocyrtus (Collembola). Proc. 2nd Intern. Sem. Apterygota 1986, 1, 61–65. [Google Scholar]
- Nayrolles, P. Données nouvelles sur l’évolution ontogénétique des Collemboles Symphypléones. Nouv. Rev. Entomol. 1989, 6, 231–244. [Google Scholar]
- Nayrolles, P. Chetotaxie furcale des Collemboles Symphypleones. Trav. Lab. Ecobiol. Arthr. Édaph. Toulouse 1990, 6, 27–50. [Google Scholar]
- Betsch, J.M.; Waller, A. Chaetotaxic nomenclature of the head, thorax and abdomen in Symphypleona (Insecta, Collembola). Acta Zool. Fenn. 1994, 195, 5–12. [Google Scholar]
- Betsch, J.M. An ontogenetically focused chaetotaxial scheme in Symphypleona (collembolan): The 6th abdominal segment. Pedobiologia 1997, 41, 13–18. [Google Scholar]
- Börner, C. Neue Collembolenformen und zur Nomenclatur der Collembola Lubbock. Zool. Anz. 1901, 24, 696–712. [Google Scholar]
- Börner, C. Die Familien der Collembolen. Zool. Anz. 1913, 61, 315–322. [Google Scholar]
- Betsch, J.M. Contribution a l’etude de la reproduction chex les Bourletiellinae (Collemboles, Symphypleones). Pedobiologia 1974, 14, 179–181. [Google Scholar]
- Richards, W.R. Generic classification, evolution, and biogeography of the Sminthuridae of the world (Collembola). Mem. Ent. Soc. Can. 1968, 53, 1–54. [Google Scholar] [CrossRef]
- Betsch, J.M. Éléments pour une monographie des Collemboles Symplyplêones (Hexapodes, Aptérygotes). Mém. Mus. Natl. Hist. Nat. Sér. A Zool. 1980, 116, 1–227. [Google Scholar]
- Yosii, R. Dicyrtomina and Ptenothrix (Insecta: Collembola) of the Solomon Islands. Zool. J. Linn. Soc. 1969, 48, 217–236. [Google Scholar] [CrossRef]
- Prabhoo, N.R. Bark and moss inhabiting Collembola of South India. Bull. Entomol. 1971, 12, 41–47. [Google Scholar]
- Yosii, R. On some Collembola of New Caledonia, New Britain and Solomon Islands. Bull. Osaka Mus. Nat. Hist. 1960, 12, 9–38. [Google Scholar]
- Reuter, O.M. Collembola in caldariis viventia enumeravit novasque species descripsit. Med. Soc. Fauna Flora Fenn. 1890, 17, 17–28. [Google Scholar]
- Hüther, W. Wiederbeschreibung von Dicyrtomina rufescens (Reuter 1890); nebst einigen allgemeinen taxonomischen Bemerkungen (Collembola). Not. Entomol. 1965, 45, 47–52. [Google Scholar]
- Schött, H. Études sur les Collemboles du Nord. K. Sven. Vetensk. Akad. Handl. Stockh. 1902, 28, 2–48. [Google Scholar]
- Fjellberg, A. Fauna Entomologica Scandinavica 42—The Collembola of Fennoscandia and Denmark. Part II: Entomobryomorpha and Symphypleona, 1st ed.; Koninklijke Brill NV: Leiden, The Netherlands, 2007; pp. 1–264. [Google Scholar]
- Bellini, B.C.; Santos, N.M.C.; Souza, P.G.C.; Weiner, W.M. Two new species of Brazilian springtails (Hexapoda: Collembola) with notes on Neotropical Brachystomella Ågren and Seira (Lepidocyrtinus) Börner. Insect Syst. Evol. 2018, 50, 297–326. [Google Scholar] [CrossRef]
- Mari-Mutt, J.A.; Bellinger, P.F. A Catalog of Neotropical Collembola, Including Nearctic Areas of Mexico—Flora & Fauna Handbook 5, 1st ed.; Sandhill Crane Press: Gainesville, FL, USA, 1990; pp. 1–237. [Google Scholar]
- Mari-Mutt, J.A.; Bellinger, P.F. Supplement to the catalog of the Neotropical Collembola. Caribb. J. Sci. 1996, 32, 166–175. [Google Scholar]
- Mari-Mutt, J.A.; Bellinger, P.F.; Janssens, F. Checklist of the Collembola: Supplement to the Catalog of the Neotropical Collembola. Available online: http://www.collembola.org/publicat/neotrcat.htm (accessed on 28 January 2021).
- Betsch, J.-M. Etude des Collemboles de Madagascar. II. Principaux cadres génériques des Symphypléones de l’étage montagnard. Bull. Mus. Natl. Hist. Nat. 3 1974, 147, 529–569. [Google Scholar]
- Bretfeld, G. Chaetotaxy of four species of the genera Heterosminthurus, Bourletiella, Deuterosminthurus and Prorastriopes (Insecta, Collembola, Symphypleona). Zool. Jb. Syst. 1990, 117, 441–489. [Google Scholar]
- Zeppelini, D.; Silva, D.D. A new Stenognathriopes (Collembola, Symphypleona, Bourletiellidae) from Brazilian coast. Zootaxa 2012, 3540, 51–58. [Google Scholar] [CrossRef]
- Betsch, J.M.; Lasebikan, B.A. Collembola du Nigéria, I. Stenognathriopes, un nouveau genre de Symphypléones. Bull. Soc. Entomol. Fr. 1979, 84, 165–170. [Google Scholar]
- Stach, J. Vorarbeiten zur Apterygoten-Fauna Polens Teil II: Apterygoten aus den Pieniny. Bull. Int. Acad. Polon. Sci. Cl. Sci. Math. Ser. B Sci. Nat. 1920, 2, 133–233. [Google Scholar]
- Banks, N. The Smynthuridae of Long Island, New York. J. N. Y. Entomol. Soc. 1899, 6, 193–197. [Google Scholar]
- Gisin, H. Collembolenfauna Europas. M. Hist. Nat. Geneve 1960, 50, 1–312. [Google Scholar]
- Stach, J. A new genus Andiella n.g. from the Andes, and revision of the genera of the tribe Bourletiellini Börn. (Collembola). Ann. Zool. 1955, 16, 1–59. [Google Scholar]
- Baquero, E.; Moraza, M.L.; Jordana, R. A new species of Polydiscia (Acari, Prostigmata, Tanaupodidae) with reference to its host: A new species of Deuterosminthurus (Collembola, Symphypleona, Bourletiellidae). Zootaxa 2003, 188, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Bretfeld, G. Description of old and new European taxa of the genus Fasciosminthurus Gisin, 1960 n. comb. (Collembola, Symphypleona) mainly based on chaetotaxy. Mit. Zool. Mus. Kiel 1992, 4 (Suppl. S4), 7–36. [Google Scholar]
- Bretfeld, G. Generic division of Prorastriopes Betsch, 1977 with description of Cyprania n.g. from Cyprus (Collembola, Symphypleona). Mit. Zool. Mus. Kiel 1992, 4 (Suppl. S4), 37–60. [Google Scholar]
- Delamare-Deboutteville, C. Collemboles Nouveaux du Sénégal—Contribution à la Connaissance des Bourletiellini C.B. Bull. Soc. Entomol. Fr. 1947, 52, 103–107. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Families: Neelidae and Dicyrtomidae, 1st ed.; Panstwowe Wydawnictwo Naukowe: Kraków, Poland, 1957; pp. 1–113. [Google Scholar]
- Medeiros, G.S.; Bellini, B.C. Two new species of Temeritas Richards (Collembola, Symphypleona, Sminthuridae) from Neotropical Region, with comments on the genus. Zootaxa 2019, 4586, 536–552. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, G.S.; Greenslade, P.; Bellini, B.C. The rare Richardsitas Betsch (Collembola, Symphypleona, Sminthuridae): A new species from Australia with comments on the genus and on the Sminthurinae. Insects 2020, 11, 519. [Google Scholar] [CrossRef] [PubMed]
- Snider, R.J. A contribution to the Dicyrtomidae (Collembola) of Hawaii. Zool. Scr. 1990, 19, 73–99. [Google Scholar] [CrossRef]
- Christiansen, K.A.; Bellinger, P. Insects of Hawaii: A Manual of the Insects of the Hawaiian Islands, Including an Enumeration of the Species and Notes on Their Origin, Distribution, Hosts, Parasites, etc. 15—Collembola, 1st ed.; University of Hawaii Press: Honolulu, HI, USA, 1992; pp. 1–445. [Google Scholar]
- Christiansen, K.A.; Bellinger, P. Biogeography of Hawaiian Collembola: The simple principles and complex reality. Orient. Insects 1994, 28, 309–351. [Google Scholar] [CrossRef]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | |||||||
|---|---|---|---|---|---|---|---|
| A. aueti [11,12] | A. caatinguensis | A. franzkafkai [13] | A. richardsi [12] | A. salinensis [12] | |||
| (Arlé, 1961) | sp. nov. | Palacios-Vargas and Cabrera, 2015 | (Arlé, 1971) | (Arlé, 1971) | |||
| Records: | PA, MT | PB and RN | Moropotente | PA | PA | ||
| Characteristics | Brazil | Brazil | Nicaragua | Brazil | Brazil | ||
| Sexual dimorphism in color pattern | – | – | + | + | – | ||
| Color pattern of males’ head | 1 median | 1 median | 1 postocular and | two lateral | diffuse, occipital head | ||
| longitudinal strip | longitudinal strip | 1 lateral bands | longitudinal band | depigmented | |||
| Body color pattern of males | 2 lateral spot and | 2 lateral spot and | W–shaped spot | two lateral | depigmented | ||
| 1 longitudinal strip | 1 longitudinal strip | strongly pigmented | longitudinal bands | ||||
| depigmented | depigmented | ||||||
| Color pattern of female’s head | as of the males | as of the males | 2 postocular spots | 1 interantennal spots | as of the males | ||
| Body color pattern of females | as of the males | as of the males | W–shaped spot weakly | 3 spots depigmented | as of the males | ||
| Legs and furcula pigmentation | – | + | – | – | – | ||
| Ant IV subsegments | 6–7 | 8 | 9–11 | 8 | 8 | ||
| Male’s γ frontal chaetae (inferior) | short and robust | short and robust | fringed apex | swollen apex | hooked apex | ||
| Male’s γ frontal chaetae (superior) | swollen apex | hooked apex | fringed apex | swollen apex | hooked apex | ||
| Male’s interocular spines | 3 | 0–4 | – | 0 * | 0 * | ||
| Male’s interocular spine-like chaetae | ? | 5–9 | 4 | 0 * | 0 * | ||
| Male’s unpaired chaetae (f lines) | larger | normal | normal | normal | normal | ||
| Large abdomen ventral chaetae | ? | 6–7 | 3 | ? | ? | ||
| Abd VI robust spine-like chaeta (?) | ? | + | – | – | – | ||
| Unguis inner tooth | – | + | – | vestigial | vestigial | ||
| Unguis III pseudonychia | ? | – | + | – | – | ||
| Male’s genital plate | ? | 3 + 4–5 | 3 + 7 * | ? | ? | ||
| Female’s genital plate | ? | 1 + 4–5 | 0 + 4 | ? | ? | ||
| Manubrium dorsal chaetae | ? | 9 | 6 | ? | ? | ||
| Dens chaetae (per line) | inner | 7 larger | 7 larger | 6 gently larger | 6 gently larger | 7 larger | |
| outer | 8 | 8 | 8–9 | 8 | 8 | ||
| dorsal | 8 | 8 | 8 | 8 | 8 | ||
| ventral formula | 1 *,3,1,1…0 | 3,3,1,1…1 | 3,3,1,1…1 | 3,3,1,1…1 | 3,3,1,1…1 | ||
| Mucro shape | wide | wide | thin | wide | wide | ||
| Mucro edges | inner | irregular | two invaginations | smooth | irregular | smooth | |
| outer | two invaginations | two invaginations | smooth | irregular | four invaginations | ||
| Species | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C. christianseni | C. cruciata | C. discolor ** | C. gladiata | C. guyanensis | C. nymphascopula | C. pallida | C. rufescens ** | C. trivandrana | ||
| References: | [15] | [8] | [16,42] | sp. nov. | [18] | [17] | [8] | [16,17,18,40,41] | [38] | |
| Type Locality: | Suriname | India | Sweden | Brazil | French Guiana | Puerto Rico | India | Finland | India | |
| Characteristics | Species-Group: | bougainvilleae | rufescens | bougainvilleae | rufescens | rufescens | formosana | rufescens | rufescens | rufescens |
| Head color pattern | irregular spots | 2 dorso-lateral spots | irregular spots | mostly pigmented | diffuse | irregular spots | 1 median spot | diffuse | irregular spots | |
| Large abdomen color pattern | dorsally | 2 dark knife-shaped forms | almost stripped + cross-shaped form | 2 dark knife-shaped forms | light sword-shape form | irregular spots | 2 dark knife-shaped forms | diffuse | light median stripe | median spots |
| laterally | J-shaped spot | all pigmented | J-shaped spot | all pigmented | narrow dark band | irregular spots | diffuse | large dark band | irregular spots | |
| Ant III cup sensilla | ? | ? | ? | 10 | 9 | 10 | ? | 10 | 8 | |
| Ant II cup sensilla | ? | ? | ? | 2 | 3 | ? | ? | 3 | 3 | |
| Clypeal f line acanthoid chaeta | – | ? | + | + | + | + | ? | +5 | ? | |
| Clypeal cup sensilla | 8 | ? | 8 | 6 | 5 | 5 | ? | 85 | ? | |
| Clypeal unpaired chaetae (superior) | 5 | 6 | 5 | 4–5 | 5 | 4–5 | ? | 55 | 5 | |
| Collophore chaetae | ? | ? | 1 | 2 | 2 | ? | ? | 1 | 1 | |
| Tenaculum chaetae | ? | ? | 1 | 2 | 2 | ? | ? | 1 | 2 | |
| Manubrial chaetae | ? | 9 | 10 | 9 | 9 | ? | ? | 10 | 9 | |
| Dens dorsal chaetae * | 23 | 24 | 24 | 25 | 24 | 21 | 23 | 21–24 | 24 | |
| Dens ventral formula | 4,2,1,1,1,0,1 | 3,2,1,1,0,0,1 | 3,2,1,1,1,0,1 | 4,2,1,1,0,0,1 | 3,2,1,1,0,0,1 | 4,2,1,1,0,0,1 | 3,2,1,1,0,0,1 | 3,2,1,1,0,0,1 | 2,2,1,1,0,0,1 | |
| Abdominal neosminthuroid chaetae | 5 | ? | 5 | 4–5 | 5 | 4 | ? | 5 | ? | |
| Small abdomen of the female | as1 (M) | BA | RC | RC | RC | BA | RC | RC | RC/BA | RC |
| as2 (M’) | ? | RC | RC | RC | BA | RC | RC | RC | RC | |
| as3 (N’) | ? | RC | RC | RC | RC | RC | RC | RC *** | RC | |
| ms1 (a0) | BA | BA | BA | BA | BA | RC | BA | BA | BA | |
| ms2 (N) | BA | BA | BA | BA | BA | RC | BA | BA | BA | |
| ms4 (H) | BA | BA | RC | BA | BA | RC | RC | BA | BA | |
| ami1 (T) | –? | RC | RC | RC | RC | RC | RC | RC | RC | |
| mi1 (L) | BA | BA | RC | BA | BA | RC | RC | BA | BA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cipola, N.G.; Medeiros, G.D.S.; Oliveira, V.A.M.D.; Barbosa, L.G.D.M.; Lycarião, T.A.; Bellini, B.C. Contributions to the Taxonomy of Arlesminthurus Bretfeld and Calvatomina Yosii (Collembola, Symphypleona, Appendiciphora), with the Description of New Species from Northeastern Brazil. Insects 2021, 12, 433. https://doi.org/10.3390/insects12050433
Cipola NG, Medeiros GDS, Oliveira VAMD, Barbosa LGDM, Lycarião TA, Bellini BC. Contributions to the Taxonomy of Arlesminthurus Bretfeld and Calvatomina Yosii (Collembola, Symphypleona, Appendiciphora), with the Description of New Species from Northeastern Brazil. Insects. 2021; 12(5):433. https://doi.org/10.3390/insects12050433
Chicago/Turabian StyleCipola, Nikolas G., Gleyce Da S. Medeiros, Viviane A. M. De Oliveira, Luís G. De M. Barbosa, Thainá A. Lycarião, and Bruno C. Bellini. 2021. "Contributions to the Taxonomy of Arlesminthurus Bretfeld and Calvatomina Yosii (Collembola, Symphypleona, Appendiciphora), with the Description of New Species from Northeastern Brazil" Insects 12, no. 5: 433. https://doi.org/10.3390/insects12050433