
Geographic Monitoring of Insecticide Resistance Mutations in Native and Invasive Populations of the Fall Armyworm

 ,
,  , , , ,
, , , , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
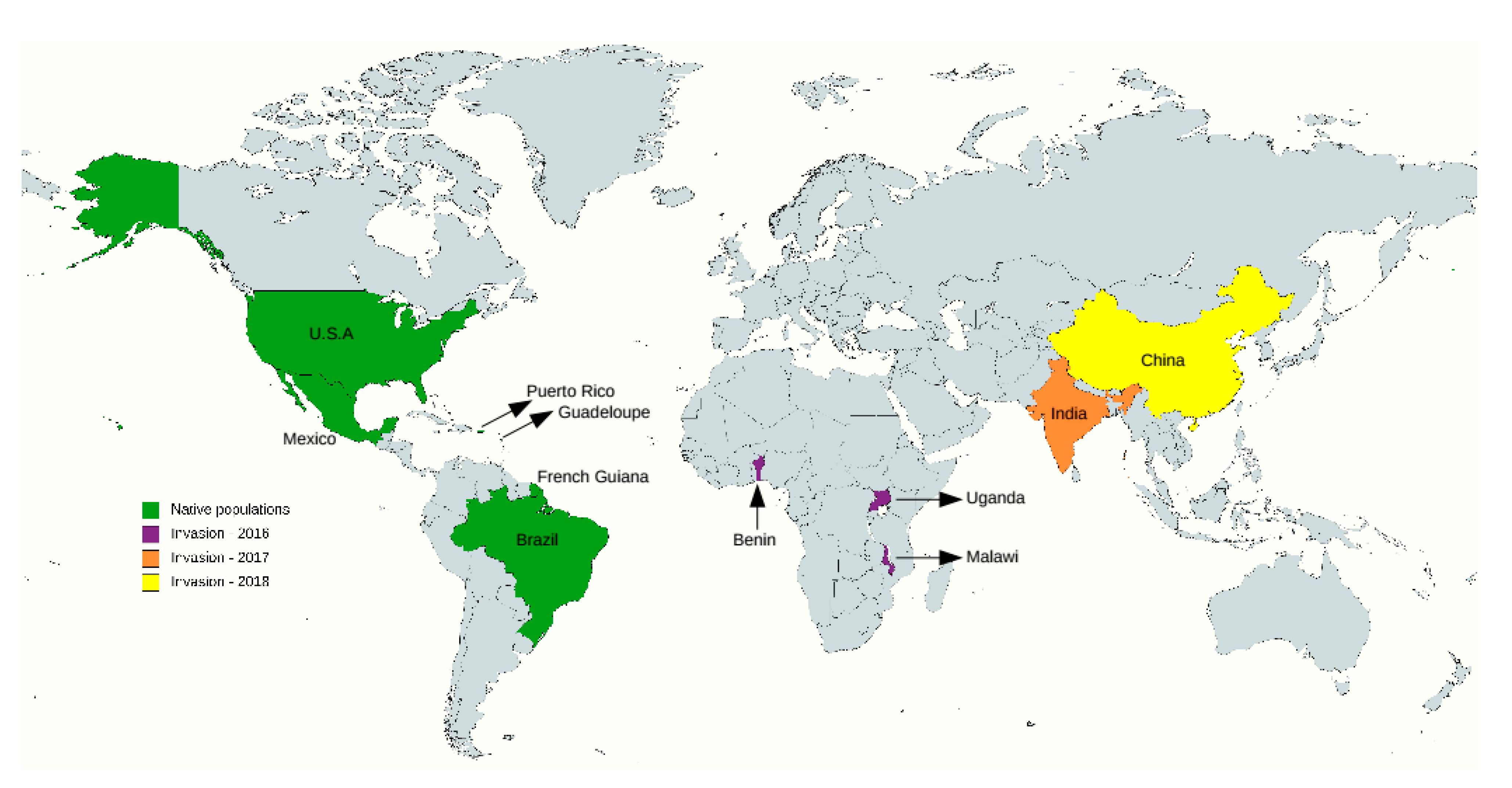
2.1. Resequencing Data
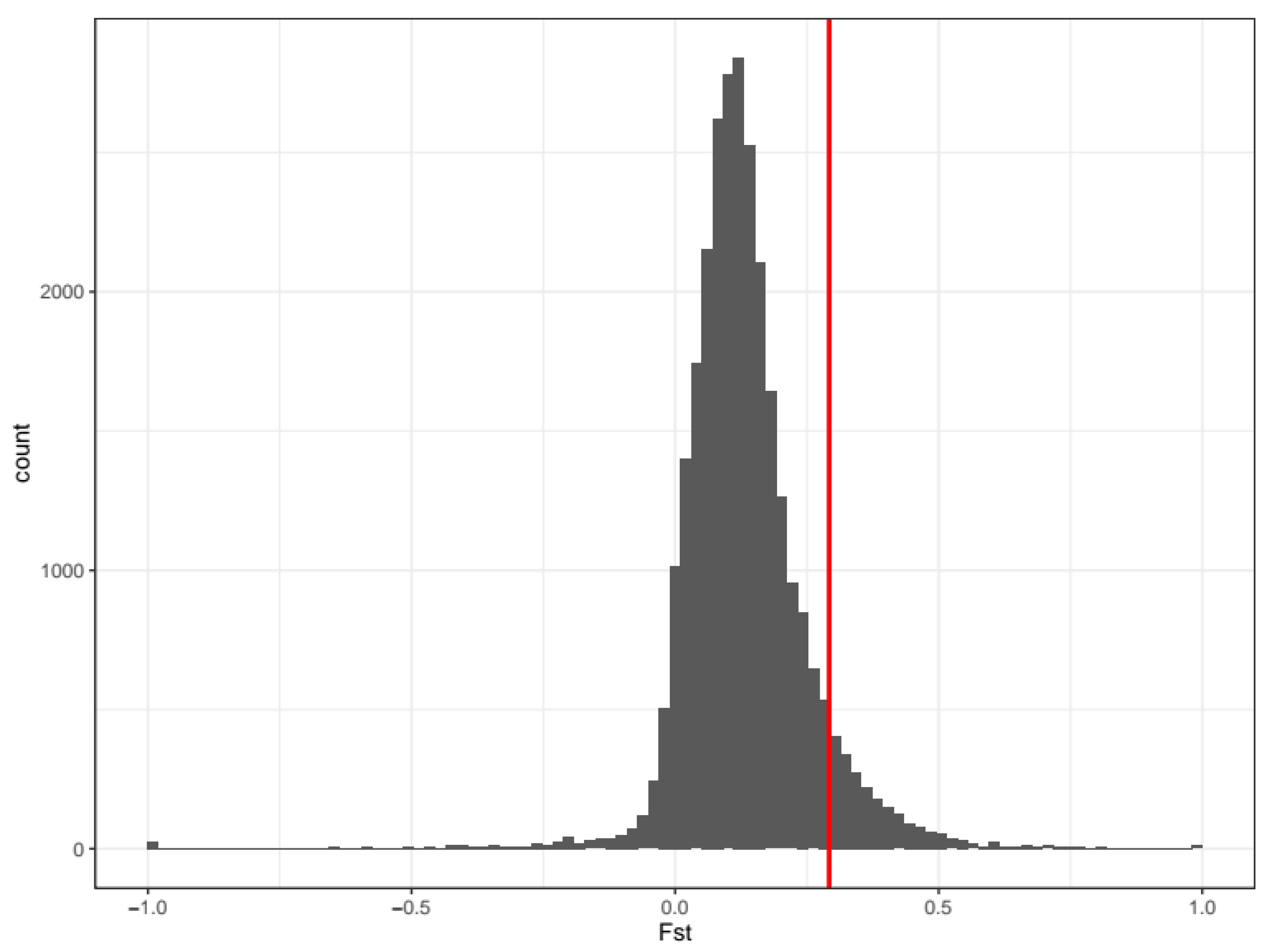
2.2. Gene Annotation and Statistical Analysis
3. Results
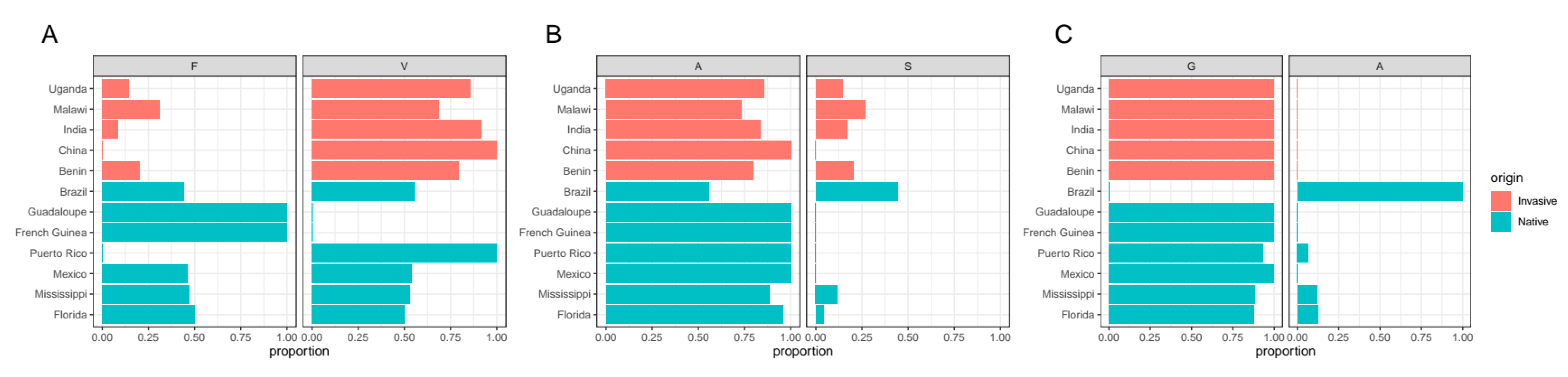
3.1. Bt Insecticide Resistance–ABCC2 Gene
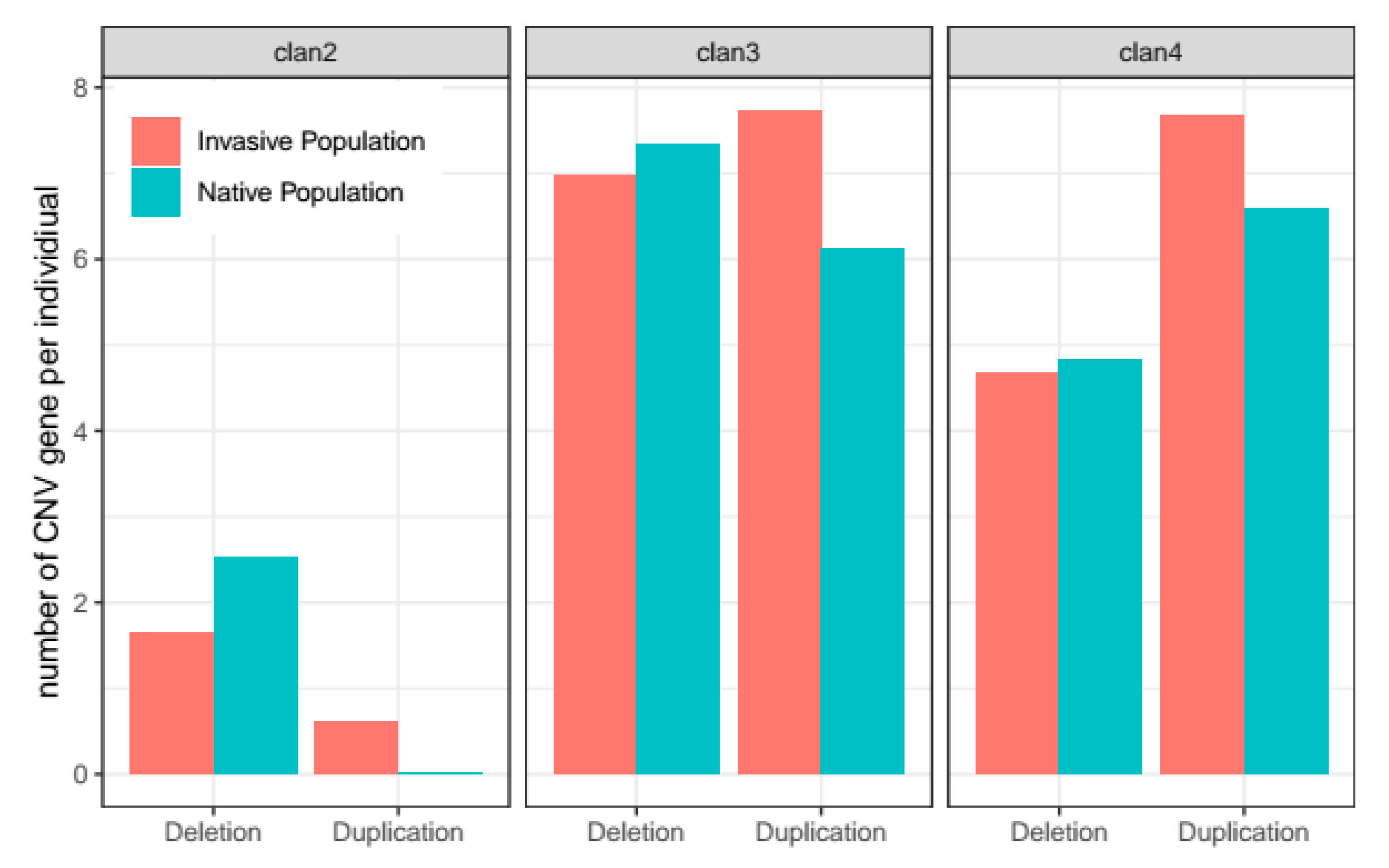
3.2. Synthetic Insecticide Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect Resistance to Bt Crops: Lessons from the First Billion Acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Heckel, D.G.; Ferré, J. Mechanisms of Resistance to Insecticidal Proteins from Bacillus thuringiensis. Annu. Rev. Entomol. 2021, 66, 121–140. [Google Scholar] [CrossRef]
- Hemingway, J.; Field, L.; Vontas, J. An Overview of Insecticide Resistance. Science 2002, 298, 96–97. [Google Scholar] [CrossRef]
- Carvalho, R.A.; Omoto, C.; Field, L.M.; Williamson, M.S.; Bass, C. Investigating the Molecular Mechanisms of Organophosphate and Pyrethroid Resistance in the Fall Armyworm Spodoptera frugiperda. PLoS ONE 2013, 8, e62268. [Google Scholar] [CrossRef]
- Shan, T.; Chen, C.; Ding, Q.; Chen, X.; Zhang, H.; Chen, A.; Shi, X.; Gao, X. Molecular Characterization and Expression Profiles of Nicotinic Acetylcholine Receptors in Bradysia odoriphaga. Pestic. Biochem. Physiol. 2020, 165, 104563. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.-Y.; Kwon, D.H.; Jeong, I.H.; Koh, Y.H. An Investigation of the Molecular and Biochemical Basis Underlying Chlorantraniliprole-Resistant Drosophila Strains and Their Cross-Resistance to Other Insecticides. Arch. Insect Biochem. Physiol. 2018, 99, e21514. [Google Scholar] [CrossRef]
- Lin, L.; Hao, Z.; Cao, P.; Yuchi, Z. Homology Modeling and Docking Study of Diamondback Moth Ryanodine Receptor Reveals the Mechanisms for Channel Activation, Insecticide Binding and Resistance. Pest Manag. Sci. 2020, 76, 1291–1303. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; de Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Gouin, A.; Bretaudeau, A.; Nam, K.; Gimenez, S.; Aury, J.-M.; Duvic, B.; Hilliou, F.; Durand, N.; Montagné, N.; Darboux, I.; et al. Two Genomes of Highly Polyphagous Lepidopteran Pests (Spodoptera frugiperda, Noctuidae) with Different Host-Plant Ranges. Sci. Rep. 2017, 7, 11816. [Google Scholar] [CrossRef]
- Gui, F.; Lan, T.; Zhao, Y.; Guo, W.; Dong, Y.; Fang, D.; Liu, H.; Li, H.; Wang, H.; Hao, R. Genomic and Transcriptomic Analysis Unveils Population Evolution and Development of Pesticide Resistance in Fall Armyworm Spodoptera frugiperda. Protein Cell 2020. [Google Scholar] [CrossRef]
- Xiao, H.; Ye, X.; Xu, H.; Mei, Y.; Yang, Y.; Chen, X.; Yang, Y.; Liu, T.; Yu, Y.; Yang, W. The Genetic Adaptations of Fall Armyworm Spodoptera frugiperda Facilitated Its Rapid Global Dispersal and Invasion. Mol. Ecol. Resour. 2020, 20, 1050–1068. [Google Scholar] [CrossRef] [PubMed]
- Jakka, S.R.K.; Gong, L.; Hasler, J.; Banerjee, R.; Sheets, J.J.; Narva, K.; Blanco, C.A.; Jurat-Fuentes, J.L. Field-Evolved Mode 1 Resistance of the Fall Armyworm to Transgenic Cry1Fa-Expressing Corn Associated with Reduced Cry1Fa Toxin Binding and Midgut Alkaline Phosphatase Expression. Appl. Environ. Microbiol. 2016, 82, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and Characterization of Field Resistance to Bt Maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Monnerat, R.; Martins, E.; Macedo, C.; Queiroz, P.; Praça, L.; Soares, C.M.; Moreira, H.; Grisi, I.; Silva, J.; Soberon, M.; et al. Evidence of Field-Evolved Resistance of Spodoptera frugiperda to Bt Corn Expressing Cry1F in Brazil That Is Still Sensitive to Modified Bt Toxins. PLoS ONE 2015, 10, e0119544. [Google Scholar] [CrossRef]
- Omoto, C.; Bernardi, O.; Salmeron, E.; Sorgatto, R.J.; Dourado, P.M.; Crivellari, A.; Carvalho, R.A.; Willse, A.; Martinelli, S.; Head, G.P. Field-Evolved Resistance to Cry1Ab Maize by Spodoptera frugiperda in Brazil. Pest Manag. Sci. 2016, 72, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Chandrasena, D.I.; Signorini, A.M.; Abratti, G.; Storer, N.P.; Olaciregui, M.L.; Alves, A.P.; Pilcher, C.D. Characterization of Field-Evolved Resistance to Bacillus Thuringiensis-Derived Cry1F δ-Endotoxin in Spodoptera frugiperda Populations from Argentina. Pest Manag. Sci. 2018, 74, 746–754. [Google Scholar] [CrossRef]
- Schlum, K.A.; Lamour, K.; de Bortoli, C.P.; Banerjee, R.; Meagher, R.; Pereira, E.; Murua, M.G.; Sword, G.A.; Tessnow, A.E.; Viteri Dillon, D.; et al. Whole Genome Comparisons Reveal Panmixia among Fall Armyworm (Spodoptera frugiperda) from Diverse Locations. BMC Genom. 2021, 22, 179. [Google Scholar] [CrossRef]
- Banerjee, R.; Hasler, J.; Meagher, R.; Nagoshi, R.; Hietala, L.; Huang, F.; Narva, K.; Jurat-Fuentes, J.L. Mechanism and DNA-Based Detection of Field-Evolved Resistance to Transgenic Bt Corn in Fall Armyworm (Spodoptera frugiperda). Sci. Rep. 2017, 7, 10877. [Google Scholar] [CrossRef]
- Flagel, L.; Lee, Y.W.; Wanjugi, H.; Swarup, S.; Brown, A.; Wang, J.; Kraft, E.; Greenplate, J.; Simmons, J.; Adams, N.; et al. Mutational Disruption of the ABCC2 Gene in Fall Armyworm, Spodoptera frugiperda, Confers Resistance to the Cry1Fa and Cry1A.105 Insecticidal Proteins. Sci. Rep. 2018, 8, 7255. [Google Scholar] [CrossRef]
- Boaventura, D.; Ulrich, J.; Lueke, B.; Bolzan, A.; Okuma, D.; Gutbrod, O.; Geibel, S.; Zeng, Q.; Dourado, P.M.; Martinelli, S.; et al. Molecular Characterization of Cry1F Resistance in Fall Armyworm, Spodoptera frugiperda from Brazil. Insect Biochem. Mol. Biol. 2020, 116, 103280. [Google Scholar] [CrossRef]
- Guan, F.; Zhang, J.; Shen, H.; Wang, X.; Padovan, A.; Walsh, T.K.; Tay, W.T.; Gordon, K.H.J.; James, W.; Czepak, C.; et al. Whole-Genome Sequencing to Detect Mutations Associated with Resistance to Insecticides and Bt Proteins in Spodoptera frugiperda. Insect Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Whalon, M.E.; Terán-Santofimio, H.; Rodriguez-Maciel, J.C.; DiFonzo, C. Field-Evolved Resistance of the Fall Armyworm (Lepidoptera: Noctuidae) to Synthetic Insecticides in Puerto Rico and Mexico. J. Econ. Entomol. 2019, 112, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, S.; Abdelgaffar, H.; Goff, G.L.; Hilliou, F.; Blanco, C.A.; Hänniger, S.; Bretaudeau, A.; Legeai, F.; Nègre, N.; Jurat-Fuentes, J.L.; et al. Adaptation by Copy Number Variation Increases Insecticide Resistance in the Fall Armyworm. Commun. Biol. 2020, 3, 664. [Google Scholar] [CrossRef] [PubMed]
- Boaventura, D.; Bolzan, A.; Padovez, F.E.; Okuma, D.M.; Omoto, C.; Nauen, R. Detection of a Ryanodine Receptor Target-Site Mutation in Diamide Insecticide Resistant Fall Armyworm, Spodoptera frugiperda. Pest Manag. Sci. 2020, 76, 47–54. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a New Alien Invasive Pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J. Fall Armyworm: Impacts and Implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Pashley, D.P. Host-Associated Genetic Differentiation in Fall Armyworm (Lepidoptera: Noctuidae): A Sibling Species Complex? Ann. Entomol. Soc. Am. 1986, 79, 898–904. [Google Scholar] [CrossRef]
- Pashley, D.P. Host-associated differentiation in armyworms (Lepidoptera: Noctuidae): An allozymic and mitochondrial DNA perspective. In Electrophoretic Studies on Agricultural Pests; Loxdale, H.D., Hollander, J.D., Eds.; Clarendon Press: Oxford, UK, 1989; Volume 39, pp. 103–144. [Google Scholar]
- Dumas, P.; Legeai, F.; Lemaitre, C.; Scaon, E.; Orsucci, M.; Labadie, K.; Gimenez, S.; Clamens, A.-L.; Henri, H.; Vavre, F.; et al. Spodoptera frugiperda (Lepidoptera: Noctuidae) Host-Plant Variants: Two Host Strains or Two Distinct Species? Genetica 2015, 143, 305–316. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Goergen, G.; Plessis, H.D.; van den Berg, J.; Meagher, R. Genetic Comparisons of Fall Armyworm Populations from 11 Countries Spanning Sub-Saharan Africa Provide Insights into Strain Composition and Migratory Behaviors. Sci. Rep. 2019, 9, 8311. [Google Scholar] [CrossRef]
- Yainna, S.; Tay, W.T.; Fiteni, E.; Legeai, F.; Clamens, A.-L.; Gimenez, S.; Frayssinet, M.; Asokan, R.; Kalleshwaraswamy, C.M.; Deshmukh, S.; et al. Genomic Balancing Selection Is Key to the Invasive Success of the Fall Armyworm. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, B.; Zheng, W.; Liu, C.; Zhang, D.; Zhao, S.; Li, Z.; Xu, P.; Wilson, K.; Withers, A.; et al. Genetic Structure and Insecticide Resistance Characteristics of Fall Armyworm Populations Invading China. Mol. Ecol. Resour. 2020, 20, 1682–1696. [Google Scholar] [CrossRef] [PubMed]
- Tay, W.T.; Rane, R.; Padovan, A.; Walsh, T.; Elfekih, S.; Downes, S.; Nam, K.; d’Alençon, E.; Zhang, J.; Wu, Y.; et al. Whole Genome Sequencing of Global Spodoptera frugiperda Populations: Evidence for Complex, Multiple Introductions across the Old World. bioRxiv 2020. [Google Scholar] [CrossRef]
- Boaventura, D.; Martin, M.; Pozzebon, A.; Mota-Sanchez, D.; Nauen, R. Monitoring of Target-Site Mutations Conferring Insecticide Resistance in Spodoptera frugiperda. Insects 2020, 11, 545. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce Framework for Analyzing next-Generation DNA Sequencing Data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zheng, Z.; Cai, Y.; Chen, T.; Li, C.; Fu, W.; Jiang, Y. CNVcaller: Highly Efficient and Widely Applicable Software for Detecting Copy Number Variations in Large Populations. GigaScience 2017, 6. [Google Scholar] [CrossRef]
- Create Your Own Custom Map. Available online: https://mapchart.net/index.html (accessed on 17 May 2021).
- Slater, G.S.C.; Birney, E. Automated Generation of Heuristics for Biological Sequence Comparison. BMC Bioinform. 2005, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The Variant Call Format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Walsh, T.K.; Joussen, N.; Tian, K.; McGaughran, A.; Anderson, C.J.; Qiu, X.; Ahn, S.-J.; Bird, L.; Pavlidi, N.; Vontas, J.; et al. Multiple Recombination Events between Two Cytochrome P450 Loci Contribute to Global Pyrethroid Resistance in Helicoverpa armigera. PLoS ONE 2018, 13, e0197760. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Montoya, W.A.; Elfekih, S.; North, H.L.; Meier, J.I.; Warren, I.A.; Tay, W.T.; Gordon, K.H.; Specht, A.; Paula-Moraes, S.V.; Rane, R. Adaptive Introgression across Semipermeable Species Boundaries between Local Helicoverpa zea and Invasive Helicoverpa armigera Moths. Mol. Biol. Evol. 2020, 37, 2568–2583. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Xiao, Y.; Xu, P.; Yang, X.; Wu, Q.; Wu, K. Insecticide Resistance Monitoring for the Invasive Populations of Fall Armyworm, Spodoptera frugiperda in China. J. Integr. Agric. 2021, 20, 783–791. [Google Scholar] [CrossRef]
- Zhang, W. Global Pesticide Use: Profile, Trend, Cost/Benefit and More. Proc. Int. Acad. Ecol. Environ. Sci. 2018, 8, 1–27. [Google Scholar]
- Lanfear, R.; Kokko, H.; Eyre-Walker, A. Population Size and the Rate of Evolution. Trends Ecol. Evol. 2014, 29, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.; Munch, K.; Mailund, T.; Nater, A.; Greminger, M.P.; Krützen, M.; Marquès-Bonet, T.; Schierup, M.H. Evidence That the Rate of Strong Selective Sweeps Increases with Population Size in the Great Apes. Proc. Natl. Acad. Sci. USA 2017, 114, 1613–1618. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.; Nhim, S.; Robin, S.; Bretaudeau, A.; Nègre, N.; d’Alençon, E. Positive Selection Alone Is Sufficient for Whole Genome Differentiation at the Early Stage of Speciation Process in the Fall Armyworm. BMC Evol. Biol. 2020, 20, 152. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutations | Individuals with Resistance Mutations |
|---|---|
| 2 bp insertion and a frameshift mutation | * PR1, PR5, PR12, PR14, PR16, * PR18, * PR19, * PR27, PR30, * PR31, * PR33 |
| GY deletion | CC44, CC69 |
| P799K/R | CC44, CC69 |
| 12 bp insertion and a frameshift mutation | * rCC25, * rCC5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yainna, S.; Nègre, N.; Silvie, P.J.; Brévault, T.; Tay, W.T.; Gordon, K.; dAlençon, E.; Walsh, T.; Nam, K. Geographic Monitoring of Insecticide Resistance Mutations in Native and Invasive Populations of the Fall Armyworm. Insects 2021, 12, 468. https://doi.org/10.3390/insects12050468
Yainna S, Nègre N, Silvie PJ, Brévault T, Tay WT, Gordon K, dAlençon E, Walsh T, Nam K. Geographic Monitoring of Insecticide Resistance Mutations in Native and Invasive Populations of the Fall Armyworm. Insects. 2021; 12(5):468. https://doi.org/10.3390/insects12050468
Chicago/Turabian StyleYainna, Sudeeptha, Nicolas Nègre, Pierre J. Silvie, Thierry Brévault, Wee Tek Tay, Karl Gordon, Emmanuelle dAlençon, Thomas Walsh, and Kiwoong Nam. 2021. "Geographic Monitoring of Insecticide Resistance Mutations in Native and Invasive Populations of the Fall Armyworm" Insects 12, no. 5: 468. https://doi.org/10.3390/insects12050468
APA StyleYainna, S., Nègre, N., Silvie, P. J., Brévault, T., Tay, W. T., Gordon, K., dAlençon, E., Walsh, T., & Nam, K. (2021). Geographic Monitoring of Insecticide Resistance Mutations in Native and Invasive Populations of the Fall Armyworm. Insects, 12(5), 468. https://doi.org/10.3390/insects12050468