
Susceptibility of Tuta absoluta (Lepidoptera: Gelechiidae) Pupae to Soil Applied Entomopathogenic Fungal Biopesticides


Abstract
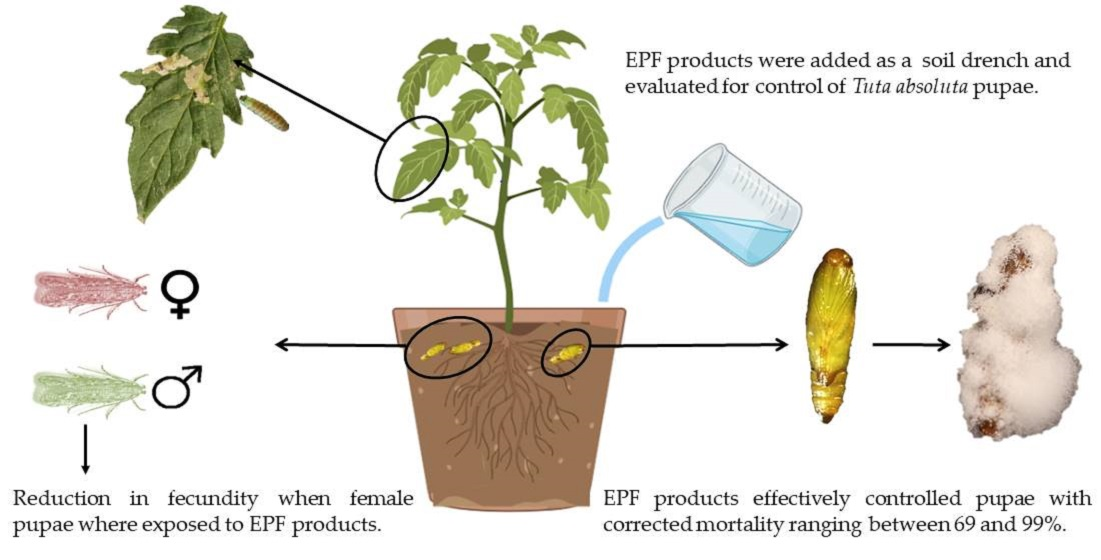
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Entomopathogenic Fungi
2.3. Laboratory Dose-Response Bioassays
2.4. Growth Tunnel Experiment
2.5. Effect on Fecundity and Fertility
2.6. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brévault, T.; Sylla, S.; Diatte, M.; Bernadas, G.; Diarra, K. Tuta absoluta Meyrick (Lepidoptera: Gelechiidae): A New Threat to Tomato Production in Sub-Saharan Africa. Afr. Entomol. 2014, 22, 441–444. [Google Scholar] [CrossRef]
- de Campos, M.R.; Biondi, A.; Adiga, A.; Guedes, R.N.C.; Desneux, N. From the Western Palaearctic region to beyond: Tuta absoluta 10 years after invading Europe. J. Pest Sci. 2017, 90, 787–796. [Google Scholar] [CrossRef]
- Chidege, M.; Al-Zaidi, S.; Hassan, N.; Julie, A.; Kaaya, E.; Mrogoro, S. First record of tomato leaf miner Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) in Tanzania. Agric. Food Secur. 2016, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Ruescas, D.C.; Tabone, E.; Frandon, J.; et al. Biological invasion of European tomato crops by Tuta absoluta: Ecology, geographic expansion and prospects for biological control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Idriss, G.E.A.; Mohamed, S.A.; Khamis, F.; Du Plessis, H.; Ekesi, S. Biology and performance of two indigenous larval parasitoids on Tuta absoluta (Lepidoptera: Gelechiidae) in Sudan. Biocontrol Sci. Technol. 2018, 28, 614–628. [Google Scholar] [CrossRef]
- Pfeiffer, D.G.; Muniappan, R.; Sall, D.; Diatta, P.; Diongue, A.; Dieng, E.O. First Record of Tuta absoluta (Lepidoptera: Gelechiidae) in Senegal. Fla. Entomol. 2013, 96, 661–662. [Google Scholar] [CrossRef]
- Urbaneja, A.; Vercher, R.; Navarro, V.; Garcıa, M.F.; Porcuna, J.L. La polilla del tomate, Tuta absoluta. Phytoma España 2007, 194, 16–23. [Google Scholar]
- Visser, D.; Uys, V.; Nieuwenhuis, R.; Pieterse, W. First records of the tomato leaf miner Tuta absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae) in South Africa. BioInvasions Rec. 2017, 6, 301–305. [Google Scholar] [CrossRef]
- Biondi, A.; Guedes, R.N.C.; Wan, F.-H.; Desneux, N. Ecology, Worldwide Spread, and Management of the Invasive South American Tomato Pinworm, Tuta absoluta: Past, Present, and Future. Annu. Rev. Entomol. 2018, 63, 239–258. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Picanço, M.C. The tomato borer Tuta absoluta in South America: Pest status, management and insecticide resistance. EPPO Bull. 2012, 42, 211–216. [Google Scholar] [CrossRef]
- Roditakis, E.; Grispou, M.; Morou, E.; Kristofferson, J.B.; Roditakis, N.; Nauen, R.; Vontas, J.; Tsagkarakou, A. Current status of insecticide resistance in Q biotype Bemisia tabaci populations from Crete. Pest Manag. Sci. 2009, 65, 313–322. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Y. High Levels of Resistance to Chlorantraniliprole Evolved in Field Populations of Plutella xylostella. J. Econ. Entomol. 2012, 105, 1019–1023. [Google Scholar] [CrossRef]
- El-Arnaouty, S.A.; Pizzol, J.; Galal, H.H.; Kortam, M.N.; Afifi, A.I.; Beyssat, V.; Desneux, N.; Biondi, A.; Heikal, I.H. Assessment of two Trichogramma species for the control of Tuta absoluta in North African tomato greenhouses. Afr. Entomol. 2014, 22, 801–809. [Google Scholar] [CrossRef]
- Mandour, N.; Sarhan, A.; Atwa, D. The Integration between Trichogramma evanescens West. (Hymenoptera: Trichogrammatidae) and Selected Bioinsecticides for Controlling the Potato Tuber Moth Phthorimaea operculella (Zell.) (Lepidoptera: Gelechiidae) of Stored Potatoes. J. Plant Prot. Res. 2012, 52, 40–46. [Google Scholar] [CrossRef]
- Senior, L.J.; McEwen, P.K. The use of lacewings in biological control. In Lacewings in the Crop Environment; McEwen, P.K., New, T.R., Washington, A.E., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 296–302. [Google Scholar]
- Shalaby, H.H.; Faragalla, F.H.; El-Saadany, H.M.; Ibrahim, A.A. Efficacy of three entomopathogenic agents for control the tomato borer, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Nat. Sci. 2013, 11, 63–64. [Google Scholar]
- Never, Z.; Patrick, A.N.; Musa, C.; Ernest, M.; Zekeya, N.; Ndakidemi, P.A.; Chacha, M.; Mbega, E. Tomato Leafminer, Tuta absoluta (Meyrick 1917), an emerging agricultural pest in Sub-Saharan Africa: Current and prospective management strategies. Afr. J. Agric. Res. 2017, 12, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Ferracini, C.; Bueno, V.H.P.; Dindo, M.L.; Ingegno, B.L.; Luna, M.G.; Gervassio, N.G.S.; Sánchez, N.E.; Siscaro, G.; Van Lenteren, J.C.; Zappalà, L.; et al. Natural enemies of Tuta absoluta in the Mediterranean basin, Europe and South America. Biocontrol Sci. Technol. 2019, 29, 578–609. [Google Scholar] [CrossRef]
- Coombes, C.A.; Hill, M.P.; Moore, S.D.; Dames, J.F. Potential of entomopathogenic fungal isolates for control of the soil-dwelling life stages of Thaumatotibia leucotreta Meyrick (Lepidoptera: Tortricidae) in citrus. Afr. Entomol. 2017, 25, 235–238. [Google Scholar] [CrossRef]
- Garrido-Jurado, I.; Ruano, F.; Campos, M.; Quesada-Moraga, E. Effects of soil treatments with entomopathogenic fungi on soil dwelling non-target arthropods at a commercial olive orchard. Biol. Control 2011, 59, 239–244. [Google Scholar] [CrossRef]
- Alikhani, M.; Safavi, S.A.; Iranipour, S. Effect of the entomopathogenic fungus, Metarhizium anisopliae (Metschnikoff) Sorokin, on demographic fitness of the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Egypt. J. Biol. Pest Control 2019, 29, 23. [Google Scholar] [CrossRef]
- Klieber, J.; Reineke, A. The entomopathogen Beauveria bassiana has epiphytic and endophytic activity against the tomato leaf miner Tuta absoluta. J. Appl. Entomol. 2016, 140, 580–589. [Google Scholar] [CrossRef]
- Tadele, S.; Emana, G. Entomopathogenic effect of Beauveria bassiana (Bals.) and Metarhizium anisopliae (Metschn.) on Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) larvae under laboratory and glasshouse conditions in Ethiopia. J. Plant Pathol. Microbiol. 2017, 8, 411–441. [Google Scholar]
- Contreras, J.; Mendoza, J.E.; Martínez-Aguirre, M.R.; García-Vidal, L.; Izquierdo, J.; Bielza, P. Efficacy of Enthomopathogenic Fungus Metarhizium anisopliae against Tuta absoluta (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2014, 107, 121–124. [Google Scholar] [CrossRef]
- Genç, H. The tomato leafminer, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae):pupal key characters for sexing individuals. Turk. J. Zool. 2016, 40, 801–805. [Google Scholar] [CrossRef]
- Shanmugam, V.; Seethapathy, P. Isolation and characterization of white muscardine fungi Beauveria bassiana (Bals.) Vuill.—A causative of mulberry silkworm. J. Entomol. Zool. Stud. 2017, 5, 512–515. [Google Scholar]
- France, A.; Gerding-González, M.; Gerding, M.; Sandoval, A. Patogenicidad de una colección de cepas nativas de Metarhizium spp. y Beauveria spp. en Aegorhinus superciliosus, Asynonychus cervinus y Otiorhynchus sulcatus. Agric. Técnica 2000, 60, 3. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- TIBCO Software, Inc. Statistica (Data Analysis Software System), Version 13.3. 2017. Available online: www.tibco.com (accessed on 15 November 2020).
- Mansour, R.; Brévault, T.; Chailleux, A.; Cherif, A.; Grissa-Lebdi, K.; Haddi, K.; Mohamed, S.A.; Nofemela, R.S.; Oke, A.; Sylla, S.; et al. Occurrence, biology, natural enemies and management of Tuta absoluta in Africa. Entomol. Gen. 2018, 38, 83–112. [Google Scholar] [CrossRef]
- Jaronski, S.T. Ecological factors in the inundative use of fungal entomopathogens. BioControl 2010, 55, 159–185. [Google Scholar] [CrossRef]
- Kabaluk, J.T.; Ericsson, J.D. Environmental and behavioral constraints on the infection of wireworms by Metarhizium anisopliae. Environ. Entomol. 2007, 36, 1415–1420. [Google Scholar] [CrossRef]
- Mkiga, A.M.; Mohamed, S.A.; Du Plessis, H.; Khamis, F.M.; Akutse, K.S.; Ekesi, S. Metarhizium anisopliae and Beauveria bassiana: Pathogenicity, Horizontal Transmission, and Their Effects on Reproductive Potential of Thaumatotibia leucotreta (Lepidoptera: Tortricidae). J. Econ. Entomol. 2020, 113, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Bajracharya, A.S.R.; Bhat, B. Life cycle of South American tomato leaf miner, Tuta absoluta (Meyrick, 1917) in Nepal. J. Entomol. Zool. Stud. 2018, 6, 287–290. [Google Scholar]
- Kleespies, R.G.; Zimmermann, G. Effect of Additives on the Production, Viability and Virulence of Blastospores of Metarhizium anisopliae. Biocontrol Sci. Technol. 1998, 8, 207–214. [Google Scholar] [CrossRef]
- Abdel-Raheem, M.A.; Ismail, I.A.; Abdel-Rahman, R.S.; Abdel-Rhman, I.E.; Reyad, N.F. Efficacy of three entomopathogenic fungi on tomato leaf miner, Tuta absoluta in tomato crop in Egypt. J. Agric. Res. 2015, 1, 15–21. [Google Scholar]
- İnanlı, C.; Yoldas, Z.; Birgücü, A.K. Effects of entomopathogenic fungi, Beauveria bassiana (Bals.) and Metarhizium anisopliae (Metsch.) on larvae and egg stages of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Ege Üniversitesi Ziraat Fakültesi Dergisi 2012, 49, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.; Gerding, M.; France, A. Effectivity of entomopathogenic fungus strains on tomato moth Tuta absoluta Meyrick (Lepidoptera: Gelechiidae) larvae. Agric. Técnica 2006, 66, 159–165. [Google Scholar]
- Garrido-Jurado, I.; Torrent, J.; Barrón, V.; Corpas, A.; Quesada-Moraga, E. Soil properties affect the availability, movement, and virulence of entomopathogenic fungi conidia against puparia of Ceratitis capitata (Diptera: Tephritidae). Biol. Control 2011, 58, 277–285. [Google Scholar] [CrossRef]
- Ignoffo, C.M.; Hostetter, D.L.; Pinnell, R.E.; Garcia, C. Vertical Movement of Conidia of Nomuraea rileyi through Sand and Loam Soils. J. Econ. Entomol. 1977, 70, 163–164. [Google Scholar] [CrossRef]
- Uchoa-Fernandes, M.A.; Vilela, E.F.; Della Lucia, T.M.C. Ritmo diário de atratao sexual em Scrobipalpuloides absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae). Rev. Bras. Biol. 1995, 55, 67–73. [Google Scholar]
- Kader, M.; Senge, M.; Mojid, M.; Ito, K. Recent advances in mulching materials and methods for modifying soil environment. Soil Tillage Res. 2017, 168, 155–166. [Google Scholar] [CrossRef]
- Martín-Closas, L.; Costa, J.; Pelacho, A.M. Agronomic effects of biodegradable films on crop and field environment. In Soil Degradable Bioplastics for a Sustainable Modern Agriculture; Malinconico, M., Ed.; Springer: Berlin, Germany, 2017; pp. 67–104. [Google Scholar]
- Bandopadhyay, S.; Martin-Closas, L.; Pelacho, A.M.; DeBruyn, J.M. Biodegradable plastic mulch films: Impacts on soil microbial communities and ecosystem functions. Front. Microbiol. 2018, 9, 819. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
| Commercial Name and Manufacturer | ACTIVE Ingredient | Experimental Name | Batch Concentration | Target Pest Registration |
|---|---|---|---|---|
| Metarril WP E9 Koppert SA (Pty) (Falcon Ln, Lanseria, 1739, South Africa) | Metarhizium anisopliae (Metsch.) Sorok., strain E9 | MET-WP | 2.00 × 109 conidia/g | Mahanarva fimbriolata (Stål) (Hemiptera: Cercopidae) |
| Real Metarhizium 69 Real IPM (SA) Ltd. (Grabouw, 7160, South Africa) | Metarhizium anisopliae (Metsch.) Sorok., strain ICIPE 69 | MET-ES | 3.23 × 109 conidia/mL | Fruit flies (Diptera) Mealybug (Hemiptera) Weevils (Coleoptera) Thrips (Thysanoptera) Whiteflies (Hemiptera) |
| Eco-Bb® Plant Health Products (Pty) (Nottingham Road, 3280, South Africa) | Beauveria bassiana (Bals.) Vuill., strain R444 | BB-WP | 2.20 × 109 conidia/g | Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) Tetranychus spp. (Acari: Tetranychidae) Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae) Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) Whiteflies (Hemiptera) |
| Real Beauveria Real IPM (SA) Ltd. (Grabouw, 7160, South Africa) | Beauveria bassiana (Bals.) Vuill., strain BB02 | BB-ES | 2.74 × 107 conidia/mL | Thrips (Thysanoptera) |
| M. Anisopliae-ES | B. Bassiana-ES |
|---|---|
| M. anisopliae (Male) × M. anisopliae (Female) MM × MF | B. bassiana (Male) × B. bassiana (Female) BM × BF |
| M. anisopliae (Male) × Control (Female) MM × CF | B. bassiana (Male) × Control (Female) BM × CF |
| Control (Male) × M. anisopliae (Female) CM × MF | Control (Male) × B. bassiana (Female) CM × BF |
| Control | |
| Control (Male) × Control (Female) CM × CF | |
| Treatment | n a (df) | LC50 | FL (95%) b | LC80 | FL (95%) | Slope | SE | Χ2 c | n d | Mean Corrected Mortality (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| BB-ES | 280 (5) | 1.87 × 107 | 1.3 × 107–2.90 × 107 a | 1.09 × 108 | 6.25 × 107–2.50 × 108 | 1.10 | 0.13 | 2.57 | 100 | 98.8 ± 1.3 a |
| MET-ES | 240 (4) | 1.38 × 108 | 9.40 × 107–1.88 × 108 b | 4.31 × 108 | 3.15 × 108–6.38 × 108 | 1.70 | 0.19 | 2.45 | 100 | 90.0 ± 4.1 ab |
| MET-WP | 329 (6) | 3.84 × 109 | 2.82 × 109–5.5 × 109 d | 1.73 × 1010 | 1.05 × 1010–4.25 × 1010 | 1.29 | 0.18 | 5.4 | 100 | 88.8 ± 4.4 ab |
| BB-WP | 350 (5) | 1.49 × 109 | 1.02 × 109–2.0 × 109 c | 5.26 × 109 | 3.9 × 109–7.34 × 109 | 1.53 | 0.16 | 3.67 | 100 | 68.8 ± 6.3 b |
| H(3) = 13.67 |
| Treatment | n | Mean Fecundity per Female | Mean Percentage Fertility |
|---|---|---|---|
| BM × BF | 10 | 33.00 ± 9.59 b * | 93.99 ± 2.48 a * |
| BM × CF | 9 | 57.67 ± 17.40 ab | 92.19 ± 2.33 a |
| CM × BF | 10 | 33.90 ± 9.17 b | 92.40 ± 4.43 a |
| MM × MF | 10 | 35.60 ± 8.06 b | 96.08 ± 1.50 a |
| MM × CF | 10 | 57.90 ± 10.11 ab | 95.55 ± 1.63 a |
| CM × MF | 9 | 67.33 ± 12.03 ab | 97.26 ± 1.18 a |
| CM × CF | 10 | 85.60 ± 11.41 a | 96.56 ± 1.22 a |
| Treatments | n a | Mean Percentage Moth Emergence c | n b | Mean Percentage Mycosed Moths d |
|---|---|---|---|---|
| BB-ES | 157 | 31.60 ± 4.82 a | 39 | 82.05 ± 6.22 a |
| MET-ES | 189 | 65.82 ± 4.32 b | 43 | 6.98 ± 3.93 b |
| Control | 102 | 80.50 ± 3.20 b | 60 | 0.00 ± 0.00 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erasmus, R.; van den Berg, J.; du Plessis, H. Susceptibility of Tuta absoluta (Lepidoptera: Gelechiidae) Pupae to Soil Applied Entomopathogenic Fungal Biopesticides. Insects 2021, 12, 515. https://doi.org/10.3390/insects12060515
Erasmus R, van den Berg J, du Plessis H. Susceptibility of Tuta absoluta (Lepidoptera: Gelechiidae) Pupae to Soil Applied Entomopathogenic Fungal Biopesticides. Insects. 2021; 12(6):515. https://doi.org/10.3390/insects12060515
Chicago/Turabian StyleErasmus, Reynardt, Johnnie van den Berg, and Hannalene du Plessis. 2021. "Susceptibility of Tuta absoluta (Lepidoptera: Gelechiidae) Pupae to Soil Applied Entomopathogenic Fungal Biopesticides" Insects 12, no. 6: 515. https://doi.org/10.3390/insects12060515