
Hyperparasitism of Acroclisoides sinicus (Huang and Liao) (Hymenoptera: Pteromalidae) on Two Biological Control Agents of Halyomorpha halys



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Origin of Insects
2.2. No-Choice Tests
2.3. Two-Choice Test
2.4. Olfactometer Trial
2.5. Assessment of Acroclisoides sinicus Egg Load
2.6. Statistical Analysis
3. Results
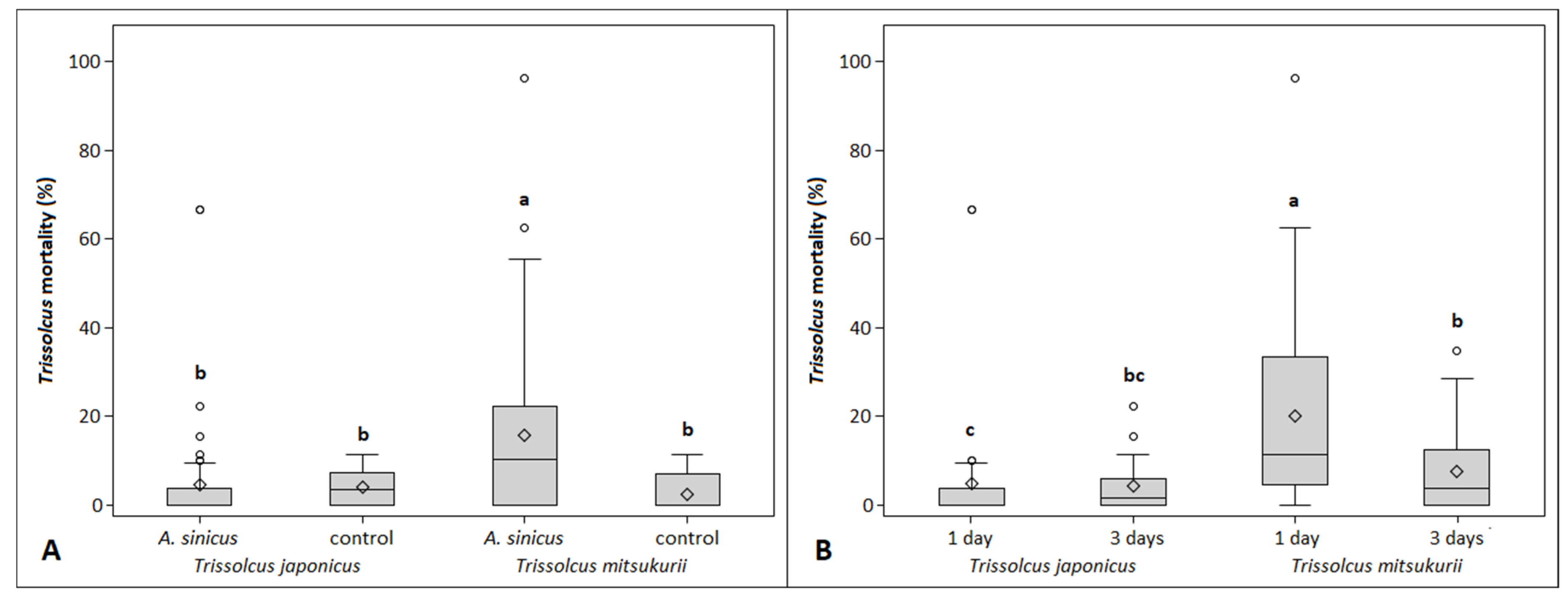
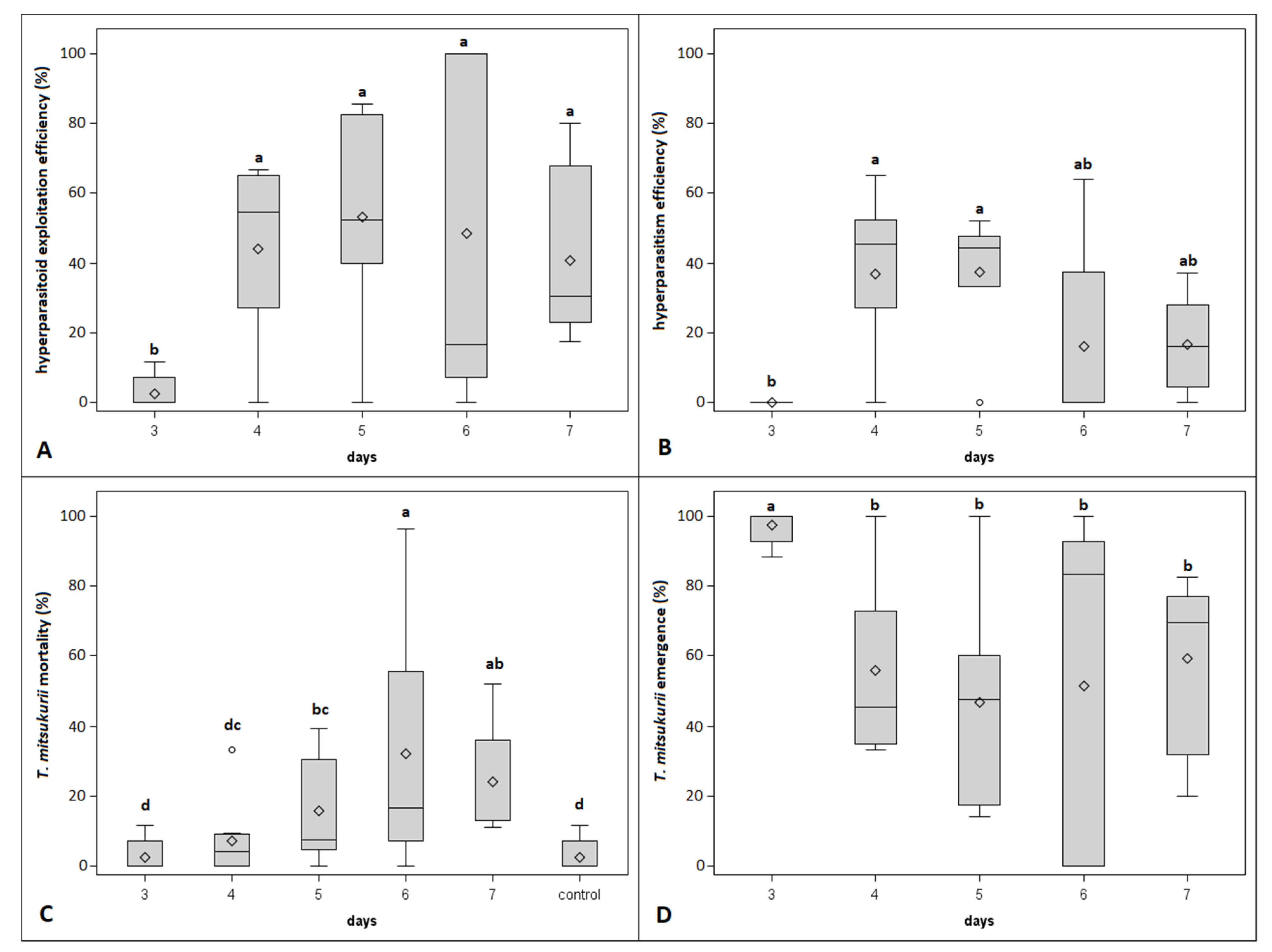
3.1. No-Choice Experiment
3.2. Two-Choice Experiment
3.3. Olfactometer Trial
3.4. Assessment of Acroclisoides sinicus Egg Load
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive Brown Marmorated Stink Bug in North America and Europe: History, Biology, Ecology, and Management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [Green Version]
- Maistrello, L.; Dioli, P.; Vaccari, G.; Caruso, S. First records in Italy of the Asian Stinkbug Halyomorpha halys, a new Threat for Fruit Crops. ATTI Giornate Fitopatol. 2014, 1, 283–288. [Google Scholar]
- Maistrello, L.; Vaccari, G.; Caruso, S.; Costi, E.; Bortolini, S.; Macavei, L.; Foca, G.; Ulrici, A.; Bortolotti, P.P.; Nannini, R.; et al. Monitoring of the invasive Halyomorpha halys, a new key pest of fruit orchards in northern Italy. J. Pest Sci. 2017, 90, 1231–1244. [Google Scholar] [CrossRef]
- Bariselli, M.; Bugiani, R.; Maistrello, L. Distribution and damage caused by Halyomorpha halys in Italy. EPPO Bull. 2016, 46, 332–334. [Google Scholar] [CrossRef]
- Moore, L.; Tirello, P.; Scaccini, D.; Toews, M.D.; Duso, C.; Pozzebon, A. Characterizing damage potential of the brown marmorated stink bug in cherry orchards in Italy. Entomol. Gen. 2019, 39, 271–283. [Google Scholar] [CrossRef]
- Bosco, L.; Moraglio, S.T.; Tavella, L. Halyomorpha halys, a serious threat for hazelnut in newly invaded areas. J. Pest Sci. 2018, 91, 661–670. [Google Scholar] [CrossRef]
- Zapponi, L.; Tortorici, F.; Anfora, G.; Bardella, S.; Bariselli, M.; Benvenuto, L.; Bernardinelli, I.; Butturini, A.; Caruso, S.; Colla, R.; et al. Assessing the Distribution of Exotic Egg Parasitoids of Halyomorpha halys in Europe with a Large-Scale Monitoring Program. Insects 2021, 12, 316. [Google Scholar] [CrossRef]
- Moraglio, S.T.; Tortorici, F.; Pansa, M.G.; Castelli, G.; Pontini, M.; Scovero, S.; Visentin, S.; Tavella, L. A 3-year survey on parasitism of Halyomorpha halys by egg parasitoids in northern Italy. J. Pest Sci. 2020, 93, 183–194. [Google Scholar] [CrossRef]
- Scaccini, D.; Falagiarda, M.; Tortorici, F.; Martinez-Sañudo, I.; Tirello, P.; Reyes-Domínguez, Y.; Gallmetzer, A.; Tavella, L.; Zandigiacomo, P.; Duso, C.; et al. An insight into the role of Trissolcus mitsukurii as biological control agent of Halyomorpha halys in Northeastern Italy. Insects 2020, 11, 306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, F.; Gariepy, T.; Mason, P.; Gillespie, D.; Talamas, E.; Haye, T. Seasonal parasitism and host specificity of Trissolcus japonicus in northern China. J. Pest Sci. 2017, 90, 1127–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila, G.A.; Chen, J.-H.; Li, W.; Alavi, M.; Mi, Q.; Sandanayaka, M.; Zhang, F.; Zhang, J. Seasonal Abundance and Diversity of Egg Parasitoids of Halyomorpha halys in Kiwifruit Orchards in China. Insects 2021, 12, 428. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Yao, Y.X.; Qiu, L.F.; Li, Z.X. A new species of Trissolcus (Hymenoptera: Scelionidae) parasitizing eggs of Halyomorpha halys (Heteroptera: Pentatomidae) in China with comments on its biology. Ann. Entomol. Soc. Am. 2009, 102, 39–47. [Google Scholar] [CrossRef]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the Biology, Ecology, and Management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef]
- Haye, T.; Fischer, S.; Zhang, J.; Gariepy, T. Can native egg parasitoids adopt the invasive brown marmorated stink bug, Halyomorpha halys (Heteroptera: Pentatomidae), in Europe? J. Pest Sci. 2015, 88, 693–705. [Google Scholar] [CrossRef]
- Stahl, J.M.; Babendreier, D.; Haye, T. Using the egg parasitoid Anastatus bifasciatus against the invasive brown marmorated stink bug in Europe: Can non-target effects be ruled out? J. Pest Sci. 2018, 91, 1005–1017. [Google Scholar] [CrossRef] [Green Version]
- Stahl, J.M.; Babendreier, D.; Marazzi, C.; Caruso, S.; Costi, E.; Maistrello, L.; Haye, T. Can Anastatus bifasciatus be used for augmentative biological control of the brown marmorated stink bug in fruit orchards? Insects 2019, 10, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraglio, S.T.; Tortorici, F.; Visentin, S.; Pansa, M.G.; Tavella, L. Trissolcus kozlovi in North Italy: Host Specificity and Augmentative Releases against Halyomorpha halys in Hazelnut Orchards. Insects 2021, 12, 464. [Google Scholar] [CrossRef]
- Talamas, E.J.; Herlihy, M.V.; Dieckhoff, C.; Hoelmer, K.A.; Buffington, M.; Bon, M.-C.; Weber, D.C. Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae) emerges in North America. J. Hymenopt. Res. 2015, 43, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Stahl, J.; Tortorici, F.; Pontini, M.; Bon, M.-C.; Hoelmer, K.; Marazzi, C.; Tavella, L.; Haye, T. First discovery of adventive populations of Trissolcus japonicus in Europe. J. Pest Sci. 2019, 92, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini Peverieri, G.; Talamas, E.; Bon, M.C.; Marianelli, L.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Roversi, P.F.; Hoelmer, K. Two Asian egg parasitoids of Halyomorpha halys (Stål) (Hemiptera, Pentatomidae) emerge in northern Italy: Trissolcus mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J. Hymenopt. Res. 2018, 67, 37–53. [Google Scholar] [CrossRef]
- Dieckhoff, C.; Wenz, S.; Renninger, M.; Reißig, A.; Rauleder, H.; Zebitz, C.P.W.; Reetz, J.; Zimmermann, O. Add Germany to the List—Adventive Population of Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae) Emerges in Germany. Insects 2021, 12, 414. [Google Scholar] [CrossRef] [PubMed]
- Decreto 42967 del 9 giugno 2020. Immissione in natura della specie non autoctona Trissolcus japonicus quale Agente di Controllo Biologico del fitofago Halyomorpha halys ai sensi del Decreto del Presidente della Repubblica 8 settembre 1997, n. 357, art. 12. 2020. Available online: http://agricoltura.regione.campania.it/difesa/files/DM_halyomorpha.pdf (accessed on 3 July 2021).
- Grissell, E.E.; Smith, D.R. First report of Acroclisoides Girault and Dodd (Hymenoptera: Pteromalidae) in the western hemisphere, with description of a new species. Proc. Entomol. Soc. Washingt. 2006, 108, 923–929. [Google Scholar]
- Sabbatini Peverieri, G.; Mitroiu, M.-D.; Bon, M.-C.; Balusu, R.; Benvenuto, L.; Bernardinelli, I.; Fadamiro, H.; Falagiarda, M.; Fusu, L.; Grove, E.; et al. Surveys of stink bug egg parasitism in Asia, Europe and North America, morphological taxonomy, and molecular analysis reveal the Holarctic distribution of Acroclisoides sinicus (Huang & Liao) (Hymenoptera, Pteromalidae). J. Hymenopt. Res. 2019, 74, 123–151. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.R.; Seymour, J.E. Two species of Acroclisoides Girault and Dodd (Hymenoptera: Pteromalidae) parasitic on Trissolcus basalis (Wollaston) (Hymenoptera: Scelionidae), a parasitoid of Nezara viridula (L.) (Hemiptera: Pentatomidae). Aust. J. Entomol. 1992, 31, 299–300. [Google Scholar] [CrossRef]
- Tillman, G.; Toews, M.; Blaauw, B.; Sial, A.; Cottrell, T.; Talamas, E.; Buntin, D.; Joseph, S.; Balusu, R.; Fadamiro, H.; et al. Parasitism and predation of sentinel eggs of the invasive brown marmorated stink bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae), in the southeastern US. Biol. Control 2020, 145, 104247. [Google Scholar] [CrossRef]
- Gariepy, T.D.; Haye, T.; Zhang, J. A molecular diagnostic tool for the preliminary assessment of host-parasitoid associations in biological control programmes for a new invasive pest. Mol. Ecol. 2014, 23, 3912–3924. [Google Scholar] [CrossRef]
- Giovannini, L.; Sabbatini-Peverieri, G.; Tillman, P.G.; Hoelmer, K.A.; Roversi, P.F. Reproductive and Developmental Biology of Acroclisoides sinicus, a Hyperparasitoid of Scelionid Parasitoids. Biology 2021, 10, 229. [Google Scholar] [CrossRef]
- Sullivan, D.J. Insect Hyperparasitism. Annu. Rev. Entomol. 1987, 32, 49–70. [Google Scholar] [CrossRef]
- Nofemela, R.S. The effect of obligate hyperparasitoids on biological control: Differential vulnerability of primary parasitoids to hyperparasitism can mitigate trophic cascades. Biol. Control 2013, 65, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini Peverieri, G.; Dieckhoff, C.; Giovannini, L.; Marianelli, L.; Roversi, P.F.; Hoelmer, K. Rearing Trissolcus japonicus and Trissolcus mitsukurii for biological control of Halyomorpha halys. Insects 2020, 11, 787. [Google Scholar] [CrossRef]
- Talamas, E.J.; Buffington, M.L.; Hoelmer, K. Revision of palearctic Trissolcus Ashmead (Hymenoptera, Scelionidae). J. Hymenopt. Res. 2017, 56, 3–185. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini Peverieri, G.; Giovannini, L.; Benvenuti, C.; Madonni, L.; Hoelmer, K.; Roversi, P.F. Characteristics of the meconia of European egg parasitoids of Halyomorpha halys. J. Hymenopt. Res. 2020, 77, 187–201. [Google Scholar] [CrossRef]
- Mayer, C.J.; Vilcinskas, A.; Gross, J. Pathogen-induced Release of Plant Allomone Manipulates Vector Insect Behavior. J. Chem. Ecol. 2008, 34, 1518–1522. [Google Scholar] [CrossRef]
- Belda, C.; Riudavets, J. Attraction of the parasitoid Anisopteromalus calandrae (Howard) (Hymenoptera: Pteromalidae) to odors from grain and stored product pests in a Y-tube olfactometer. Biol. Control 2010, 54, 29–34. [Google Scholar] [CrossRef]
- Papaj, D.R. Ovarian Dynamics and Host Use. Annu. Rev. Entomol. 2000, 45, 423–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jervis, M.A.; Copland, M.J.W.; Harvey, J.A. The Life-cycle. In Insects as Natural Enemies; Springer: Dordrecht, The Netherlands, 2007; pp. 73–165. [Google Scholar]
- Donaldson, J.S.; Walter, G.H. Effects of egg availability and egg maturity on the ovipositional activity of the parasitic wasp, Coccophagus atratus. Physiol. Entomol. 1988, 13, 407–417. [Google Scholar] [CrossRef]
- Bin, F.; Vinson, S.B. Efficacy assessment in egg parasitoids (Hymenoptera): Proposal for a unified terminology. In Proceedings of the Trichogramma and Other Egg Parasitoids 3rd International Symposium on Le Colloques de l’INRA, San Antonio, TX, USA, 23–27 September 1990; Wajnberg, E., Vinson, S.B., Eds.; Institut National de la Recherche Agronomique: Paris, France, 1991; Volume 56, p. 246. [Google Scholar]
- SAS Institute Inc. PROC User’s Manual, 6th ed.; SAS Institute: Cary, NC, USA, 2016; Available online: http://support.sas.com/documentation/cdl/en/indbug/68442/PDF/default/indbug.pdf (accessed on 15 March 2021).
- Costi, E.; Haye, T.; Maistrello, L. Surveying native egg parasitoids and predators of the invasive Halyomorpha halys in Northern Italy. J. Appl. Entomol. 2019, 143, 299–307. [Google Scholar] [CrossRef]
- Campbell, R.W. Some Ichneumonid-Sarcophagid Interactions in the Gypsy Moth Porthetria dispar (L.) (Lepidoptera: Lymantriidae). Can. Entomol. 1963, 95, 337–345. [Google Scholar] [CrossRef]
- Jervis, M.A.; Kidd, N.A.C. Host-Feeding Strategies in Hymenopteran Parasitoids. Biol. Rev. 1986, 61, 395–434. [Google Scholar] [CrossRef]
- Strand, M.R.; Ratner, S.; Vinson, S.B. Maternally induced host regulation by the egg parasitoid Telenomus heliothidis. Physiol. Entomol. 1983, 8, 469–475. [Google Scholar] [CrossRef]
- Asgari, S.; Rivers, D.B. Venom Proteins from Endoparasitoid Wasps and Their Role in Host-Parasite Interactions. Annu. Rev. Entomol. 2011, 56, 313–335. [Google Scholar] [CrossRef]
- Desneux, N.; Barta, R.J.; Hoelmer, K.A.; Hopper, K.R.; Heimpel, G.E. Multifaceted determinants of host specificity in an aphid parasitoid. Oecologia 2009, 160, 387–398. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Bale, J.; Bigler, F.; Hokkanen, H.M.T.; Loomans, A.J.M. Assessing risks of releasing exotic biological control agents of arthropod pests. Annu. Rev. Entomol. 2006, 51, 609–634. [Google Scholar] [CrossRef] [Green Version]
- Louda, S.M.; Pemberton, R.W.; Johnson, M.T.; Follett, P.A. Nontarget Effects—The Achilles’ Heel of Biological Control? Retrospective Analyses to Reduce Risk Associated with Biocontrol Introductions. Annu. Rev. Entomol. 2003, 48, 365–396. [Google Scholar] [CrossRef] [PubMed]
- Murray, T.J.; Withers, T.M.; Mansfield, S. Choice versus no-choice test interpretation and the role of biology and behavior in parasitoid host specificity tests. Biol. Control 2010, 52, 153–159. [Google Scholar] [CrossRef]
- Vinson, S.B. Host Selection by Insect Parasitoids. Annu. Rev. Entomol. 1976, 21, 109–133. [Google Scholar] [CrossRef]
- Abram, P.K.; Cusumano, A.; Abram, K.; Colazza, S.; Peri, E. Testing the habituation assumption underlying models of parasitoid foraging behavior. PeerJ 2017, 5, e3097. [Google Scholar] [CrossRef]
- Cusumano, A.; Harvey, J.A.; Bourne, M.E.; Poelman, E.H.; de Boer, J.G. Exploiting chemical ecology to manage hyperparasitoids in biological control of arthropod pests. Pest Manag. Sci. 2020, 76, 432–443. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, D.J.; Völkl, W. Hyperparasitism: Multitrophic ecology and behavior. Annu. Rev. Entomol. 1999, 44, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Rosi, M.C.; Isidoro, N.; Colazza, S.; Bin, F. Source of the host marking pheromone in the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). J. Insect Physiol. 2001, 47, 989–995. [Google Scholar] [CrossRef]
- Jervis, M.A.; Heimpel, G.E.; Ferns, P.N.; Harvey, J.A.; Kidd, N.A.C. Life-history strategies in parasitoid wasps: A comparative analysis of “ovigeny”. J. Anim. Ecol. 2001, 70, 442–458. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-X.; Wang, W.-X.; Zhang, Y.-B.; Wang, W.; Lu, S.-L.; Wan, F.-H. Adult diet affects the life history and host-killing behavior of a host-feeding parasitoid. Biol. Control 2015, 81, 58–64. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mele, A.; Scaccini, D.; Pozzebon, A. Hyperparasitism of Acroclisoides sinicus (Huang and Liao) (Hymenoptera: Pteromalidae) on Two Biological Control Agents of Halyomorpha halys. Insects 2021, 12, 617. https://doi.org/10.3390/insects12070617
Mele A, Scaccini D, Pozzebon A. Hyperparasitism of Acroclisoides sinicus (Huang and Liao) (Hymenoptera: Pteromalidae) on Two Biological Control Agents of Halyomorpha halys. Insects. 2021; 12(7):617. https://doi.org/10.3390/insects12070617
Chicago/Turabian StyleMele, Alberto, Davide Scaccini, and Alberto Pozzebon. 2021. "Hyperparasitism of Acroclisoides sinicus (Huang and Liao) (Hymenoptera: Pteromalidae) on Two Biological Control Agents of Halyomorpha halys" Insects 12, no. 7: 617. https://doi.org/10.3390/insects12070617
APA StyleMele, A., Scaccini, D., & Pozzebon, A. (2021). Hyperparasitism of Acroclisoides sinicus (Huang and Liao) (Hymenoptera: Pteromalidae) on Two Biological Control Agents of Halyomorpha halys. Insects, 12(7), 617. https://doi.org/10.3390/insects12070617