
Effect of Soy Leaf Flavonoids on Pea Aphid Probing Behavior


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant and Aphid Cultures
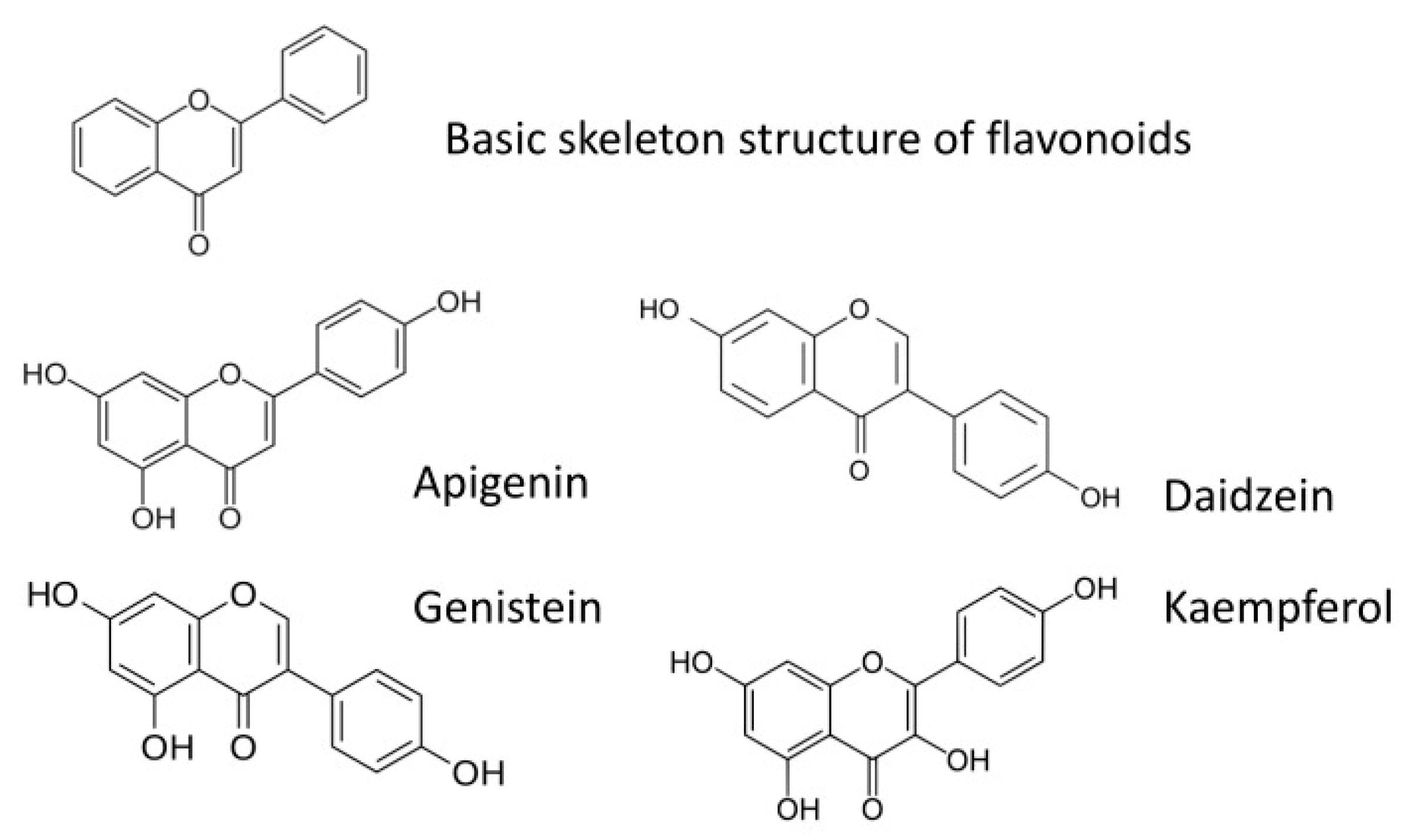
2.2. Application of Flavonoids
2.3. Monitoring of Aphid Probing Behavior (EPG No-Choice Test)
2.4. Statistical Analysis
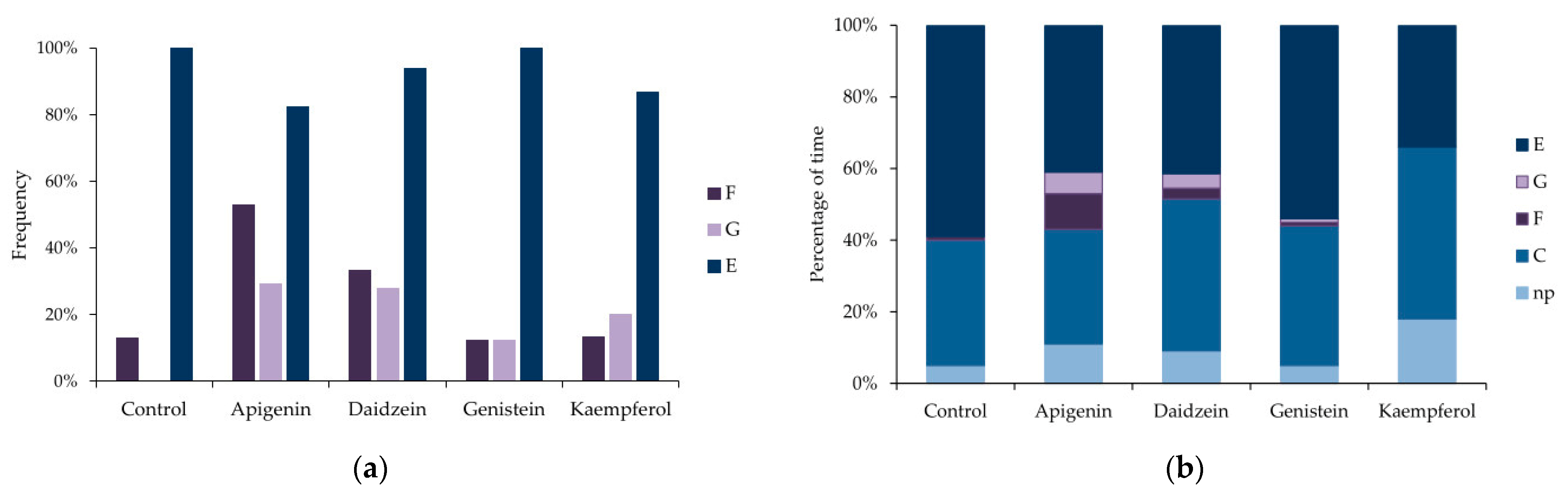
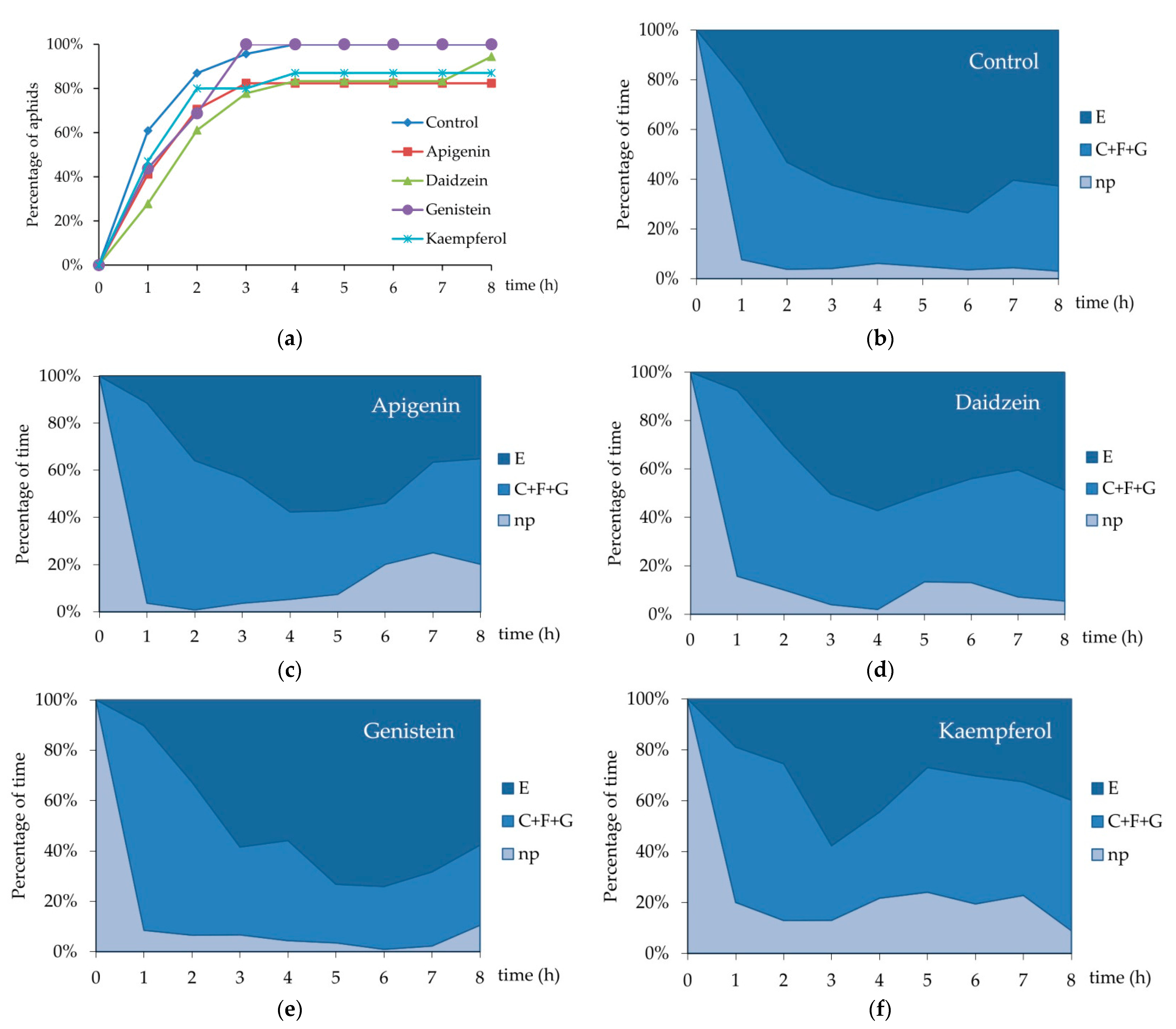
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agati, G.; Azzarellob, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Nakata, R.; Aboshi, T.; Yoshinaga, N.; Teraishi, M.; Okumoto, Y.; Ishihara, A.; Morisaka, H.; Huffaker, A.; Schmelz, E.A.; et al. Insect-induced daidzein, formononetin and their conjugates in soybean leaves. Metabolites 2014, 4, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Stacey, G.; Yu, O. Endogenous isoflavones are essential for the establishment of symbiosis between soybean and Bradyrhizobium japonicum. Plant J. 2006, 48, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Pabich, M.; Materska, M. Biological effect of soy isoflavones in the prevention of civilization diseases. Nutrients 2019, 11, 1660. [Google Scholar] [CrossRef]
- Peiretti, P.G.; Karamać, M.; Janiak, M.; Longato, E.; Meineri, G.; Amarowicz, R.; Gai, F. Phenolic composition and antioxidant activities of soybean (Glycine max (L.) Merr.) plant during growth cycle. Agronomy 2019, 9, 153. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed]
- Bentivenha, J.P.F.; Canassa, V.F.; Baldin, E.L.L.; Borguini, M.L.; Lima, G.P.P.; Lourenção, A.L. Role of the rutin and genistein flavonoids in soybean resistance to Piezodorus guildinii (Hemiptera: Pentatomidae). Arthropod Plant Interact. 2018, 12, 311–320. [Google Scholar] [CrossRef]
- Hoffmann-Campo, C.B.; Harborne, J.B.; McCaffery, A.R. Pre-ingestive and post-ingestive effects of soya bean extracts and rutin on Trichoplusia ni growth. Entomol. Exp. Appl. 2001, 98, 181–194. [Google Scholar] [CrossRef]
- Piubelli, G.C.; Hoffmann-Campo, C.B.; Moscardi, F.; Miyakubo, S.H.; de Oliveira, M.C. Are chemical compounds important for soybean resistance to Anticarsia gemmatalis? J. Chem. Ecol. 2005, 31, 1509–1524. [Google Scholar] [CrossRef]
- Isman, M.B.; Duffey, S.S. Toxicity of tomato phenolic compounds to the fruitworm, Heliothis zea. Entomol. Exp. Appl. 1982, 31, 370–3761. [Google Scholar] [CrossRef]
- Hohenstein, J.D.; Studham, M.E.; Klein, A.; Kovinich, N.; Barry, K.; Lee, Y.-J.; MacIntosh, G.C. Transcriptional and chemical changes in soybean leaves in response to long-term aphid colonization. Front. Plant Sci. 2019, 10, 310. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.V.; Dung, D.M.; Dai, D.N. Accumulation of flavonoids in soybean under effects of cyanobacterial crude extract and aphid infestation. Vietnam. J. Sci. Technol. 2016, 54, 271–277. [Google Scholar] [CrossRef]
- War, A.R.; Sharma, S.P.; Sharma, H.C. Differential induction of flavonoids in groundnut in response to Helicoverpa armigera and Aphis craccivora infestation. Int. J. Insect Sci. 2016, 8, 55–64. [Google Scholar] [CrossRef]
- Mai, V.C.; Drzewiecka, K.; Jeleń, H.; Narożna, D.; Rucińska-Sobkowiak, R.; Kęsy, J.; Floryszak-Wieczorek, J.; Gabryś, B.; Morkunas, I. Differential induction of Pisum sativum defense signaling molecules in response to pea aphid infestation. Plant Sci. 2014, 221–222, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Morkunas, I.; Woźniak, A.; Formela, M.; Mai, V.C.; Marczak, Ł.; Narożna, D.; Borowiak-Sobkowiak, B.; Kühn, C.; Grimm, B. Pea aphid infestation induces changes in flavonoids, antioxidative defence, soluble sugars and sugar transporter expression in leaves of pea seedlings. Protoplasma 2016, 253, 1063–1079. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, A.; Formela, M.; Bilman, P.; Grześkiewicz, K.; Bednarski, W.; Marczak, Ł.; Narożna, D.; Dancewicz, K.; Mai, V.C.; Borowiak-Sobkowiak, B.; et al. The dynamics of the defense strategy of pea induced by exogenous nitric oxide in response to aphid infestation. Int. J. Mol. Sci. 2017, 18, 329. [Google Scholar] [CrossRef] [PubMed]
- Yuan, E.; Yan, H.; Gao, J.; Guo, H.; Ge, F.; Sun, Y. Increases in genistein in Medicago sativa confer resistance against the Pisum host race of Acyrthosiphon pisum. Insects 2019, 10, 97. [Google Scholar] [CrossRef]
- Parrott, W.; Walker, D.; Zhu, S.; Boerma, H.R.; All, J. Genomics of insect-soybean interactions. In Genetics and Genomics of Soybean. Plant Genetics and Genomics: Crops and Models; Stacey, G., Ed.; Springer: New York, NY, USA, 2008; Volume 2, pp. 269–292. [Google Scholar]
- Chang, H.-X.; Hartman, G.L. Characterization of insect resistance loci in the USDA soybean germplasm collection using genome-wide association studies. Front. Plant Sci. 2017, 8, 670. [Google Scholar] [CrossRef]
- Lattanzio, V.; Arpaia, S.; Cardinali, A.; Di Venere, D.; Linsalata, V. Role of endogenous flavonoids in resistance mechanism of Vigna to aphids. J. Agric. Food Chem. 2000, 48, 5316–5320. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V. Phenolic compounds: Introduction. In Natural Products; Ramawat, K.G., Merillon, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1544–1580. [Google Scholar]
- Simmonds, M.S.J. Flavonoid–insect interactions: Recent advances in our knowledge. Phytochemistry 2003, 64, 21–30. [Google Scholar] [CrossRef]
- Hay, W.T.; Behle, R.W.; Berhow, M.A. Biopesticide synergy when combining plant flavonoids and entomopathogenic baculovirus. Sci. Rep. 2020, 10, 6806. [Google Scholar] [CrossRef] [PubMed]
- Frazier, J.L.; Chyb, S. Use of feeding inhibitors in insect control. In Regulatory Mechanisms in Insect Feeding; Chapman, R.F., de Boer, G., Eds.; Springer: Boston, MA, USA, 1995; pp. 364–377. [Google Scholar]
- Grimaldi, D.; Engel, M.S. Evolution of the Insects; Cambridge University Press: New York, NY, USA, 2005; p. 770. [Google Scholar]
- Pettersson, J.; Tjallingii, W.F.; Hardie, J. Host-plant selection and feeding. In Aphids as Crop Pests; van Emden, H.F., Harrington, R., Eds.; CABI: Wallingford, UK, 2017; pp. 173–195. [Google Scholar]
- Tjallingii, W.F.; Hogen Esch, T.H. Fine-structure of aphid stylet routes in plant-tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]
- Agati, G.; Stefano, G.; Biricolti, S.; Tattini, M. Mesophyll distribution of antioxidant flavonoid glycosides in Ligustrum vulgare leaves under contrasting sunlight irradiance. Ann. Bot. 2009, 104, 853–861. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Sieve element acceptance by aphids. Eur. J. Entomol. 1994, 91, 47–52. [Google Scholar]
- Prado, E.; Tjallingii, W.F. Aphid activities during sieve element punctures. Entomol. Exp. Appl. 1994, 72, 157–165. [Google Scholar] [CrossRef]
- Petrussa, E.; Braidot, E.; Zancani, M.; Peresson, C.; Bertolini, A.; Patui, S.; Vianello, A. Plant flavonoids—Biosynthesis, transport and involvement in stress responses. Int. J. Mol. Sci. 2013, 14, 14950–14973. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, B.F.; Zangerl, A.R.; Dermody, O.; Bilgin, D.D.; Casteel, C.L.; Zavala, J.A.; DeLucia, E.H.; Berenbaum, M.R. Impact of elevated levels of atmospheric CO2 and herbivory on flavonoids of soybean (Glycine max Linnaeus). J. Chem. Ecol. 2010, 36, 35–45. [Google Scholar] [CrossRef]
- Zhao, J. Flavonoid transport mechanisms: How to go, and with whom. Trends Plant Sci. 2015, 20, 576–585. [Google Scholar] [CrossRef]
- Goławska, S.; Łukasik, I. Antifeedant activity of luteolin and genistein against the pea aphid, Acyrthosiphon pisum. J. Pest Sci. 2012, 85, 443–450. [Google Scholar] [CrossRef]
- Goławska, S.; Sprawka, I.; Łukasik, I.; Goławski, A. Are naringenin and quercetin useful chemicals in pest-management strategies? J. Pest Sci. 2014, 87, 173–180. [Google Scholar] [CrossRef]
- Goławska, S.; Sprawka, I.; Łukasik, I. Effect of saponins and apigenin mixtures on feeding behavior of the pea aphid, Acyrthosiphon pisum Harris. Biochem. Syst. Ecol. 2014, 55, 137–144. [Google Scholar] [CrossRef]
- Stec, K.; Kordan, B.; Gabrys, B. Quercetin and rutin as modifiers of aphid probing behavior. Molecules 2021, 26, 3622. [Google Scholar] [CrossRef]
- Stec, K.; Kordan, B.; Sergiel, I.; Biesaga, M.; Gasik, J.; Bocianowski, J.; Gabryś, B. Antixenosis in Glycine max (L.) Merr against Acyrthosiphon pisum (Harris). Sci. Rep. 2021, 11, 15289. [Google Scholar] [CrossRef]
- Martin, B.; Collar, L.; Tjallingii, W.F.; Fereres, A. Intracellular ingestion and salivation by aphids may cause the acquisition and inoculation of non-persistently transmitted plant viruses. J. Gen. Virol. 1997, 78, 2701–2705. [Google Scholar] [CrossRef] [PubMed]
- Tjallingii, W.F.; Mayoral, A. Criteria for host-plant acceptance by aphids. In Proceedings of the 8th International Symposium on Insect-Plant Relationships, Wageningen, The Netherlands, 9–13 March 1992; Menken, S.B.J., Visser, J.H., Harrewijn, P., Eds.; Springer, Series Entomologica: Dordrecht, The Netherlands, 1992; Volume 49, pp. 280–282. [Google Scholar]
- Halarewicz-Pacan, A.; Gabryś, B.; Dancewicz, K.; Wawrzeńczyk, C. Enantiospecific effect of limonene and limonene-derived bicyclic lactones on settling and probing behaviour of the peach-potato aphid Myzus persicae (Sulz.). J. Plant Prot. Res. 2003, 43, 133–142. [Google Scholar]
- Paprocka, M.; Gliszczyńska, A.; Dancewicz, K.; Gabryś, B. Novel hydroxy- and epoxy-cis-jasmone and dihydrojasmone derivatives affect the foraging of the peach potato aphid Myzus persicae (Sulzer) (Homoptera: Aphididae). Molecules 2018, 23, 2362. [Google Scholar] [CrossRef] [PubMed]
- Polonsky, J.; Bhatnagar, S.C.; Griffiths, D.C.; Pickett, J.A.; Woodcock, C.M. Activity of qassinoids as antifeedants against aphids. J. Chem. Ecol. 1989, 15, 993–998. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Aphid-plant interactions: What goes on in the depth of the tissue? Proc. Exp. Appl. Entomol. 1995, 6, 163–169. [Google Scholar]
- Backus, E.A.; Cervantes, F.A.; Guedes, R.N.C.; Li, A.Y.; Wayadande, A.C. AC-DC electropenetrography for in-depth studies of feeding and oviposition behaviors. Ann. Entomol. Soc. Am. 2019, 112, 236–248. [Google Scholar] [CrossRef]
- Huang, F.; Tjallingii, W.F.; Zhang, P.; Zhang, J.; Lu, Y.; Lin, J. EPG waveform characteristics of solenopsis mealybug stylet penetration on cotton. Entomol. Exp. Appl. 2012, 143, 47–54. [Google Scholar] [CrossRef]
- van Helden, M.; Tjallingii, W.F. Tissue localization of lettuce resistance to the aphid Nasonovia ribisnigri using electrical penetration graphs. Entomol. Exp. Appl. 1993, 68, 269–278. [Google Scholar] [CrossRef]
- Garzo, E.; Rizzo, E.; Fereres, A.; Gomez, S.K. High levels of arbuscular mycorrhizal fungus colonization on Medicago truncatula reduces plant suitability as a host for pea aphids (Acyrthosiphon pisum). Insect Sci. 2020, 27, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Ten Broeke, C.J.M.; Diecke, M.; van Loon, J. Feeding behaviour and performance of different populations of the black currant-lettuce aphid, Nasonovia ribisnigri, on resistant and susceptible lettuce. Entomol. Exp. Appl. 2013, 148, 130–171. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH; Monograph; Julius Kühn-Institut: Quedlinburg, Germany, 2018. [Google Scholar]
- Joschinski, J.; Beer, K.; Helfrich-Forster, C.; Krauss, J. Pea aphids (Hemiptera: Aphididae) have diurnal rhythms when raised independently of a host plant. J. Insect Sci. 2016, 16, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Schliephake, E.; Habekuss, A.; Scholz, M.; Ordon, F. Barley yellow dwarf virus transmission and feeding behaviour of Rhopalosiphum padi on Hordeum bulbosum clones. Entomol. Exp. Appl. 2013, 146, 347–356. [Google Scholar] [CrossRef]
- Chapman, R.F. Chemosensory regulation of feeding. In Regulatory Mechanisms in Insect Feeding, 1st ed.; Chapman, R.F., de Boer, G., Eds.; Chapman & Hall: New York, NY, USA, 1995; pp. 364–381. [Google Scholar]
- Wensler, R.J.D.; Filshie, B.K. Gustatory sense organs in the food canal of aphids. J. Morphol. 1969, 129, 473–492. [Google Scholar] [CrossRef]
- Powell, G.; Tosh, C.R.; Hardie, J. Host plant selection by aphids: Behavioral, evolutionary, and applied perspectives. Annu. Rev. Entomol. 2006, 51, 309–330. [Google Scholar] [CrossRef]
- Powell, G.; Hardie, J. The chemical ecology of aphid host alternation: How do return migrants find the primary host plant? Appl. Entomol. Zool. 2001, 36, 259–267. [Google Scholar] [CrossRef]
- Prado, E.; Tjallingii, W.F. Behavioral evidence for local reduction of aphid-induced resistance. J. Insect Sci. 2007, 7, 48. [Google Scholar] [CrossRef]
- Pompon, J.; Pelletier, Y. Changes in aphid probing behaviour as a function of insect age and plant resistance level. Bull. Entomol. Res. 2012, 102, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. Symbiotic microorganisms: Untapped resources for insect pest control. Trends Biotechnol. 2007, 25, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. The nutritional physiology of aphids. Adv. Insect Physiol. 2003, 31, 73–140. [Google Scholar]
- Dancewicz, K.; Sznajder, K.; Załuski, D.; Kordan, B.; Gabryś, B. Behavioral sensitivity of Myzus persicae to volatile isporenoids in plant tissues. Entomol. Exp. Appl. 2016, 160, 229–240. [Google Scholar] [CrossRef]
- Powell, G.; Maniar, S.P.; Pickett, J.A.; Hardie, J. Aphid responses to non-host epicuticular lipids. In Proceedings of the 10th International Symposium on Insect-Plant Relationships, Oxford, UK, 4–10 July 1998; Simpson, S.J., Mordue, A.J., Hardie, J., Eds.; Springer, Series Entomologica: Dordrecht, The Netherlands, 1999; Volume 56, pp. 115–123. [Google Scholar]
- Todd, J.C.; Rouf Mian, M.A.; Backus, E.A.; Finer, J.J.; Redinbaugh, M.G. Feeding Behavior of Soybean Aphid (Hemiptera: Aphididae) Biotype 2 on Resistant and Susceptible Soybean. J. Econ. Entomol. 2016, 109, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Baudry, X.; Doury, G.; Couty, A. Antagonist effects of the leek Allium porrum as a companion plant on aphid host plant colonization. Sci. Rep. 2021, 11, 4032. [Google Scholar] [CrossRef] [PubMed]
- Nalam, V.J.; Han, J.; Pitt, W.J.; Acharya, S.R.; Nachappa, P. Location, location, location: Feeding site affects aphid performance by altering access and quality of nutrients. PLoS ONE 2021, 16, 0245380. [Google Scholar] [CrossRef]
- Serrano, M.; Coluccia, F.; Torres, M.; L’Haridon, F.; Métraux, J.P. The cuticle and plant defense to pathogens. Front. Plant Sci. 2014, 5, 274. [Google Scholar] [CrossRef]
- Ziv, C.; Zhao, Z.; Gao, Y.G.; Xia, Y. Multifunctional roles of plant cuticle during plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1088. [Google Scholar] [CrossRef]
- Powell, G.; Hardie, J.; Pickett, J.A. Behavioural evidence for detection of the repellent polygodial by aphid antennal tip sensilla. Physiol. Entomol. 1995, 20, 141–146. [Google Scholar] [CrossRef]
- Hardie, J.; Holyoak, M.; Taylor, N.J.; Griffiths, D.C. The combination of electronic monitoring and video-assisted observations of plant penetration by aphids and behavioural effects of polygodial. Entomol. Exp. Appl. 1992, 62, 233–239. [Google Scholar] [CrossRef]
- Powell, G.; Hardie, J.; Pickett, J.A. Effects of the antifeedant polygodial on plant penetration by aphids, assessed by video and electrical recording. Entomol. Exp. Appl. 1993, 68, 193–200. [Google Scholar] [CrossRef]
- Wróblewska-Kurdyk, A.; Gniłka, R.; Dancewicz, K.; Grudniewska, A.; Wawrzeńczyk, C.; Gabryś, B. β-thujone and its derivatives modify the probing behavior of the peach potato aphid. Molecules 2019, 24, 1847. [Google Scholar] [CrossRef] [PubMed]
- van Hoof, H.A. An Investigation of the Biological Transmission of a Non-Persistent Virus; The Agricultural University: Wageningen, The Netherlands, 1958. [Google Scholar]
- Alvarez, A.E.; Garzo, E.; Verbeek, M.; Vosman, B.; Dicke, M.; Tjallingii, W.F. Infection of potato plants with potato leafroll virus changes attraction and feeding behaviour of Myzus persicae. Entomol. Exp. Appl. 2007, 125, 135–144. [Google Scholar] [CrossRef]
- Gabryś, B.; Tjallingii, W.F.; van Beek, T.A. Analysis of EPG recorded probing by cabbage aphid on host plant parts with different glucosinolate contents. J. Chem. Ecol. 1997, 23, 1661–1673. [Google Scholar] [CrossRef]
- Kordan, B.; Dancewicz, K.; Wróblewska, A.; Gabryś, B. Intraspecific variation in alkaloid profile of four lupine species with implications for the pea aphid probing behaviour. Phytochem. Lett. 2012, 5, 71–77. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Electrical recording of stylet penetration activities. In Aphids, Their Biology, Natural Enemies and Control; Minks, A.K., Harrewijn, P., Eds.; Elsevier: Amsterdam, The Netherlands, 1988; Volume 2B, pp. 95–108. [Google Scholar]
- Marchetti, E.; Civolani, S.; Leis, M.; Chicca, M.; Tjallingii, W.F.; Pasqualini, E.; Baronio, P. Tissue location of resistance in apple to the rosy apple aphid established by electrical penetration graphs. Bull. Insectology 2009, 62, 203–208. [Google Scholar]
- Garzo, E.; Moreno, A.; Plaza, M.; Fereres, A. feeding behavior and virus-transmission ability of insect vectors exposed to systemic insecticides. Plants 2020, 9, 895. [Google Scholar] [CrossRef] [PubMed]
- Wamonje, F.O.; Donnelly, R.; Tungadi, T.D.; Murphy, A.M.; Pate, A.E.; Woodcock, C.; Caulfield, J.; Mutuku, J.M.; Bruce, T.J.A.; Gilligan, C.A.; et al. Different plant viruses induce changes in feeding behavior of specialist and generalist aphids on common bean that are likely to enhance virus transmission. Front. Plant Sci. 2020, 10, 1811. [Google Scholar] [CrossRef]
- Machado-Assefh, C.R.; Alvarez, A.E. Probing behavior of aposymbiotic green peach aphid (Myzus persicae) on susceptible Solanum tuberosum and resistant Solanum stoloniferum plants. Insect Sci. 2018, 25, 127–136. [Google Scholar] [CrossRef]
- He, Y.-Q.; Zhang, Y.-Q.; Chen, J.-N.; Chen, W.-L.; Zeng, X.-Y.; Chen, H.-T.; Ding, W. Effects of Aphidius gifuensis on the feeding behavior and potato virus Y transmission ability of Myzus persicae. Insect Sci. 2018, 25, 1025–1034. [Google Scholar] [CrossRef]
- Will, T.; Vilcinskas, A. The structural sheath protein of aphids is required for phloem feeding. Insect Bioch. Mol. Biol. 2015, 57, 34–40. [Google Scholar] [CrossRef]
- Wróblewska-Kurdyk, A.; Dancewicz, K.; Gliszczyńska, A.; Gabryś, B. New insight into the behaviour modifying activity of two natural sesquiterpenoids farnesol and nerolidol towards Myzus persicae (Sulzer) (Homoptera: Aphididae). Bull. Entomol. Res. 2020, 110, 249–258. [Google Scholar] [CrossRef]
- Chandran, P.; Reese, J.C.; Khan, S.A.; Wang, D.; Schapaugh, W.; Campbell, L.R. Feeding behavior comparison of soybean aphid (Hemiptera: Aphididae) biotypes on different soybean genotypes. J. Econ. Entomol. 2013, 106, 2234–2240. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kordan, B.; Wróblewska-Kurdyk, A.; Bocianowski, J. Variation in susceptibility of rapeseed cultivars to the peach potato aphid. J. Pest Sci. 2021, 94, 435–449. [Google Scholar] [CrossRef]
- Kordan, B.; Stec, K.; Słomiński, P.; Laszczak-Dawid, A.; Wróblewska-Kurdyk, A.; Gabryś, B. Antixenosis potential in pulses against the pea aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2019, 112, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Stec, K.; Kozłowska, J.; Wróblewska-Kurdyk, A.; Kordan, B.; Anioł, M.; Gabryś, B. Effect of naringenin and its derivatives on the probing behavior of Myzus persicae (Sulz.). Molecules 2020, 25, 3185. [Google Scholar] [CrossRef] [PubMed]
- Dancewicz, K.; Szumny, A.; Wawrzeńczyk, C.; Gabryś, B. Repellent and antifeedant activities of citral-derived lactones against the peach potato aphid. Int. J. Mol. Sci. 2020, 21, 8029. [Google Scholar] [CrossRef]
- Spiller, N.J.; Koenders, L.; Tjallingii, W.F. Xylem ingestion by aphids—A strategy for maintaining water balance. Entomol. Exp. Appl. 1990, 55, 101–104. [Google Scholar] [CrossRef]
- Pompon, J.; Quiring, D.; Goyer, C.; Giordanengo, P.; Pelletier, Y. A phloem-sap feeder mixes phloem and xylem sap to regulate osmotic potential. J. Insect Physiol. 2011, 57, 1317–1322. [Google Scholar] [CrossRef]
- Will, T.; van Bel, A.J.E. Physical and chemical interactions between aphids and plants. J. Exp. Bot. 2006, 57, 729–737. [Google Scholar] [CrossRef]
- Gabryś, B.; Pawluk, M. Acceptability of different species of Brassicaceae as hosts for the cabbage aphid. In Proceedings of the 10th International Symposium on Insect-Plant Relationships, Oxford, UK, 4–10 July 1998; Simpson, S.J., Mordue, A.J., Hardie, J., Eds.; Springer, Series Entomologica: Dordrecht, The Netherlands, 1999; Volume 56, pp. 105–109. [Google Scholar]
- Kordan, B.; Stec, K.; Słomiński, P.; Giertych, M.J.; Wróblewska-Kurdyk, A.; Gabryś, B. Susceptibility of forage legumes to infestation by the pea aphid Acyrthosiphon pisum (Harris) (Hemiptera: Aphididae). Crop. Pasture Sci. 2018, 69, 775–784. [Google Scholar] [CrossRef]
- Halarewicz, A.; Gabryś, B. Probing behavior of bird cherry-oat aphid Rhopalosiphum padi (L.) on native bird cherry Prunus padus L. and alien invasive black cherry Prunus serotina Erhr. in Europe and the role of cyanogenic glycosides. Arthropod Plant Interact. 2012, 6, 497–505. [Google Scholar] [CrossRef]
- Grudniewska, A.; Dancewicz, K.; Białońska, A.; Ciunik, Z.; Gabryś, B.; Wawrzeńczyk, C. Synthesis of piperitone-derived halogenated lactones and their effect on aphid probing, feeding, and settling behavior. RSC Adv. 2011, 1, 498–510. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Salivary secretions by aphids interacting with proteins of phloem wound responses. J. Exp. Bot. 2006, 57, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; Tjallingii, W.F.; Thonnessen, A.; van Bel, A.J.E. Molecular sabotage of plant defense by aphid saliva. Proc. Natl. Acad. Sci. USA 2007, 104, 10536–10541. [Google Scholar] [CrossRef] [PubMed]
- Schliephake, E. Aphid resistance in raspberry and feeding behaviour of Amphorophora idaei. J. Plant Dis. Prot. 2010, 117, 60–66. [Google Scholar] [CrossRef]
- Ramírez, C.C.; Niemeyer, H.M. Salivation into sieve elements in relation to plant chemistry: The case of the aphid Sitobion fragariae and the wheat, Triticum aestivum. In Proceedings of the 10th International Symposium on Insect-Plant Relationships, Oxford, UK, 4–10 July 1998; Simpson, S.J., Mordue, A.J., Hardie, J., Eds.; Springer, Series Entomologica: Dordrecht, The Netherlands, 1999; Volume 56, pp. 111–114. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| EPG Variable 1 | Control | Apigenin | Daidzein | Genistein | Kaempferol | |
|---|---|---|---|---|---|---|
| General aspects | n = 23 | n = 17 | n = 18 | n = 16 | n = 15 | |
| Total duration of non-probing 2 | h | 0.4 ± 0.1 ab | 0.9 ± 0.3 a | 0.7 ± 0.2 ab | 0.4 ± 0.1 ab | 1.4 ± 0.4 b |
| Total duration of probing 2 | h | 7.6 ± 0.1 ab | 7.1 ± 0.3 a | 7.3 ± 0.2 ab | 7.6 ± 0.1 ab | 6.6 ± 0.4 b |
| Total duration of pathway (C) 2 | h | 2.8 ± 0.2 a | 2.5 ± 0.3 a | 3.4 ± 0.4 a | 3.1 ± 0.4 a | 3.8 ± 0.4 a |
| Total duration of derailed stylet activities (F) 2 | min | 6.4 ± 3.9 a | 50.1 ± 16.3 a | 16.6 ± 7.1 a | 4.8 ± 3.4 a | 0.1 ± 0.0 a |
| Total duration of xylem phase (G) 2 | min | 0.0 ± 0.0 a | 28.1 ± 15.7 a | 20.9 ± 10.4 a | 6.9 ± 4.7 a | 1.5 ± 0.8 a |
| Total duration of pathway C+F+G 2 | h | 2.9 ± 0.3 a | 3.8 ± 0.3 a | 4.0 ± 0.4 a | 3.3 ± 0.4 a | 3.8 ± 0.4 a |
| Total duration of phloem phase E1+E2 2 | h | 4.7 ± 0.3 a | 3.3 ± 0.5 a | 3.3 ± 0.5 a | 4.3 ± 0.5 a | 2.7 ± 0.6 a |
| Total duration of phloem sap ingestion phase E2 2 | h | 4.6 ± 0.3 a | 3.2 ± 0.5 a | 3.2 ± 0.5 a | 4.3 ± 0.5 a | 2.7 ± 0.6 a |
| Phloem phase index (E1+E2)/(C+E1+E2+G+F) 2 | 0.62 ± 0.04 a | 0.42 ± 0.07 a | 0.43 ± 0.07 a | 0.56 ± 0.06 a | 0.37 ± 0.08 a | |
| Number of probes 2 | # | 16.5 ± 2.0 b | 6.5 ± 0.9 a | 12.4 ± 1.5 ab | 11.1 ± 1.5 ab | 22.8 ± 4.0 b |
| Number of brief probes <180 s 2 | # | 8.4 ± 1.4 b | 1.5 ± 0.4 a | 5.6 ± 1.0 b | 4.8 ± 1.0 ab | 12.1 ± 2.5 b |
| Mean duration of a probe 2 | h | 0.7 ± 0.2 b | 1.7 ± 0.4 a | 0.7 ± 0.1 ab | 0.9 ± 0.1 ab | 0.6 ± 0.2 b |
| Mean duration of np intervals 2 | min | 1.4 ± 0.1 a | 9.3 ± 4.2 ab | 3.2 ± 0.7 ab | 2.5 ± 0.5 ab | 3.5 ± 0.7 b |
| Time to first probe 2 | min | 1.3 ± 0.4 ab | 0.3 ± 0.1 a | 1.8 ± 0.3 b | 1.6 ± 0.7 b | 1.4 ± 0.3 b |
| Duration of first probe 2 | min | 30.6 ± 12.2 a | 80.3 ± 30.5 a | 14.5 ± 8.3 a | 23.3 ± 8.2 a | 13.8 ± 7.5 a |
| Probing in non-phloem tissues before phloem phase | ||||||
| Number of probes before first phloem phase 3 | # | n = 23 4.8 ± 1.0 ab | n = 14 1.7 ± 0.3 b | n = 17 5.6 ± 1.2 a | n = 16 4.2 ± 0.7 ab | n = 13 5.8 ± 2.2 ab |
| Number of brief probes before first phloem phase 3 | # | n = 23 3.2 ± 0.7 b | n = 14 0.7 ± 0.3 a | n = 17 2.6 ± 0.5 b | n = 16 2.3 ± 0.4 ab | n = 13 3.8 ± 1.5 ab |
| Time from first probe to first phloem phase E1 3 | h | n = 23 1.0 ± 0.2 a | n = 14 1.2 ± 0.2 a | n = 17 2.2 ± 0.5 a | n = 16 1.6 ± 0.2 a | n = 13 1.1 ± 0.3 a |
| Time from first probe to first phloem phase E1+E2 3 | h | n = 23 1.0 ± 0.2 b | n = 14 1.3 ± 0.2 ab | n = 17 2.9 ± 0.7 a | n = 16 1.6 ± 0.2 ab | n = 13 1.1 ± 0.3 ab |
| Time from first probe to first sustained sap ingestion phase (E2 > 10 min) 5 | h | n = 23 1.1 ± 0.2 b | n = 14 1.4 ± 0.2 ab | n = 17 2.7 ± 0.6 a | n = 16 1.7 ± 0.2 ab | n = 13 1.4 ± 0.3 ab |
| Probing in phloem tissues | n = 23 | n = 14 | n = 17 | n = 16 | n = 13 | |
| Number of phloem phases E1+E2 3 | # | 4.6 ± 0.5 b | 2.9 ± 0.3 ab | 2.8 ± 0.6 a | 2.5 ± 0.3 ab | 2.8 ± 0.4 ab |
| Mean duration of phloem phase E1 + E2 4 | h | 1.6 ± 0.6 a | 1.8 ± 0.5 a | 2.1 ± 0.4 a | 2.2 ± 0.4 a | 1.4 ± 0.5 a |
| Mean duration of phloem sap ingestion phase E2 4 | h | 1.4 ± 0.4 a | 1.3 ± 0.5 a | 2.1 ± 0.5 a | 2.2 ± 0.4 a | 1.4 ± 0.5 a |
| Phloem salivation index E1/(E1 + E2) 3 | 0.04 ± 0.01 a | 0.03 ± 0.01 a | 0.03 ± 0.01 a | 0.02 ± 0.01 a | 0.05 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stec, K.; Kordan, B.; Gabryś, B. Effect of Soy Leaf Flavonoids on Pea Aphid Probing Behavior. Insects 2021, 12, 756. https://doi.org/10.3390/insects12080756
Stec K, Kordan B, Gabryś B. Effect of Soy Leaf Flavonoids on Pea Aphid Probing Behavior. Insects. 2021; 12(8):756. https://doi.org/10.3390/insects12080756
Chicago/Turabian StyleStec, Katarzyna, Bożena Kordan, and Beata Gabryś. 2021. "Effect of Soy Leaf Flavonoids on Pea Aphid Probing Behavior" Insects 12, no. 8: 756. https://doi.org/10.3390/insects12080756
APA StyleStec, K., Kordan, B., & Gabryś, B. (2021). Effect of Soy Leaf Flavonoids on Pea Aphid Probing Behavior. Insects, 12(8), 756. https://doi.org/10.3390/insects12080756