
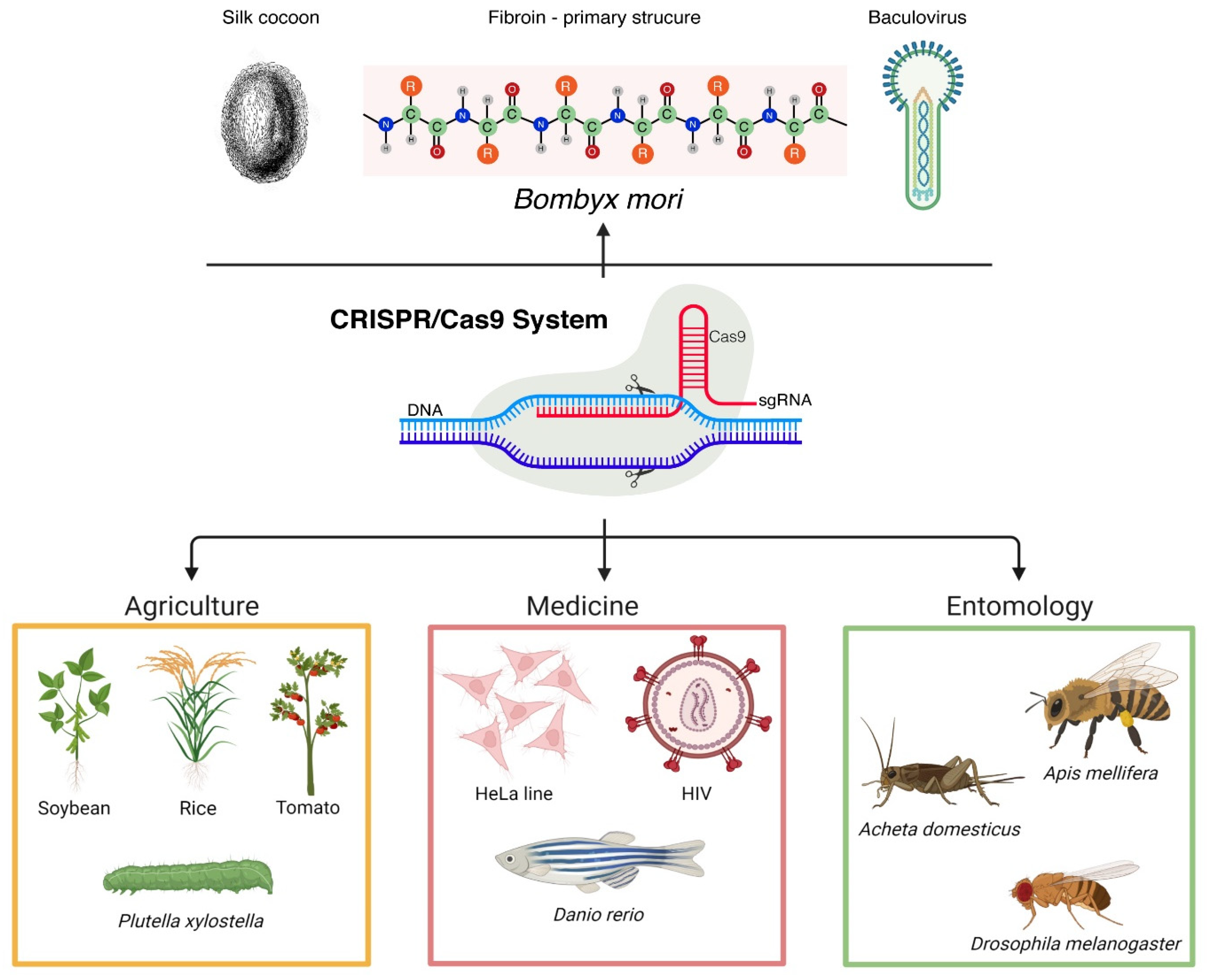
Advances in Editing Silkworms (Bombyx mori) Genome by Using the CRISPR-Cas System

 ,
,  ,
, 
 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. The CRISPR-Cas System
2.1. The CRISPR-Cas Complex Role in the Immunity System
2.2. The CRISPR-Cas System as a Genome Editing Tool
2.3. CRISPR-Cas9 in Entomology
3. Bombyx mori
B. mori as a Model Organism
4. Applications of CRISPR-Cas in B. mori
Applicability of CRISPR-Cas in Anti-BmNPV Therapy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Moon, S.B.; Kim, D.Y.; Ko, J.H.; Kim, Y.S. Recent advances in the CRISPR genome editing tool set. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guha, T.K.; Wai, A.; Hausner, G. Programmable Genome Editing Tools and their Regulation for Efficient Genome Engineering. Comput. Struct. Biotechnol. J. 2017, 15, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Y.; Hong, W.; Huang, M.; Wu, M.; Zhao, X. Applications of genome editing technology in the targeted therapy of human diseases: Mechanisms, advances and prospects. Signal Transduct. Target. Ther. 2020, 5, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Schuijff, M.; De Jong, M.D.T.; Dijkstra, A.M. AQ methodology study on divergent perspectives on CRISPR-Cas9 in the Netherlands. BMC Med. Ethics 2021, 22, 48. [Google Scholar] [CrossRef]
- Zhang, D.; Hussain, A.; Manghwar, H.; Xie, K.; Xie, S.; Zhao, S.; Larkin, R.M.; Qing, P.; Jin, S.; Ding, F. Genome editing with the CRISPR-Cas system: An art, ethics and global regulatory perspective. Plant Biotechnol. J. 2020, 18, 1651–1669. [Google Scholar] [CrossRef]
- Li, P.; Wang, L.; Yang, J.; Di, L.; Li, J. Applications of the CRISPR-Cas system for infectious disease diagnostics. Expert Rev. Mol. Diagn. 2021, 21, 723–732. [Google Scholar] [CrossRef]
- Sezutsu, H.; Sumitani, M.; Kondo, M.; Kobayashi, I.; Takasu, Y.; Suzuki, T.; Yonemura, N.; Iizuka, T.; Uchino, K.; Tamura, T.; et al. Construction of a Platform for the Development of Pharmaceutical and Medical Applications Using Transgenic Silkworms. Yakugaku Zasshi J. Pharm. Soc. Jpn. 2018, 138, 863–874. [Google Scholar] [CrossRef]
- Kurihara, H.; Sezutsu, H.; Tamura, T.; Yamada, K. Production of an active feline interferon in the cocoon of transgenic silkworms using the fibroin H-chain expression system. Biochem. Biophys. Res. Commun. 2007, 355, 976–980. [Google Scholar] [CrossRef]
- Nakaya, H.; Tatematsu, K.I.; Sezutsu, H.; Kuwabara, N.; Koibuchi, N.; Takeda, S. Secretory expression of thyroid hormone receptor using transgenic silkworms and its DNA binding activity. Protein Expr. Purif. 2020, 176, 105723. [Google Scholar] [CrossRef]
- Itoh, K.; Kobayashi, I.; Nishioka, S.; Hidaka, T.; Tsuji, D.; Sezutsu, H. A transgenic silkworm overexpressing human lysosomal enzyme as a novel resource for producing recombinant glycobiologics and its application to development of enzyme replacement therapy for lysosomal diseases. Mol. Genet. Metab. 2016, 120, S69. [Google Scholar] [CrossRef]
- Xu, H.; O’Brochta, D.A. Advanced technologies for genetically manipulating the silkworm Bombyx mori, a model lepidopteran insect. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150487. [Google Scholar] [CrossRef] [Green Version]
- Montali, A.; Romanelli, D.; Cappellozza, S.; Grimaldi, A.; De Eguileor, M.; Tettamanti, G. Timing of autophagy and apoptosis during posterior silk gland degeneration in Bombyx mori. Arthropod Struct. Dev. 2017, 46, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.Y.; Smagghe, G.; Xia, Q.Y. Genome editing in Bombyx mori: New opportunities for silkworm functional genomics and the sericulture industry. Insect Sci. 2018, 26, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA—Guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Nidhi, S.; Anand, U.; Oleksak, P.; Tripathi, P.; Lal, J.A.; Thomas, G.; Kuca, K.; Tripathi, V. Novel CRISPR—Cas Systems: An Updated Review of the Current Achievements, Applications, and Future Research Perspectives. Int. J. Mol. Sci. 2021, 22, 3327. [Google Scholar] [CrossRef] [PubMed]
- Hahn, F.; Loures, L.S.; Sparks, C.A.; Kanyuka, K.; Nekrasov, V. Efficient CRISPR/Cas-Mediated Targeted Mutagenesis in Spring and Winter Wheat Varieties. Plants 2021, 10, 1481. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Yoosefzadeh Najafabadi, M.; Adamek, K.; Torkamaneh, D.; Jones, A.M.P. Synergizing off-target predictions for in silico insights of CENH3 Knockout in Cannabis through CRISPR/Cas. Molecules 2021, 26, 2053. [Google Scholar] [CrossRef]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide Sequence of the iap Gene, Responsible for Alkaline Phosphatase Isozyme Conversion in Escherichia coli, and Identification of the Gene Product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [Green Version]
- Gophna, U.; Brodt, A. CRISPR/Cas systems in archaea. Mob. Genet. Elem. 2012, 2, 63–64. [Google Scholar] [CrossRef] [Green Version]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the immune system of Bacteria and Archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Shabbir, M.A.B.; Shabbir, M.Z.; Wu, Q.; Mahmood, S.; Sajid, A.; Maan, M.K.; Ahmed, S.; Naveed, U.; Hao, H.; Yuan, Z. CRISPR-cas system: Biological function in microbes and its use to treat antimicrobial resistant pathogens. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 21. [Google Scholar] [CrossRef] [PubMed]
- Rath, D.; Amlinger, L.; Rath, A.; Lundgren, M. The CRISPR-Cas immune system: Biology, mechanisms and applications. Biochimie 2015, 117, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Faure, G.; Shmakov, S.A.; Yan, W.X.; Cheng, D.R.; Scott, D.A.; Peters, J.E.; Makarova, K.S.; Koonin, E.V. CRISPR–Cas in mobile genetic elements: Counter-defence and beyond. Nat. Rev. Microbiol. 2019, 17, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Mcdonald, N.D.; Regmi, A.; Morreale, D.P.; Borowski, J.D.; Boyd, E.F. CRISPR-Cas systems are present predominantly on mobile genetic elements in Vibrio species. BMC Genom. 2019, 20, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haft, D.H.; Selengut, J.; Mongodin, E.F.; Nelson, K.E. A Guild of 45 CRISPR-Associated (Cas) Protein Families and Multiple CRISPR/Cas Subtypes Exist in Prokaryotic Genomes. PLoS Comput. Biol. 2005, 1, e60. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Horvath, P.; Moineau, S.; Mojica, F.J.M.; Wolf, Y.I.; Yakunin, A.F.; et al. Evolution and classification of the CRISPR-Cas systems. Nat. Rev. Microbiol. 2011, 9, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Koonin, E.V. Annotation and Classification of CRISPR-Cas Systems. In CRISPR; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–27. [Google Scholar] [CrossRef]
- Koonin, E.V.; Makarova, K.S. Origins and evolution of CRISPR-Cas systems. Philos. Trans. R. Soc. B 2019, 374, 20180087. [Google Scholar] [CrossRef]
- Alkhnbashi, O.S.; Shah, S.A.; Garrett, R.A.; Saunders, S.J.; Costa, F.; Backofen, R. Characterizing leader sequences of CRISPR loci. Bioinformatics 2016, 32, i576–i585. [Google Scholar] [CrossRef] [Green Version]
- Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Garrett, R.A.; Saunders, S.J.; Backofen, R. CRISPRstrand: Predicting repeat orientations to determine the crRNA-encoding strand at CRISPR loci. Bioinformatics 2014, 30, i489–i496. [Google Scholar] [CrossRef] [Green Version]
- McGinn, J.; Marraffini, L.A. Molecular mechanisms of CRISPR–Cas spacer acquisition. Nat. Rev. Microbiol. 2018, 17, 7–12. [Google Scholar] [CrossRef]
- Sorek, R.; Lawrence, C.M.; Wiedenheft, B. CRISPR-Mediated Adaptive Immune Systems in Bacteria and Archaea. Annu. Rev. Biochem. 2013, 82, 237–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.; Barrangou, R. Applications of CRISPR-Cas systems in lactic acid bacteria. FEMS Microbio. Rev. 2020, 44, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Hryhorowicz, M.; Lipiński, D.; Zeyland, J.; Słomski, R. CRISPR/Cas9 Immune System as a Tool for Genome Engineering. Arch. Immunol. Ther. Exp. 2016, 65, 233–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terns, M.P.; Terns, R.M. CRISPR-based adaptive immune systems. Curr. Opin. Microbiol. 2011, 14, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newsom, S.; Parameshwaran, H.P.; Martin, L.; Rajan, R. The CRISPR-Cas Mechanism for Adaptive Immunity and Alternate Bacterial Functions Fuels Diverse Biotechnologies. Front. Cell. Infect. Microbiol. 2021, 10, 1–10. [Google Scholar] [CrossRef]
- Marraffini, L.A.; Sontheimer, E.J. CRISPR interference: RNA-directed adaptive immunity in bacteria and archaea. Nat. Rev. Genet. 2010, 11, 181–190. [Google Scholar] [CrossRef]
- Ishino, Y.; Krupovic, M.; Forterre, P. History of CRISPR-Cas from Encounter with a Mysterious Repeated Sequence to Genome Editing Technology. J. Bacteriol. 2018, 200, e00580-17. [Google Scholar] [CrossRef] [Green Version]
- Sontheimer, E.J.; Barrangou, R. The Bacterial Origins of the CRISPR Genome-Editing Revolution. Hum. Gene Ther. 2015, 26, 413–424. [Google Scholar] [CrossRef]
- El-Mounadi, K.; Morales-Floriano, M.L.; Garcia-Ruiz, H. Principles, Applications, and Biosafety of Plant Genome Editing Using CRISPR-Cas9. Front. Plant Sci. 2020, 11, 56. [Google Scholar] [CrossRef]
- Agustın-Pavon, C.; Isalan, M. Synthetic biology and therapeutic strategies for the degenerating brain. Bioessays 2014, 36, 979–990. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Tang, L.; Xiang, H.; Jin, L.; Li, Q.; Dong, Y.; Wang, W.; Zhang, G. Advances in genome editing technology and its promising application in evolutionary and ecological studies. Gigascience 2014, 3, 2047-217X. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavakoli, K.; Pour-Aboughadareh, A.; Kianersi, F.; Poczai, P.; Etminan, A.; Shooshtari, L. Applications of CRISPR-Cas9 as an Advanced Genome Editing System in Life Sciences. BioTech 2021, 10, 14. [Google Scholar] [CrossRef]
- Guha, T.K.; Edgell, D.R. Applications of Alternative Nucleases in the Age of CRISPR/Cas9. Int. J. Mol. Sci. 2017, 18, 2565. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.H. Genome-Editing Technologies: Concept, Pros, and Cons of Various Genome-Editing Techniques and Bioethical Concerns for Clinical Application. Mol. Ther. Nucleic Acids 2019, 16, 326–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chira, S.; Gulei, D.; Hajitou, A.; Zimta, A.A.; Cordelier, P.; Berindan-Neagoe, I. CRISPR/Cas9: Transcending the Reality of Genome Editing. Mol. Ther. Nucleic Acids 2017, 7, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Manghwar, H.; Lindsey, K.; Zhang, X.; Jin, S. CRISPR/Cas System: Recent Advances and Future Prospects for Genome Editing. Trends Plant Sci. 2019, 24, 1102–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Shi, Y.; Wu, J.; Li, H.; Smagghe, G.; Liu, T. CRISPR/Cas9 in lepidopteran insects: Progress, application and prospects. J. Insect Physiol. 2021, 135, 104325. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Kumar, R.; Das, A.; Won, S.Y.; Shukla, P. CRISPR-Cas9 system: A genome-editing tool with endless possibilities. J. Biotechnol. 2020, 319, 36–53. [Google Scholar] [CrossRef]
- Zhang, Y.; Showalter, A.M. CRISPR/Cas9 Genome Editing Technology: A Valuable Tool for Understanding Plant Cell Wall Biosynthesis and Function. Front. Plant Sci. 2020, 11, 589517. [Google Scholar] [CrossRef] [PubMed]
- Collias, D.; Beisel, C.L. CRISPR technologies and the search for the PAM-free nuclease. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ren, S.; Yu, S.; Pan, H.; Li, T.; Ge, S.; Zhang, J.; Xia, N. Methods Favoring Homology-Directed Repair Choice in Response to CRISPR/Cas9 Induced-Double Strand Breaks. Int. J. Mol. Sci. 2020, 21, 6461. [Google Scholar] [CrossRef]
- Bernheim, A.; Calvo-villamañán, A.; Basier, C.; Cui, L.; Rocha, E.; Touchon, M.; Bikard, D. Inhibition of NHEJ repair by type II-A CRISPR-Cas systems in bacteria. Nat. Commun. 2017, 8, 25–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Stazio, M.; Foschi, N.; Athanasakis, E.; Gasparini, P.; d’Adamo, A.P. Systematic analysis of factors that improve homologous direct repair (HDR) efficiency in CRISPR/Cas9 technique. PLoS ONE 2021, 16, e0247603. [Google Scholar] [CrossRef]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Douris, V.; Denecke, S.; Van Leeuwen, T.; Bass, C.; Nauen, R.; Vontas, J. Using CRISPR/Cas9 genome modification to understand the genetic basis of insecticide resistance: Drosophila and beyond. Pestic. Biochem. Physiol. 2020, 167, 104595. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Kesiraju, K.; Saakre, M.; Rathinam, M.; Raman, V.; Pattanayak, D.; Sreevathsa, R. Genome Editing for Resistance to Insect Pests: An Emerging Tool for Crop Improvement. ACS Omega 2020, 5, 20674–20683. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, J.; Ge, S.; Lai, L. CRISPR/Cas: Advances, Limitations, and Applications for Precision Cancer Research. Front. Med. 2021, 8, 649896. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, X.; Li, S.; Chen, Y.; Zhao, Y.; Wei, Y.; Qiu, Y.; Liu, Y.; Zhou, Z.; Han, J.; et al. Genome-scale CRISPR screening for potential targets of ginsenoside compound K. Cell Death Dis. 2020, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Li, Z. CRISPR-Cas systems: Overview, innovations and applications in human disease research and gene therapy. Comput. Struct. Biotechnol. J. 2020, 18, 2401–2415. [Google Scholar] [CrossRef]
- Ma, X.; He, K.; Shi, Z.; Li, M.; Li, F.; Chen, X.-X. Large-Scale Annotation and Evolution Analysis of MiRNA in Insects. Genome Biol. Evol. 2021, 13, evab083. [Google Scholar] [CrossRef] [PubMed]
- Brady, D.; Grapputo, A.; Romoli, O.; Sandrelli, F. Insect Cecropins, Antimicrobial Peptides with Potential Therapeutic Applications. Int. J. Mol. Sci. 2019, 20, 5862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romoli, O.; Mukherjee, S.; Mohid, S.A.; Dutta, A.; Montali, A.; Franzolin, E.; Brady, D.; Zito, F.; Bergantino, E.; Rampazzo, C.; et al. Enhanced Silkworm Cecropin B Antimicrobial Activity against Pseudomonas aeruginosa from Single Amino Acid Variation. ACS Infect. Dis. 2019, 5, 1200–1213. [Google Scholar] [CrossRef]
- Cui, Y.; Sun, J.; Yu, L. Application of the CRISPR gene-editing technique in insect functional genome studies—A review. Entomol. Exp. Appl. 2017, 162, 124–132. [Google Scholar] [CrossRef] [Green Version]
- De Lazzari, F.; Sandrelli, F.; Whitworth, A.J.; Bisaglia, M. Antioxidant Therapy in Parkinson’s Disease: Insights from Drosophila melanogaster. Antioxidants 2020, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Taning, C.N.T.; Van Eynde, B.; Yu, N.; Ma, S.; Smagghe, G. CRISPR/Cas9 in insects: Applications, best practices and biosafety concerns. J. Insect Physiol. 2017, 98, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.F.; Zhang, B.; Liao, C.H.; Zeng, Z.J. High-Efficiency CRISPR/Cas9-Mediated Gene Editing in Honeybee (Apis mellifera) Embryos. G3 Genes Genomes Genet. 2019, 9, 1759–1766. [Google Scholar] [CrossRef] [Green Version]
- Gratz, S.J.; Cummings, A.M.; Nguyen, J.N.; Hamm, D.C.; Donohue, L.K.; Harrison, M.M.; Wildonger, J.; O’Connor-giles, K.M. Genome Engineering of Drosophila with the CRISPR RNA-guided Cas9 nuclease. Genetics 2013, 194, 1029–1035. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Ren, M.; Wang, Z.; Zhang, B.; Rong, Y.S.; Jiao, R.; Gao, G. Highly Efficient Genome Modifications Mediated by CRISPR/Cas9 in Drosophila. Genetics 2013, 195, 289–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, T.; Batterham, P. Harnessing model organisms to study insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 61–67. [Google Scholar] [CrossRef]
- Homem, R.A.; Davies, T.G.E. An overview of functional genomic tools in deciphering insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 103–110. [Google Scholar] [CrossRef]
- Scott, J.G.; Buchon, N. Drosophila melanogaster as a powerful tool for studying insect toxicology. Pestic. Biochem. Physiol. 2019, 161, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Nakamura, T.; Watanabe, T.; Barnett, A.A.; Noji, S.; Mito, T.; Extavour, C.G. Establishment of CRISPR/Cas9-based knock-in in a hemimetabolous insect: Targeted gene tagging in the cricket Gryllus bimaculatus. bioRxiv 2021. [Google Scholar] [CrossRef]
- Donoughe, S.; Extavour, C.G. Embryonic development of the cricket Gryllus bimaculatus. Dev. Biol. 2016, 411, 140–156. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cucu, A.A.; Baci, G.M.; Moise, A.R.; Dezsi, S.; Marc, B.D.; Stângaciu, S.; Dezmiream, D.S. Towards a Better Understanding of Nutritional and Therapeutic Effects of Honey and Their Applications in Apitherapy. Appl. Sci. 2021, 11, 4190. [Google Scholar] [CrossRef]
- Baci, G.; Cucu, A.; Moise, A.R.; Dezmirean, D.S. Applicability of Honey on Silkworms (Bombyx mori) and Quality Improvement of Its Biomaterials. Appl. Sci. 2021, 11, 4613. [Google Scholar] [CrossRef]
- Kohno, H.; Suenami, S.; Takeuchi, H.; Sasaki, T.; Kubo, T. Production of Knockout Mutants by CRISPR/Cas9 in the European Honeybee, Apis mellifera L. Zool. Sci. 2016, 33, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Liang, L.; Li, Q.; Li, Z.; Zhu, Y.; Guo, Y.; Zheng, Q.; Lin, Y.; Yang, D.; Li, Z.; et al. CRISPR/Cas9 mediated knockout of Amyellow-y gene results in melanization defect of the cuticle in adult Apis mellifera. J. Insect Physiol. 2021, 132, 104264. [Google Scholar] [CrossRef] [PubMed]
- Gempe, T.; Hasselmann, M.; Schiøtt, M.; Hause, G.; Otte, M.; Beye, M. Sex Determination in Honeybees: Two Separate Mechanisms Induce and Maintain the Female Pathway. PLoS Biol. 2009, 7, e1000222. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lin, Y.; Liang, L.; Geng, H.; Zhang, M.; Nie, H.; Su, S. Transcriptional Profiles of Diploid Mutant Apis mellifera Embryos after Knockout of csd by CRISPR/Cas9. Insects 2021, 12, 704. [Google Scholar] [CrossRef]
- Horie, Y.; Watanabe, H. Recent advances in sericulture. Annu. Rev. Entomol. 1980, 25, 49–71. [Google Scholar] [CrossRef]
- Saviane, A.; Romoli, O.; Bozzato, A.; Freddi, G.; Cappelletti, C.; Rosini, E.; Cappellozza, S.; Tettamanti, G.; Sandrelli, F. Intrinsic antimicrobial properties of silk spun by genetically modified silkworm strains. Transgenic Res. 2018, 27, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Dong, Q.; Yu, Y.; Niu, B.; Ji, D.; Li, M.; Huang, Y.; Chen, X.; Tan, A. Mass spider silk production through targeted gene replacement in Bombyx mori. Proc. Natl. Acad. Sci. USA 2018, 115, 8757–8762. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Ye, X.; Wang, X.; Zhao, S.; Wu, M.; Ruan, J.; Zhong, B. High mechanical property silk produced by transgenic silkworms expressing the spidroins PySp1 and ASG1. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Leem, J.W.; Fraser, M.J.; Kim, Y.L. Transgenic and Diet-Enhanced Silk Production for Reinforced Biomaterials: A Metamaterial Perspective. Annu. Rev. Biomed. Eng. 2020, 22, 79–102. [Google Scholar] [CrossRef] [Green Version]
- Cappellozza, L.; Cappellozza, S.; Saviane, A.; Sbrenna, G. Artificial diet rearing system for the silkworm Bombyx mori (Lepidoptera: Bombycidae): Effect of vitamin C deprivation on larval growth and cocoon production. Appl. Entomol. Zool. 2005, 40, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Marrazzo, P.; Crupi, A.N.; Alviano, F.; Teodori, L.; Bonsi, L. Exploring the roles of MSCs in infections: Focus on bacterial diseases. J. Mol. Med. 2019, 97, 437–450. [Google Scholar] [CrossRef]
- Abdelli, N.; Peng, L.; Keping, C. Silkworm, Bombyx mori, as an alternative model organism in toxicological research. Environ. Sci. Pollut. Res. 2018, 25, 35048–35054. [Google Scholar] [CrossRef]
- Meng, X.; Zhu, F.; Chen, K. Silkworm: A promising model organism in life science. J. Insect Sci. 2017, 17. [Google Scholar] [CrossRef]
- Yokoi, K.; Tsubota, T.; Jouraku, A.; Sezutsu, H.; Bono, H. Reference Transcriptome Data in Silkworm Bombyx mori. Insects 2021, 12, 519. [Google Scholar] [CrossRef]
- Hamamoto, H.; Tonoike, A.; Narushima, K.; Horie, R.; Sekimizu, K. Silkworm as a model animal to evaluate drug candidate toxicity and metabolism. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 334–339. [Google Scholar] [CrossRef]
- Hamamoto, H.; Kurokawa, K.; Kaito, C.; Kamura, K.; Razanajatovo, I.M.; Kusuhara, H.; Santa, T.; Sekimizu, K. Quantitative Evaluation of the Therapeutic Effects of Antibiotics Using Silkworms Infected with Human Pathogenic Microorganisms. Antimicrob. Agents Chemother. 2004, 48, 774–779. [Google Scholar] [CrossRef] [Green Version]
- Kaito, C.; Sekimizu, K. A silkworm model of pathogenic bacterial infection. Drug Discov. Ther. 2007, 1, 89–93. [Google Scholar]
- Montali, A.; Berini, F.; Brivio, M.F.; Mastore, M.; Saviane, A.; Cappellozza, S.; Marinelli, F.; Tettamanti, G. A Silkworm Infection Model for In Vivo Study of Glycopeptide Antibiotics. Antibiotics 2020, 9, 300. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Tateyama, Y.; Sugita, T. Evaluation of Antibacterial Drugs Using Silkworms Infected by Cutibacterium acnes. Insects 2021, 12, 619. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Ishii, M.; Ishii, K.; Miyaguchi, W.; Horie, R.; Inagaki, Y.; Hamamoto, H.; Tatematsu, K.-I.; Uchino, K.; Tamura, T.; et al. Transgenic silkworms expressing human insulin receptors for evaluation of therapeutically active insulin receptor agonists. Biochem. Biophys. Res. Commun. 2014, 455, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Ishii, M.; Hayashi, Y.; Miyazaki, S.; Sugita, T.; Sumiya, E.; Sekimizu, K. Diabetic silkworms for evaluation of therapeutically effective drugs against type II diabetes. Sci. Rep. 2015, 5, 10722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, Y.; Suzuki, J.; Watanabe, K.; Shimizu, T.; Watarai, M. Effect of Vitamin A on Listeria monocytogenes Infection in a Silkworm Model. PLoS ONE 2016, 11, e0163747. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Kurakado, S.; Sugita, T. Evaluating Candida albicans biofilm formation in silkworms. Med. Mycol. 2021, 59, 201–205. [Google Scholar] [CrossRef]
- Jiang, G.; Song, J.; Hu, H.; Tong, X.; Dai, F. Evaluation of the silkworm lemon mutant as an invertebrate animal model for human sepiapterin reductase deficiency. R. Soc. Open Sci. 2020, 7, 191888. [Google Scholar] [CrossRef]
- Wang, L.; Su, M.; Zhao, X.; Hong, J.; Yu, X.; Xu, B.; Sheng, L.; Liu, N.; Shen, W.; Li, B.; et al. Nanoparticulate TiO 2 Protection of Midgut Damage in the Silkworm (Bombyx mori) Following Phoxim Exposure. Arch. Environ. Contam. Toxicol. 2015, 68, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Zhao, X.; Wang, Q.; Niu, B.; Liu, Y.; He, L.; Weng, H.; Meng, Z.; Chen, Y. Genotoxicity evaluation of low doses of avermectin to hemocytes of silkworm (Bombyx mori) and response of gene expression to DNA damage. Pestic. Biochem. Physiol. 2011, 101, 159–164. [Google Scholar] [CrossRef]
- Vyjayanthi, N.; Subramanyam, M.V.V. Effect of Fenvalerate-20EC on Sericigenous Insects: II. Digestive Enzymes in the Nutritive Physiology of Silkworm, Bombyx mori L. Ecotoxicol. Environ. Saf. 2002, 53, 212–220. [Google Scholar] [CrossRef]
- Yasu, T.; Matsumoto, Y.; Sugita, T. Pharmacokinetics of voriconazole and its alteration by Candida albicans infection in silkworms. J. Antibiot. 2021, 74, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, Y.; Matsumoto, Y.; Kataoka, K.; Matsuhashi, N.; Sekimizu, K. Evaluation of drug-induced tissue injury by measuring alanine aminotransferase (ALT) activity in silkworm hemolymph. BMC Pharmacol. Toxicol. 2012, 13, 13. [Google Scholar] [CrossRef] [Green Version]
- Pandiarajan, J.; Jeyarani, V.; Balaji, S.; Krishnan, M. Silver Nanoparticles an Accumulative Hazard in Silkworm: Bombyx mori. Austin J. Biotechnol. Bioeng. 2016, 3, 1057. [Google Scholar]
- Gad, A.A. Toxicity effect of Silver Nanoparticles to the Haemocytes and Antioxidant activity of Silkworm Bombyx mori. Physiol. Entomol. 2020, 45, 154–160. [Google Scholar] [CrossRef]
- Meng, X.; Abdlli, N.; Wang, N.; Lü, P.; Nie, Z.; Dong, X.; Lu, S.; Chen, K. Effects of Ag Nanoparticles on Growth and Fat Body Proteins in Silkworms (Bombyx mori). Biol. Trace Elem. Res. 2017, 180, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Nouara, A.; Lü, P.; Chen, L.; Pan, Y.; Yang, Y.; Chen, K. Silver effects on silkworm, Bombyx mori. J. Toxicol. Sci. 2018, 43, 697–709. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, W.; Ma, L.; Cui, X.; Lynch, I.; Wu, G. Acute toxicity of Zinc Oxide nanoparticles to silkworm (Bombyx mori L.). Chemosphere 2020, 259, 127481. [Google Scholar] [CrossRef]
- Li, K.L.; Zhang, Y.H.; Xing, R.; Zhou, Y.F.; Chen, X.D.; Wang, H.; Song, B.; Sima, Y.H.; Xu, S.Q. Different toxicity of cadmium telluride, silicon, and carbon nanomaterials against hemocytes in silkworm, Bombyx mori. RSC Adv. 2017, 7, 50317–50327. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, Z.; Xu, J.; Zeng, B.; Ling, L.; You, L.; Chen, Y.; Huang, Y.; Tan, A. The CRISPR/Cas System mediates efficient genome engineering in Bombyx mori. Cell Res. 2013, 23, 1414–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, T.; Daimon, T.; Uchino, K.; Banno, Y.; Katsuma, S.; Sezutsu, H.; Tamura, T.; Shimada, T. Transgenic analysis of the BmBLOS2 gene that governs the translucency of the larval integument of the silkworm, Bombyx mori. Insect Mol. Biol. 2010, 19, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, S.; Wang, X.; Chang, J.; Gao, J.; Shi, R.; Zhang, J.; Lu, W.; Liu, Y.; Zhao, P.; et al. Highly efficient multiplex targeted mutagenesis and genomic structure variation in Bombyx mori cells using CRISPR/Cas9. Insect Biochem. Mol. Biol. 2014, 49, 35–42. [Google Scholar] [CrossRef]
- Sakuma, T.; Nishikawa, A.; Kume, S.; Chayama, K.; Yamamoto, T. Multiplex genome engineering in human cells using all-in-one CRISPR/Cas9 vector system. Sci. Rep. 2014, 4, 5400. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Chang, J.; Wang, X.; Liu, Y.; Zhang, J.; Lu, W.; Gao, J.; Shi, R.; Zhao, P.; Xia, Q. CRISPR/Cas9 mediated multiplex genome editing and heritable mutagenesis of BmKu70 in Bombyx mori. Sci. Rep. 2014, 4, 4489. [Google Scholar] [CrossRef] [Green Version]
- Fujinaga, D.; Kohmura, Y.; Okamoto, N.; Kataoka, H.; Mizoguchi, A. Insulin-like growth factor (IGF)-like peptide and 20-hydroxyecdysone regulate the growth and development of the male genital disk through different mechanisms in the silkmoth, Bombyx mori. Insect Biochem. Mol. Biol. 2017, 87, 35–44. [Google Scholar] [CrossRef]
- Fujinaga, D.; Shiomi, K.; Yagi, Y.; Kataoka, H.; Mizoguchi, A. An insulin-like growth factor-like peptide promotes ovarian development in the silkmoth Bombyx mori. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Wang, X.; Ma, S.; Peng, Z.; Cao, Y.; Xia, Q. CRISPR-Mediated Endogenous Activation of Fibroin Heavy Chain Gene Triggers Cellular Stress Responses in Bombyx mori Embryonic Cells. Insects 2021, 12, 552. [Google Scholar] [CrossRef]
- Cui, Y.; Zhu, Y.; Lin, Y.; Chen, L.; Feng, Q.; Wang, W.; Xiang, H. New insight into the mechanism underlying the silk gland biological process by knocking out fibroin heavy chain in the silkworm. BMC Genom. 2018, 19, 215. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xia, L.; Day, B.A.; Harris, T.I.; Oliveira, P.; Knittel, C.; Licon, A.L.; Gong, C.; Dion, G.; Lewis, R.V.; et al. CRISPR/Cas9 Initiated Transgenic Silkworms as a Natural Spinner of Spider Silk. Biomacromolecules 2019, 20, 2252–2264. [Google Scholar] [CrossRef] [PubMed]
- Michlewski, G.; Cáceres, J.F. Post-transcriptional control of miRNA biogenesis. RNA 2019, 25, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, L.; Ge, X.; Li, Z.; Zeng, B.; Xu, J.; Chen, X.; Shang, P.; James, A.A.; Huang, Y.; Tan, A. MiR-2 family targets awd and fng to regulate wing morphogenesis in Bombyx mori. RNA Biol. 2015, 12, 742–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Xu, J.; Ling, L.; Luo, X.; Yang, D.; Yang, X.; Zhang, X.; Huang, Y. miR-34 regulates larval growth and wing morphogenesis by directly modulating ecdysone signalling and cuticle protein in Bombyx mori. RNA Biol. 2020, 17, 1342–1351. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Tanaka, H.; Ishibashi, J.; Imanishi, S.; Yamakawa, M. Functional Characterization of a Cactus Homolog from the Silkworm Bombyx mori. Biosci. Biotechnol. Biochem. 2009, 73, 2665–2670. [Google Scholar] [CrossRef] [Green Version]
- Park, J.W.; Yu, J.H.; Kim, S.W.; Kweon, H.Y.; Choi, K.H.; Kim, S.R. Enhancement of antimicrobial peptide genes expression in Cactus mutated Bombyx mori cells by CRISPR/Cas9. Int. J. Ind. Entomol. 2018, 37, 21–28. [Google Scholar]
- Gan, B.H.; Gaynord, J.; Rowe, S.M.; Deingruber, T.; Spring, D.R. The multifaceted nature of antimicrobial peptides: Current synthetic chemistry approaches and future directions. Chem. Soc. Rev. 2021, 50, 7820–7880. [Google Scholar] [CrossRef]
- Yang, H.; Wang, M.; Zhang, P.; Sabhat, A.; Malik, F.A.; Bhaskar, R.; Zhou, F.; Li, X.H.; Hu, J.-B.; Sun, C.G.; et al. Cloning and characterization of the Bombyx mori ecdysone oxidase. Arch. Insect Biochem. Physiol. 2011, 78, 17–29. [Google Scholar] [CrossRef]
- Li, Z.; You, L.; Zeng, B.; Ling, L.; Xu, J.; Chen, X.; Zhang, Z.; Palli, S.R.; Huang, Y.; Tan, A. Ectopic expression of ecdysone oxidase impairs tissue degeneration in Bombyx mori. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150513. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Nagaoka, S. Identification and localization of a novel ecdysone oxidase in the silkworm, Bombyx mori. J. Insect Biotechnol. Sericol. 2017, 53, 49–53. [Google Scholar]
- Tettamanti, G.; Casartelli, M. Cell death during complete metamorphosis. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liu, X.; Shiotsuki, T.; Wang, Z.; Xu, X.; Huang, Y.; Li, M.; Li, K.; Tan, A. Depletion of juvenile hormone esterase extends larval growth in Bombyx mori. Insect Biochem. Mol. Biol. 2017, 81, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ma, S.; Chang, J.; Zhang, T.; Chen, X.; Liang, Y.; Xia, Q. Programmable targeted epigenetic editing using CRISPR system in Bombyx mori. Insect Biochem. Mol. Biol. 2019, 110, 105–111. [Google Scholar] [CrossRef]
- Xing, W.; Ma, S.; Liu, Y.; Xia, Q. CRISPR/dCas9-mediated imaging of endogenous genomic loci in living Bombyx mori cells. Insect Sci. 2019, 27, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Brady, D.; Saviane, A.; Cappellozza, S.; Sandrelli, F. An Efficient Workflow for Screening and Stabilizing CRISPR/Cas9-Mediated Mutant Lines in Bombyx mori. Methods Protoc. 2020, 4, 4. [Google Scholar] [CrossRef]
- Nartey, M.A.; Sun, X.; Qin, S.; Hou, C.X.; Li, M.W. CRISPR/Cas9-based knockout reveals that the clock gene timeless is indispensable for regulating circadian behavioral rhythms in Bombyx mori. Insect Sci. 2020, 28, 1414–1425. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Liu, X.; Ma, X.; Zhang, Z.; Wang, T.; Sun, F.; Hou, C.; Li, M. A palmitoyltransferase Approximated gene Bm-app regulates wing development in Bombyx mori. Insect Sci. 2020, 27, 2–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, B.; Huang, Y.; Xu, J.; Shiotsuki, T.; Bai, H.; Palli, S.R.; Huang, Y.; Tan, X.A. The FOXO transcription factor controls insect growth and development by regulating juvenile hormone degradation in the silkworm, Bombyx mori. J. Biol. Chem. 2017, 292, 11659–11669. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.W.; Jeong, C.Y.; Yu, J.H.; Kim, S.B.; Kang, S.K.; Kim, S.W.; Kim, N.S.; Kim, K.Y.; Park, J.W. Bombyx mori kynurenine 3-monooxygenase gene editing and insect molecular breeding using the clustered regularly interspaced short palindromic repeat/CRISPR associated protein 9 system. Biotechnol. Prog. 2020, 36, e3054. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, Z.; Yang, Y.; Huang, S.; Li, K.; He, L.; Zhou, X. Genome editing reveals the function of Yorkie during the embryonic and early larval development in silkworm, Bombyx mori. Insect Mol. Biol. 2018, 27, 675–685. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Liu, X.J.; Yu, Y.; Yang, F.Y.; Li, K. The receptor tyrosine kinase torso regulates ecdysone homeostasis to control developmental timing in Bombyx mori. Insect Sci. 2020, 28, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, Y.; Liu, Z.; Wang, Y.; He, L.; Li, K.; Huang, Y. Disruption of egg-specific protein causes female sterility in Bombyx mori. Insect Sci. 2021, 1–11. [Google Scholar] [CrossRef]
- Zhu, L.; Mon, H.; Xu, J.; Lee, J.M.; Kusakabe, T. CRISPR/Cas9-mediated knockout of factors in non-homologous end joining pathway enhances gene targeting in silkworm cells. Sci. Rep. 2015, 5, 18103. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, Y.C.; Jia, S.Z.; Liang, Y.T.; Tang, Y.; Xu, Y.S.; Kawasaki, H.; Wang, H.B. Imaginal disc growth factor maintains cuticle structure and controls melanization in the spot pattern formation of Bombyx mori. PLoS Genet. 2020, 16, e1008980. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, Z.; Zhang, R.; Yu, Y.; Yang, F.; Tan, A. Molecular Disruption of Ion Transport Peptide Receptor Results in Impaired Water Homeostasis and Developmental Defects in Bombyx mori. Front. Physiol. 2020, 11, 424. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; Xu, J.; Ling, L.; Zhang, R.; Shang, P.; Huang, Y.P. CRISPR disruption of TCTP gene impaired normal development in the silkworm Bombyx mori. Insect Sci. 2018, 26, 973–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhang, S.S.; Niu, B.L.; Ji, D.F.; Liu, X.J.; Li, M.W.; Bai, H.; Palli, S.R.; Wang, C.Z.; Tan, A.J. A determining factor for insect feeding preference in the silkworm, Bombyx mori. PLoS Biol. 2019, 17, e3000162. [Google Scholar] [CrossRef] [Green Version]
- Bi, H.; Xu, X.; Li, X.; Zhang, Y.; Huang, Y.; Li, K.; Xu, J. CRISPR Disruption of BmOvo Resulted in the Failure of Emergence and Affected the Wing and Gonad Development in the Silkworm Bombyx mori. Insects 2019, 10, 254. [Google Scholar] [CrossRef] [Green Version]
- Fujii, T.; Banno, Y. Enlargement of egg size by CRISPR/Cas9-mediated knockout of a sex-linked gene in the silkworm, Bombyx mori. J. Insect Biotechnol. Sericol. 2018, 87, 3_071–3_078. [Google Scholar] [CrossRef]
- Zhang, Z.; Aslam, A.F.; Liu, X.; Li, M.; Huang, Y.; Tan, A. Functional analysis of Bombyx Wnt1 during embryogenesis using the CRISPR/Cas9 system. J. Insect Physiol. 2015, 79, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Tian, L.; Guo, Z.; Guo, S.; Zhang, J.; Gu, S.; Palli, S.R.; Cao, Y.; Li, S. 20-Hydroxyecdysone (20E) Primary Response Gene E75 Isoforms Mediate Steroidogenesis Autoregulation and Regulate Developmental Timing in Bombyx. J. Biol. Chem. 2016, 291, 18163–18175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Liu, W.; Zeng, B.; Wang, G.; Hao, D.; Huang, Y. Deletion of the Bombyx mori odorant receptor co-receptor (BmOrco) impairs olfactory sensitivity in silkworms. Insect Biochem. Mol. Biol. 2017, 86, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Baddeley, H.J.E.; Isalan, M. The Application of CRISPR/Cas Systems for Antiviral Therapy. Front. Genome Ed. 2021, 3, 745559. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.G.; Wolf, T.; Vogel, H.; Castillo, D.; Woolard, K. An RNA-Directed Gene Editing Strategy for Attenuating the Infectious Potential of Feline Immunodeficiency Virus-Infected Cells: A Proof of Concept. Viruses 2020, 12, 511. [Google Scholar] [CrossRef] [PubMed]
- Fareh, M.; Zhao, W.; Hu, W.; Casan, J.M.; Kumar, A.; Symons, J.; Zerbato, J.M.; Fong, D.; Voskoboinik, I.; Ekert, P.G.; et al. Reprogrammed CRISPR-Cas13b suppresses SARS-CoV-2 replication and circumvents its mutational escape through mismatch tolerance. Nat. Commun. 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Yao, Q.; Li, M.W.; Wang, Y.; Wang, W.B.; Lu, J.; Dong, Y.; Chen, K.P. Screening of molecular markers for NPV resistance in Bombyx mori L. (Lep., Bombycidae). J. Appl. Entomol. 2003, 127, 134–136. [Google Scholar] [CrossRef]
- Guo, H.; Zhang, B.; Zheng, X.; Sun, J.; Guo, H.; Li, G.; Zhao, G.; Xu, A.; Qian, H. Pathogenicity Detection and Genome Analysis of Two Different Geographic Strains of BmNPV. Insects 2021, 12, 890. [Google Scholar] [CrossRef]
- Isobe, R.; Kojima, K.; Matsuyama, T.; Quan, G.X.; Kanda, T.; Tamura, T.; Sahara, K.; Asano, S.I.; Bando, H. Use of RNAi technology to confer enhanced resistance to BmNPV on transgenic silkworms. Arch. Virol. 2004, 149, 1931–1940. [Google Scholar] [CrossRef]
- Qiong, Y.; Dong Xu, X.; Qing Rong, L.; Yang, X.; Ming Qiang, Y. Standard method for detecting Bombyx mori nucleopolyhedrovirus disease-resistant silkworm varieties. Rev. Bras. Entomol. 2017, 62, 19–22. [Google Scholar] [CrossRef]
- Chen, S.; Hou, C.; Bi, H.; Wang, Y.; Xu, J.; Li, M.; James, A.A.; Huang, Y.; Tan, A. Transgenic Clustered Regularly Interspaced Short Palindromic Repeat/Cas9-Mediated Viral Gene Targeting for Antiviral Therapy of Bombyx mori Nucleopolyhedrovirus. J. Virol. 2017, 91, e02465-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Hu, Z.; Qin, Q.; Dong, F.; Huang, L.; Long, J.; Chen, P.; Lu, C.; Pan, M. CRISPR/Cas9 Mediated Disruption of the immediate early-0 and 2 as a Therapeutic Approach to Bombyx mori Nucleopolyhedrovirus in Transgenic Silkworm. Insect Mol. Biol. 2018, 28, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Qin, Q.; Hu, Z.; Chen, P.; Huang, L.; Zhang, X.; Tian, T.; Lu, C.; Pan, M. Construction of a One-Vector Multiplex CRISPR/Cas9 Editing System to Inhibit Nucleopolyhedrovirus Replication in Silkworms. Virol. Sin. 2019, 34, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Bi, H.; Wang, Y.; Li, X.; Xu, J.; Liu, Z.; He, L.; Li, K.; Huang, Y. Disruption of the ovarian serine protease (Osp) gene causes female sterility in Bombyx mori and Spodoptera litura. Pest Manag. Sci. 2019, 76, 1245–1255. [Google Scholar] [CrossRef]
- Sledzinski, P.; Nowaczyk, M.; Olejniczak, M. Computatonal Tools and Resources Supporting CRISPR-Cas Experiments. Cells 2020, 9, 1288. [Google Scholar] [CrossRef]


{kind=link}
| Traits | TALEN | ZFN | CRISPR-Cas | References |
|---|---|---|---|---|
| Origin | Prokaryotic | Eukaryotic | Prokaryotic | [41] |
| Efficiency (%) | 76 | 12 | 81 | [42] |
| Specificity | Moderate | Low | High | [43,44,45,46] |
| Target site recognition | Any site | Any site | Pam motif (NGG) required | [43] |
| Multiplex potential | Low | Low | High | [43,45] |
| Processing time | Time consuming | Time consuming | Short | [45] |
| Methylation sensitive | Sensitive | Sensitive | Not sensitive | [42] |
| Engineering feasibility | Moderate/High | Moderate | Moderate/High | [42,45] |
| Dimerization required | Yes | Yes | No | [44] |
| Cost effectiveness | Moderate | No | Yes | [43,44] |
| Type of Model Organism | Brief Description | Purpose | References |
|---|---|---|---|
| Human disease model | Transgenic B. mori expressing hIR (human insulin receptor) | Drug evaluation for diabetes treatment | [97,98] |
| Bacterial (Listeria monocytogenes) infection model in B. mori | Evaluating the interaction between host and pathogen; investigating the activity of vitamin A against microbial infections | [99] | |
| Fungal (C. albicans) infection model in silkworms | Assessing the C. albicans biofilm development | [100] | |
| Inducing deletions in the BmSpr gene, leading to sepiapterin reductase deficiency | Treatment options evaluations against sepiapterin reductase deficiency | [101] | |
| Model for pesticide toxicity | Exposing silkworms to phoxim | Identifying specific biomarkers for phoxim stress; evaluating the toxicity reaction and the pretreatment with nanoparticulate titanium dioxide | [102] |
| Inducing genotoxicity by feeding the silkworms with different doses of avermectin | Exploration of certain genes that are required for the DNA repairing mechanism | [103] | |
| Treating B. mori larvae with Fenvalerate-20EC | Evaluation of Fenvalerate-20EC impact on several digestive enzymes | [104] | |
| Model for drugs toxicity | Injecting the silkworms with three different pharmacologically active agents (4-methyl umbelliferone, 7-ethoxycoumarine) | Evaluation of the metabolic pathway of these compounds | [92] |
| Exposing the silkworms to fungal infections | Exploring pharmacokinetic parameters of an antifungal agent, Voriconazole | [105] | |
| Injecting cytotoxic drugs into B. mori larvae | Evaluation of cytotoxic drugs impact | [106] | |
| Model for nanomaterials toxicity | Spreading silver nanoparticles on mulberry leaves | Toxicity evaluation of silver nanoparticles | [107,108,109,110] |
| Injecting subcutaneously zinc oxide nanoparticles | Evaluation of zinc oxide nanoparticles toxicity, accumulation, and distribution | [111] | |
| Injecting in the dorsal vein different nanoparticles with great interest in various life science branches | Investigation of different silicon and carbon nanomaterials toxicity level against hemocytes | [112] |
| Target Gene | Mutation Type | Delivery Approach | Objective | Gene Function | References |
|---|---|---|---|---|---|
| BmTim | Deletions | Plasmid | Functional gene analysis | Exhibit an impact on the embryo hatching process | [138] |
| BmApp | Deletions, insertions | mRNA | Functional gene analysis | Regulates wing development and cell mitosis | [139] |
| BmFoxo | Deletions | Plasmid | Analysis of BmFoxo and JH interaction | Involved in JH degradation | [140] |
| BmKmo | Deletions | Protein | Phenotypic analysis | Involved in the process of egg formation and eye coloring | [141] |
| BmYki | Deletions | Plasmid | Functional gene analysis | Involved in organ development and regeneration | [142] |
| BmTorso | Deletions | Plasmid | Functional gene analysis | Maintain the steroid hormones balance | [143] |
| BmEsp | Deletions | Plasmid | Functional gene analysis | Involved in a female’s reproducibility | [144] |
| BmTudor | Deletions, insertions | Plasmid | Investigating the frequency of homologous recombination | Included in the stress granule formation | [145] |
| BmIdgf | Deletions | mRNA | Analyzing the pigmentation mechanism | Plays a key role in the melanization mechanism | [146] |
| BmBngr-a2 | Deletions | Plasmid | Exploring functional studies of certain ion transport peptides | Involved in water homeostasis | [147] |
| BmTctp | Deletions | Plasmid | Functional analysis | Involved in different cell process, such as growth, development, and proliferation | [148] |
| BmGr66 | Deletions | Plasmid | A better understanding of the specific feeding preference | Involved in silkworms’ specific feeding preferences | [149] |
| BmOvo | Deletions | Plasmid | Functional analysis | Involved in germline sex determination and wing metamorphosis | [150] |
| BmPhyhd1 | Deletions | Protein | Functional analysis | Exhibits a great impact on certain features of the epithelial cells | [151] |
| BmWnt1 | Deletions | mRNA | Functional analysis | Involved in the embryogenesis | [152] |
| BmE75b | Deletions | mRNA | Functional analysis | Controls the developmental timing | [153] |
| BmOrco | Deletions | Plasmid | Exploration of adult mating behavior | Involved in silkworms’ olfactory system, being an odorant receptor co-receptor | [154] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baci, G.-M.; Cucu, A.-A.; Giurgiu, A.-I.; Muscă, A.-S.; Bagameri, L.; Moise, A.R.; Bobiș, O.; Rațiu, A.C.; Dezmirean, D.S. Advances in Editing Silkworms (Bombyx mori) Genome by Using the CRISPR-Cas System. Insects 2022, 13, 28. https://doi.org/10.3390/insects13010028
Baci G-M, Cucu A-A, Giurgiu A-I, Muscă A-S, Bagameri L, Moise AR, Bobiș O, Rațiu AC, Dezmirean DS. Advances in Editing Silkworms (Bombyx mori) Genome by Using the CRISPR-Cas System. Insects. 2022; 13(1):28. https://doi.org/10.3390/insects13010028
Chicago/Turabian StyleBaci, Gabriela-Maria, Alexandra-Antonia Cucu, Alexandru-Ioan Giurgiu, Adriana-Sebastiana Muscă, Lilla Bagameri, Adela Ramona Moise, Otilia Bobiș, Attila Cristian Rațiu, and Daniel Severus Dezmirean. 2022. "Advances in Editing Silkworms (Bombyx mori) Genome by Using the CRISPR-Cas System" Insects 13, no. 1: 28. https://doi.org/10.3390/insects13010028
APA StyleBaci, G.-M., Cucu, A.-A., Giurgiu, A.-I., Muscă, A.-S., Bagameri, L., Moise, A. R., Bobiș, O., Rațiu, A. C., & Dezmirean, D. S. (2022). Advances in Editing Silkworms (Bombyx mori) Genome by Using the CRISPR-Cas System. Insects, 13(1), 28. https://doi.org/10.3390/insects13010028