
Perception of Daily Time: Insights from the Fruit Flies


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Drosophila Circadian Timers
3. The Time Cues
3.1. Light Inputs
3.2. Temperature Inputs

3.3. Adaptation to ‘Natural’ Input Conditions
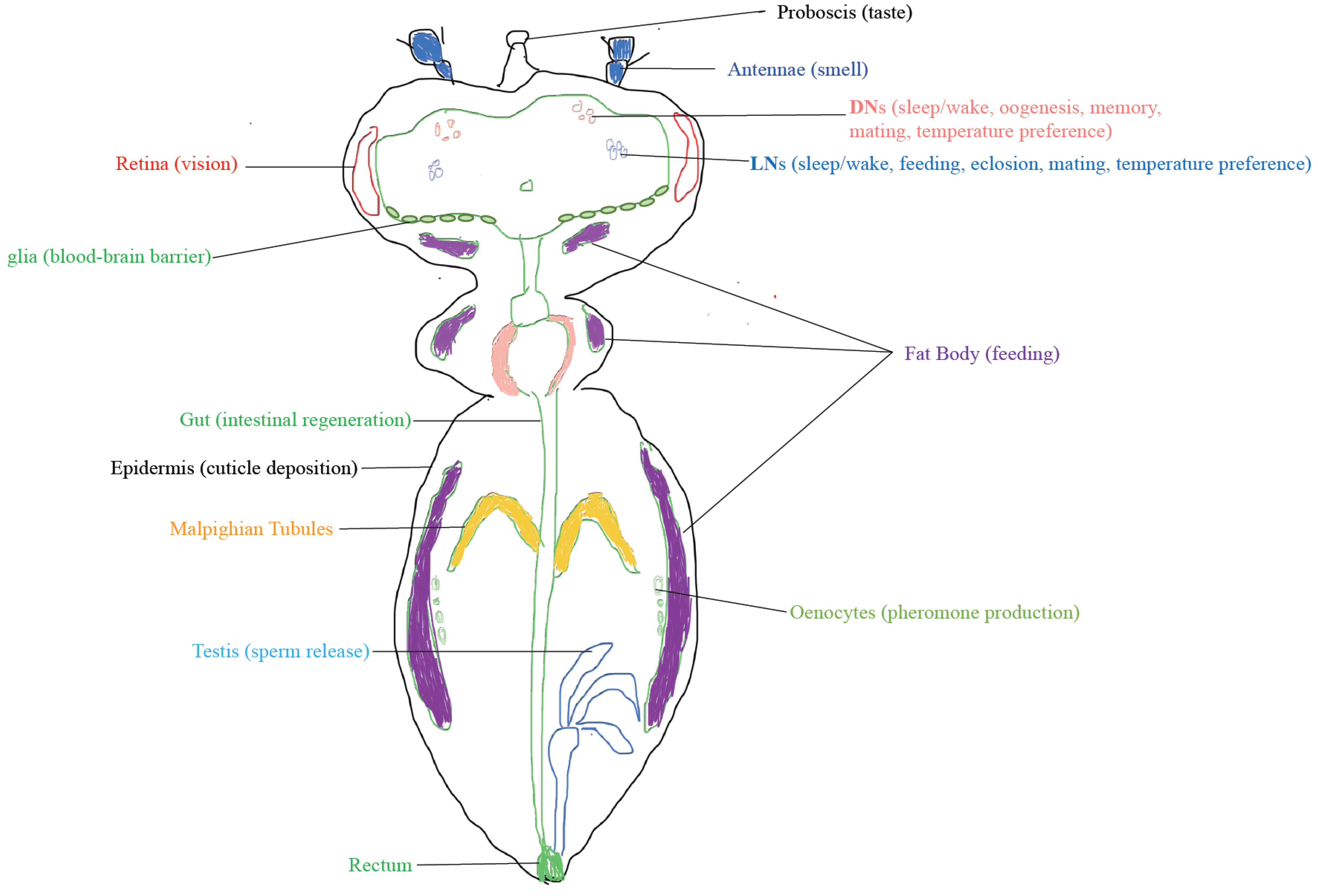
4. The Output of the Clock
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Grima, B.; Chélot, E.; Xia, R.; Rouyer, F. Morning and evening peaks of activity rely on different clock neurons of the Drosophila brain. Nature 2004, 431, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Stoleru, D.; Peng, Y.; Agosto, J.; Rosbash, M. Coupled oscillators control morning and evening locomotor behaviour of Drosophila. Nature 2004, 431, 862–868. [Google Scholar] [CrossRef]
- Stoleru, D.; Nawathean, P.; de la Paz Fernandez, M.; Menet, J.S.; Ceriani, M.F.; Rosbash, M. The Drosophila Circadian Network Is a Seasonal Timer. Cell 2007, 129, 207–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendricks, J.C.; Sehgal, A.; Pack, A.I. The need for a simple animal model to understand sleep. Prog. Neurobiol. 2000, 61, 339–351. [Google Scholar] [CrossRef]
- Paranjpe, D.A.; Anitha, D.; Kumar, S.; Kumar, D.; Verkhedkar, K.; Chandrashekaran, M.K.; Joshi, A.; Sharma, V.K. Entrainment of Eclosion Rhythm inDrosophila melanogasterPopulations Reared for More Than 700 Generations in Constant Light Environment. Chronobiol. Int. 2003, 20, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Myers, E.M. The Circadian Control of Eclosion. Chrono-Int. 2003, 20, 775–794. [Google Scholar] [CrossRef]
- De, J.; Varma, V.; Sharma, V.K. Adult Emergence Rhythm of Fruit FliesDrosophila melanogasterunder Seminatural Conditions. J. Biol. Rhythm. 2012, 27, 280–286. [Google Scholar] [CrossRef]
- Selcho, M.; Mühlbauer, B.; Hensgen, R.; Shiga, S.; Wegener, C.; Yasuyama, K. Anatomical characterization of PDF-tri neurons and peptidergic neurons associated with eclosion behavior in Drosophila. J. Comp. Neurol. 2018, 526, 1307–1328. [Google Scholar] [CrossRef]
- Chatterjee, A.; Tanoue, S.; Houl, J.H.; Hardin, P.E. Regulation of gustatory physiology and appetitive behavior by the Drosophila circadian clock. Curr. Biol. 2010, 20, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Zheng, X.; Sehgal, A. Regulation of Feeding and Metabolism by Neuronal and Peripheral Clocks in Drosophila. Cell Metab. 2008, 8, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Sakai, T.; Ishida, N. Time, love and species. Neuro Endocrinol. Lett. 2001, 22, 222–228. [Google Scholar]
- Hardeland, R. Species differences in the diurnal rhythmicity of courtship behaviour within the Melanogaster group of the genus Drosophila. Anim. Behav. 1972, 20, 170–174. [Google Scholar] [CrossRef]
- Nishinokubi, I.; Shimoda, M.; Ishida, N. Mating rhythms of Drosophila: Rescue of tim01 mutants by D. ananassae timeless. J. Circadian Rhythm. 2006, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Hamasaka, Y.; Suzuki, T.; Hanai, S.; Ishida, N. Evening circadian oscillator as the primary determinant of rhythmic motivation for Drosophila courtship behavior. Genes Cells 2010, 15, 1240–1248. [Google Scholar] [CrossRef]
- Krishnan, B.; Dryer, S.E.; Hardin, P.E. Circadian rhythms in olfactory responses of Drosophila melanogaster. Nature 1999, 400, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Tanoue, S.; Krishnan, P.; Krishnan, B.; Dryer, S.E.; Hardin, P.E. Circadian Clocks in Antennal Neurons Are Necessary and Sufficient for Olfaction Rhythms in Drosophila. Curr. Biol. 2004, 14, 638–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howlader, G.; Paranjpe, D.A.; Sharma, V.K. Non-Ventral Lateral Neuron-Based, Non-PDF-Mediated Clocks Control Circadian Egg-Laying Rhythm inDrosophila melanogaster. J. Biol. Rhythm. 2006, 21, 13–20. [Google Scholar] [CrossRef]
- Manjunatha, T.; Dass, S.H.; Sharma, V.K. Egg-laying rhythm in Drosophila melanogaster. J. Genet. 2008, 87, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Ito, C.; Goto, S.G.; Shiga, S.; Tomioka, K.; Numata, H. Peripheral circadian clock for the cuticle deposition rhythm in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2008, 105, 8446–8451. [Google Scholar] [CrossRef] [Green Version]
- Plautz, J.D.; Kaneko, M.; Hall, J.C.; Kay, S.A. Independent Photoreceptive Circadian Clocks Throughout Drosophila. Science 1997, 278, 1632–1635. [Google Scholar] [CrossRef]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef]
- Top, D.; Young, M.W. Coordination between Differentially Regulated Circadian Clocks Generates Rhythmic Behavior. Cold Spring Harb. Perspect. Biol. 2017, 10, a033589. [Google Scholar] [CrossRef] [Green Version]
- Helfrich-Förster, C.; Homberg, U. Pigment-dispersing hormone-immunoreactive neurons in the nervous system of wild-type Drosophila melanogaster and of several mutants with altered circadian rhythmicity. J. Comp. Neurol. 1993, 337, 177–190. [Google Scholar] [CrossRef]
- Renn, S.C.; Park, J.H.; Rosbash, M.; Hall, J.C.; Taghert, P.H. A pdf Neuropeptide Gene Mutation and Ablation of PDF Neurons Each Cause Severe Abnormalities of Behavioral Circadian Rhythms in Drosophila. Cell 1999, 99, 791–802. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Helfrich-Förster, C.; Lee, G.; Liu, L.; Rosbash, M.; Hall, J.C. Differential regulation of circadian pacemaker output by separate clock genes in Drosophila. Proc. Natl. Acad. Sci. USA 2000, 97, 3608–3613. [Google Scholar] [CrossRef]
- Im, S.H.; Taghert, P.H. PDF receptor expression reveals direct interactions between circadian oscillators in Drosophila. J. Comp. Neurol. 2010, 518, 1925–1945. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Lamaze, A.; De, J.; Mena, W.; Chélot, E.; Martin, B.; Hardin, P.; Kadener, S.; Emery, P.; Rouyer, F. Reconfiguration of a Multi-oscillator Network by Light in the Drosophila Circadian Clock. Curr. Biol. 2018, 28, 2007–2017.e4. [Google Scholar] [CrossRef] [Green Version]
- Pittendrigh, C.; Daan, S.A. Functional analysis of circadian pacemakers in nocturnal rodents. V. Pacemaker structure: A clock for all seasons. J. Comp. Physiol. 1976, 106, 333–335. [Google Scholar] [CrossRef]
- Jagota, A.; De La Iglesia, H.O.; Schwartz, W.J. Morning and evening circadian oscillations in the suprachiasmatic nucleus in vitro. Nat. Neurosci. 2000, 3, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Rieger, D.; Shafer, O.T.; Tomioka, K.; Helfrich-Förster, C. Functional Analysis of Circadian Pacemaker Neurons in Drosophila melanogaster. J. Neurosci. 2006, 26, 2531–2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.H. Forty Years of Prcs-What Have We Learned? Chrono-Int. 1999, 16, 711–743. [Google Scholar] [CrossRef]
- Rusak, B.; Boulos, Z. pathways for photic entrainment of mammalian circadian rhythms. Photochem. Photobiol. 1981, 34, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Reebs, S.G.; Mrosovsky, N. Effects of Induced Wheel Running on the Circadian Activity Rhythms of Syrian Hamsters: Entrainment and Phase Response Curve. J. Biol. Rhythm. 1989, 4, 39–48. [Google Scholar] [CrossRef]
- Mistlberger, R.E.; Skene, D.J. Social influences on mammalian circadian rhythms: Animal and human studies. Biol. Rev. 1999, 79, 533–556. [Google Scholar] [CrossRef]
- Golombek, D.A.; Rosenstein, R.E. Physiology of Circadian Entrainment. Physiol. Rev. 2010, 90, 1063–1102. [Google Scholar] [CrossRef] [Green Version]
- Kenagy, G.J.; Sharbaugh, S.M.; Nagy, K.A. Annual cycle of energy and time expenditure in a golden-mantled ground squirrel population. Oecologia 1989, 78, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Hut, R.A.; Van Oort, B.E.H.; Daan, S. Natural entrainment without dawn and dusk: The case of the European ground squirrel (Spermophilus citellus). J. Biol. Rhythm. 1999, 14, 290–299. [Google Scholar] [CrossRef]
- Stanewsky, R.; Kaneko, M.; Emery, P.; Beretta, B.; Wager-Smith, K.; Kay, S.A.; Rosbash, M.; Hall, J.C. The cryb Mutation Identifies Cryptochrome as a Circadian Photoreceptor in Drosophila. Cell 1998, 95, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Emery, P.; Stanewsky, R.; Helfrich-Förster, C.; Emery-Le, M.; Hall, J.C.; Rosbash, M. Drosophila CRY Is a Deep Brain Circadian Photoreceptor. Neuron 2000, 26, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Ceriani, M.F.; Darlington, T.K.; Staknis, D.; Más, P.; Petti, A.A.; Weitz, C.J.; Kay, S.A. Light-Dependent Sequestration of TIMELESS by CRYPTOCHROME. Science 1999, 285, 553–556. [Google Scholar] [CrossRef]
- Lin, F.J.; Song, W.; Meyer-Bernstein, E.; Naidoo, N.; Sehgal, A. Photic Signaling by Cryptochrome in the Drosophila Circadian System. Mol. Cell. Biol. 2001, 21, 7287–7294. [Google Scholar] [CrossRef] [Green Version]
- Koh, K.; Zheng, X.; Sehgal, A. JETLAG Resets the Drosophila Circadian Clock by Promoting Light-Induced Degradation of TIMELESS. Science 2006, 312, 1809–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, M.P.; Wager-Smith, K.; Rothenfluh-Hilfiker, A.; Young, M.W. Light-Induced Degradation of TIMELESS and Entrainment of the Drosophila Circadian Clock. Science 1996, 271, 1736–1740. [Google Scholar] [CrossRef]
- Suri, V.; Qian, Z.; Hall, J.C.; Rosbash, M. Evidence that the TIM Light Response Is Relevant to Light-Induced Phase Shifts in Drosophila melanogaster. Neuron 1998, 21, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Roberts, L.; Leise, T.L.; Noguchi, T.; Galschiodt, A.M.; Houl, J.H.; Welsh, D.K.; Holmes, T.C. Light Evokes Rapid Circadian Network Oscillator Desynchrony Followed by Gradual Phase Retuning of Synchrony. Curr. Biol. 2015, 25, 858–867. [Google Scholar] [CrossRef] [Green Version]
- Kistenpfennig, C.; Nakayama, M.; Nihara, R.; Tomioka, K.; Helfrich-Förster, C.; Yoshii, T. A Tug-of-War between Cryptochrome and the Visual System Allows the Adaptation of Evening Activity to Long Photoperiods inDrosophila melanogaster. J. Biol. Rhythm. 2017, 33, 24–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.D.; Baik, L.S.; Holmes, T.C.; Montell, C. A rhodopsin in the brain functions in circadian photoentrainment in Drosophila. Nature 2017, 545, 340–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senthilan, P.R.; Grebler, R.; Reinhard, N.; Rieger, D.; Helfrich-Förster, C. Role of Rhodopsins as Circadian Photoreceptors in the Drosophila melanogaster. Biology 2019, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helfrich-Förster, C.; Winter, C.; Hofbauer, A.; Hall, J.C.; Stanewsky, R. The Circadian Clock of Fruit Flies Is Blind after Elimination of All Known Photoreceptors. Neuron 2001, 30, 249–261. [Google Scholar] [CrossRef] [Green Version]
- Alejevski, F.; Saint-Charles, A.; Michard-Vanhée, C.; Martin, B.; Galant, S.; Vasiliauskas, D.; Rouyer, F. The HisCl1 histamine receptor acts in photoreceptors to synchronize Drosophila behavioral rhythms with light-dark cycles. Nat. Commun. 2019, 10, 252. [Google Scholar] [CrossRef]
- Yoshii, T.; Heshiki, Y.; Ibuki-Ishibashi, T.; Matsumoto, A.; Tanimura, T.; Tomioka, K. Temperature cycles driveDrosophilacircadian oscillation in constant light that otherwise induces behavioural arrhythmicity. Eur. J. Neurosci. 2005, 22, 1176–1184. [Google Scholar] [CrossRef]
- Shakhmantsir, I.; Nayak, S.; Grant, G.R.; Sehgal, A. Spliceosome factors target timeless (tim) mRNA to control clock protein accumulation and circadian behavior in Drosophila. Elife 2018, 7, e39821. [Google Scholar] [CrossRef] [PubMed]
- Miyasako, Y.; Umezaki, Y.; Tomioka, K. Separate Sets of Cerebral Clock Neurons Are Responsible for Light and Temperature Entrainment of Drosophila Circadian Locomotor Rhythms. J. Biol. Rhythm. 2007, 22, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Yadlapalli, S.; Jiang, C.; Bahle, A.; Reddy, P.; Meyhofer, E.; Shafer, O.T. Circadian clock neurons constantly monitor environmental temperature to set sleep timing. Nat. Cell Biol. 2018, 555, 98–102. [Google Scholar] [CrossRef]
- George, R.; Stanewsky, R. Peripheral Sensory Organs Contribute to Temperature Synchronization of the Circadian Clock in Drosophila melanogaster. Front. Physiol. 2021, 12, 622545. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, T.; Vanin, S.; Costa, R.; Helfrich-Förster, C. Synergic Entrainment of Drosophila’s Circadian Clock by Light and Temperature. J. Biol. Rhythm. 2009, 24, 452–464. [Google Scholar] [CrossRef]
- Harper, R.E.F.; Dayan, P.; Albert, J.T.; Stanewsky, R. Sensory Conflict Disrupts Activity of the Drosophila Circadian Network. Cell Rep. 2016, 17, 1711–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edery, I.; Rutila, J.E.; Rosbash, M. Phase Shifting of the Circadian Clock by Induction of the Drosophila period Protein. Science 1994, 263, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Gentile, C.; Sehadova, H.; Simoni, A.; Chen, C.; Stanewsky, R. Cryptochrome Antagonizes Synchronization of Drosophila’s Circadian Clock to Temperature Cycles. Curr. Biol. 2013, 23, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Menegazzi, P.; Yoshii, T.; Helfrich-Förster, C. Laboratory versus Nature. J. Biol. Rhythm. 2012, 27, 433–442. [Google Scholar] [CrossRef]
- Vanin, S.; Bhutani, S.; Montelli, S.; Menegazzi, P.; Green, E.W.; Pegoraro, M.; Sandrelli, F.; Costa, R.; Kyriacou, C.P. Unexpected features of Drosophila circadian behavioural rhythms under natural conditions. Nature 2012, 484, 371–375. [Google Scholar] [CrossRef]
- De, J.; Varma, V.; Saha, S.; Sheeba, V.; Sharma, V.K. Significance of activity peaks in fruit flies, Drosophila melanogaster, under seminatural conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 8984–8989. [Google Scholar] [CrossRef] [Green Version]
- Prabhakaran, P.M.; Sheeba, V. Insights into differential activity patterns of Drosophilids under semi-natural conditions. J. Exp. Biol. 2013, 216, 4691–4702. [Google Scholar] [CrossRef] [Green Version]
- Prabhakaran, P.M.; De, J.; Sheeba, V. Natural Conditions Override Differences in Emergence Rhythm among Closely Related Drosophilids. PLoS ONE 2013, 8, e83048. [Google Scholar] [CrossRef]
- Green, E.W.; O’Callaghan, E.K.; Hansen, C.N.; Bastianello, S.; Bhutani, S.; Vanin, S.; Armstrong, J.D.; Costa, R.; Kyriacou, C.P. Drosophila circadian rhythms in seminatural environments: Summer afternoon component is not an artifact and requires TrpA1 channels. Proc. Natl. Acad. Sci. USA 2015, 112, 8702–8707. [Google Scholar] [CrossRef] [Green Version]
- Kauranen, H.; Menegazzi, P.; Costa, R.; Helfrich-Förster, C.; Kankainen, A.; Hoikkala, A. Flies in the North. J. Biol. Rhythm. 2012, 27, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Menegazzi, P.; Benetta, E.D.; Beauchamp, M.; Schlichting, M.; Steffan-Dewenter, I.; Helfrich-Förster, C. Adaptation of Circadian Neuronal Network to Photoperiod in High-Latitude European Drosophilids. Curr. Biol. 2017, 27, 833–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchamp, M.; Bertolini, E.; Deppisch, P.; Steubing, J.; Menegazzi, P.; Helfrich-Förster, C. Closely Related Fruit Fly Species Living at Different Latitudes Diverge in Their Circadian Clock Anatomy and Rhythmic Behavior. J. Biol. Rhythm. 2018, 33, 602–613. [Google Scholar] [CrossRef]
- Pittendrigh, C.S.; Takamura, T. Latitudinal Clines in the Properties of a Circadian Pacemaker. J. Biol. Rhythm. 1989, 4, 105–123. [Google Scholar] [CrossRef] [Green Version]
- Pittendrigh, C.S.; Kyner, W.T.; Takamura, T. The Amplitude of Circadian Oscillations: Temperature Dependence, Latitudinal Clines, and the Photoperiodic Time Measurement. J. Biol. Rhythm. 1991, 6, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, E.; Schubert, F.K.; Zanini, D.; Sehadová, H.; Helfrich-Förster, C.; Menegazzi, P. Life at High Latitudes Does Not Require Circadian Behavioral Rhythmicity under Constant Darkness. Curr. Biol. 2019, 29, 3928–3936.e3. [Google Scholar] [CrossRef] [Green Version]
- Picot, M.; Cusumano, P.; Klarsfeld, A.; Ueda, R.; Rouyer, F. Light Activates Output from Evening Neurons and Inhibits Output from Morning Neurons in the Drosophila Circadian Clock. PLoS Biol. 2007, 5, e315. [Google Scholar] [CrossRef]
- Majercak, J.; Sidote, D.; Hardin, P.E.; Edery, I. How a Circadian Clock Adapts to Seasonal Decreases in Temperature and Day Length. Neuron 1999, 24, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Collins, B.H.; Rosato, E.; Kyriacou, C.P. Seasonal behavior inDrosophila melanogasterrequires the photoreceptors, the circadian clock, and phospholipase C. Proc. Natl. Acad. Sci. USA 2004, 101, 1945–1950. [Google Scholar] [CrossRef] [Green Version]
- Majercak, J.; Chen, W.-F.; Edery, I. Splicing of the period Gene 3′-Terminal Intron Is Regulated by Light, Circadian Clock Factors, and Phospholipase C Splicing of the period Gene 3 J-Terminal Intron Is Regulated by Light, Circadian Clock Factors, and Phospholipase C. Mol. Cell. Biol. 2004, 24, 3359–3372. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Edery, I. Daywake, an Anti-siesta Gene Linked to a Splicing-Based Thermostat from an Adjoining Clock Gene. Curr. Biol. 2019, 29, 1728–1734.e4. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-F.; Majercak, J.; Edery, I. Clock-Gated Photic Stimulation of Timeless Expression at Cold Temperatures and Seasonal Adaptation in Drosophila. J. Biol. Rhythm. 2006, 21, 256–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anduaga, A.M.; Evantal, N.; Patop, I.L.; Bartok, O.; Weiss, R.; Kadener, S. Author response: Thermosensitive alternative splicing senses and mediates temperature adaptation in Drosophila. Elife 2019, 8, e44642. [Google Scholar] [CrossRef] [PubMed]
- Foley, L.E.; Ling, J.; Joshi, R.; Evantal, N.; Kadener, S.; Emery, P. Drosophila PSI controls circadian period and the phase of circadian behavior under temperature cycle via tim splicing. Elife 2019, 8, e50063. [Google Scholar] [CrossRef]
- Cao, G.; Nitabach, M.N. Circadian Control of Membrane Excitability in Drosophila melanogaster Lateral Ventral Clock Neurons. J. Neurosci. 2008, 28, 6493–6501. [Google Scholar] [CrossRef]
- Sheeba, V.; Gu, H.; Sharma, V.K.; O’Dowd, D.K.; Holmes, T.C. Circadian- and Light-Dependent Regulation of Resting Membrane Potential and Spontaneous Action Potential Firing of Drosophila Circadian Pacemaker Neurons. J. Neurophysiol. 2008, 99, 976–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flourakis, M.; Kula-Eversole, E.; Hutchison, A.L.; Han, T.H.; Aranda, K.; Moose, D.L.; White, K.P.; Dinner, A.R.; Lear, B.C.; Ren, D.; et al. A Conserved Bicycle Model for Circadian Clock Control of Membrane Excitability. Cell 2015, 162, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lear, B.C.; Lin, J.-M.; Keath, J.R.; McGill, J.J.; Raman, I.M.; Allada, R. The Ion Channel Narrow Abdomen Is Critical for Neural Output of the Drosophila Circadian Pacemaker. Neuron 2005, 48, 965–976. [Google Scholar] [CrossRef] [Green Version]
- Klose, M.K.; Bruchez, M.P.; Deitcher, D.L.; Levitan, E.S. Temporally and spatially partitioned neuropeptide release from individual clock neurons. Proc. Natl. Acad. Sci. USA 2021, 118, e2101818118. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.P.; Berni, J.; Ceriani, M.F. Circadian Remodeling of Neuronal Circuits Involved in Rhythmic Behavior. PLoS Biol. 2008, 6, e69. [Google Scholar] [CrossRef] [Green Version]
- Sivachenko, A.; Li, Y.; Abruzzi, K.C.; Rosbash, M. The Transcription Factor Mef2 Links the Drosophila Core Clock to Fas2, Neuronal Morphology, and Circadian Behavior. Neuron 2013, 79, 281–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorostiza, E.A.; Depetris-Chauvin, A.; Frenkel, L.; Pírez, N.; Ceriani, M.F. Circadian Pacemaker Neurons Change Synaptic Contacts across the Day. Curr. Biol. 2014, 24, 2161–2167. [Google Scholar] [CrossRef] [Green Version]
- Petsakou, A.; Sapsis, T.P.; Blau, J. Circadian Rhythms in Rho1 Activity Regulate Neuronal Plasticity and Network Hierarchy. Cell 2015, 162, 823–835. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, M.P.; Pettibone, H.L.; Bogart, J.T.; Roell, C.J.; Davey, C.E.; Pranevicius, A.; Huynh, K.V.; Lennox, S.M.; Kostadinov, B.S.; Shafer, O.T. Sites of Circadian Clock Neuron Plasticity Mediate Sensory Integration and Entrainment. Curr. Biol. 2020, 30, 2225–2237.e5. [Google Scholar] [CrossRef]
- Duhart, J.M.; Herrero, A.; de la Cruz, G.; Ispizua, J.I.; Pírez, N.; Ceriani, M.F. Circadian Structural Plasticity Drives Remodeling of E Cell Output. Curr. Biol. 2020, 30, 5040–5048. [Google Scholar] [CrossRef]
- Liang, X.; Holy, T.E.; Taghert, P.H. Synchronous Drosophila circadian pacemakers display nonsynchronous Ca2+ rhythms in vivo. Science 2016, 351, 976–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Ho, M.C.W.; Zhang, Y.; Li, Y.; Wu, M.N.; Holy, T.E.; Taghert, P.H. Morning and Evening Circadian Pacemakers Independently Drive Premotor Centers via a Specific Dopamine Relay. Neuron 2019, 102, 843–857.e4. [Google Scholar] [CrossRef]
- Lamaze, A.; Stanewsky, R. DN1p or the “Fluffy” Cerberus of Clock Outputs. Front. Physiol. 2020, 10, 1540. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Holla, M.; Diaz, M.; Rosbash, M. A Circadian Output Circuit Controls Sleep-Wake Arousal in Drosophila. Neuron 2018, 100, 624–635.e4. [Google Scholar] [CrossRef] [Green Version]
- Lamaze, A.; Krätschmer, P.; Chen, K.-F.; Lowe, S.; Jepson, J.E. A Wake-Promoting Circadian Output Circuit in Drosophila. Curr. Biol. 2018, 28, 3098–3105.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavey, M.; Collins, B.; Bertet, C.; Blau, M.J. Circadian rhythms in neuronal activity propagate through output circuits. Nat. Neurosci. 2016, 19, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Ceriani, M.F.; Hogenesch, J.B.; Yanovsky, M.; Panda, S.; Straume, M.; Kay, S.A. Genome-wide expression analysis in Drosophila reveals genes controlling circadian behavior. J. Neurosci. 2002, 22, 9305–9319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaramillo, A.M.; Zheng, X.; Zhou, Y.; Amado, D.A.; Sheldon, A.; Sehgal, A.; Levitan, I.B. Pattern of distribution and cycling of SLOB, Slowpoke channel binding protein, in Drosophila. BMC Neurosci. 2004, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, M.P.; Chu, J.; Villella, A.; Atkinson, N.; Kay, S.A.; Ceriani, M.F. Impaired clock output by altered connectivity in the circadian network. Proc. Natl. Acad. Sci. USA 2007, 104, 5650–5655. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, D.; Bajwa, S.T.; Vanani, N.; Bajwa, T.A.; Cavanaugh, D.J. Slowpoke functions in circadian output cells to regulate rest:activity rhythms. PLoS ONE 2021, 16, e0249215. [Google Scholar] [CrossRef]
- Cavanaugh, D.J.; Geratowski, J.D.; Wooltorton, J.R.A.; Spaethling, J.M.; Hector, C.E.; Zheng, X.; Johnson, E.C.; Eberwine, J.H.; Sehgal, A. Identification of a Circadian Output Circuit for Rest:Activity Rhythms in Drosophila. Cell 2014, 157, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Barber, A.F.; Fong, S.Y.; Kolesnik, A.; Fetchko, M.; Sehgal, A. Drosophila clock cells use multiple mechanisms to transmit time-of-day signals in the brain. Proc. Natl. Acad. Sci. USA 2021, 118, e2019826118. [Google Scholar] [CrossRef] [PubMed]
- King, A.N.; Barber, A.F.; Smith, A.E.; Dreyer, A.P.; Sitaraman, D.; Nitabach, M.N.; Cavanaugh, D.J.; Sehgal, A. A Peptidergic Circuit Links the Circadian Clock to Locomotor Activity. Curr. Biol. 2017, 27, 1915–1927.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyer, A.P.; Martin, M.M.; Fulgham, C.V.; Jabr, D.A.; Bai, L.; Beshel, J.; Cavanaugh, D.J. A circadian output center controlling feeding:fasting rhythms in Drosophila. PLoS Genet. 2019, 15, e1008478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De, J.; Chatterjee, A. Perception of Daily Time: Insights from the Fruit Flies. Insects 2022, 13, 3. https://doi.org/10.3390/insects13010003
De J, Chatterjee A. Perception of Daily Time: Insights from the Fruit Flies. Insects. 2022; 13(1):3. https://doi.org/10.3390/insects13010003
Chicago/Turabian StyleDe, Joydeep, and Abhishek Chatterjee. 2022. "Perception of Daily Time: Insights from the Fruit Flies" Insects 13, no. 1: 3. https://doi.org/10.3390/insects13010003
APA StyleDe, J., & Chatterjee, A. (2022). Perception of Daily Time: Insights from the Fruit Flies. Insects, 13(1), 3. https://doi.org/10.3390/insects13010003