
Discrepancy in Sterol Usage between Two Polyphagous Caterpillars, Mythimna separata and Spodoptera frugiperda

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Culture and Artificial Diet
2.2. Growth Requirement of Cholesterol
2.3. Larval Response to Atypical Sterol Diets
2.4. Larval Performance on Cholestanone
2.5. Statistical Analyses
3. Results
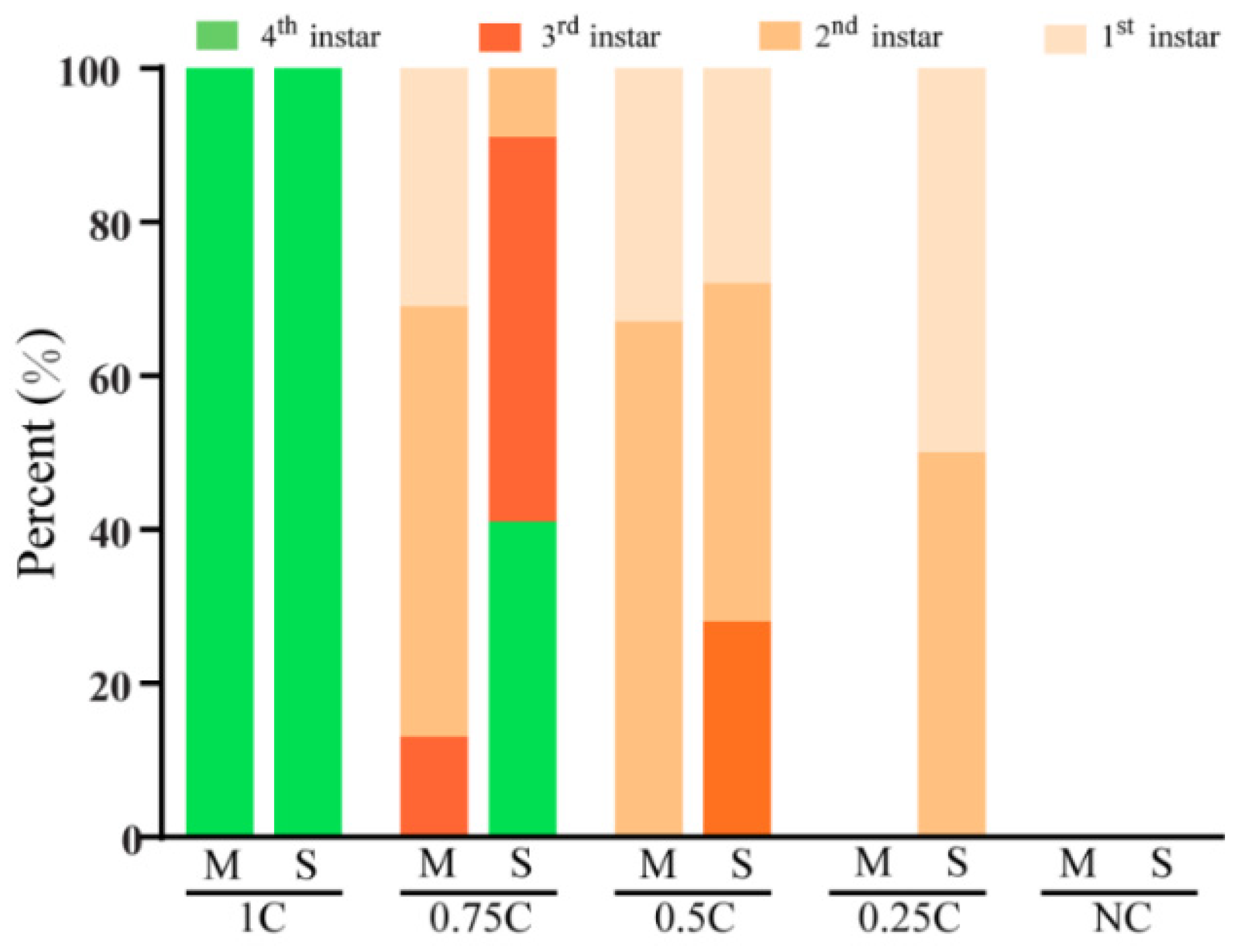
3.1. Growth Requirement of Cholesterol
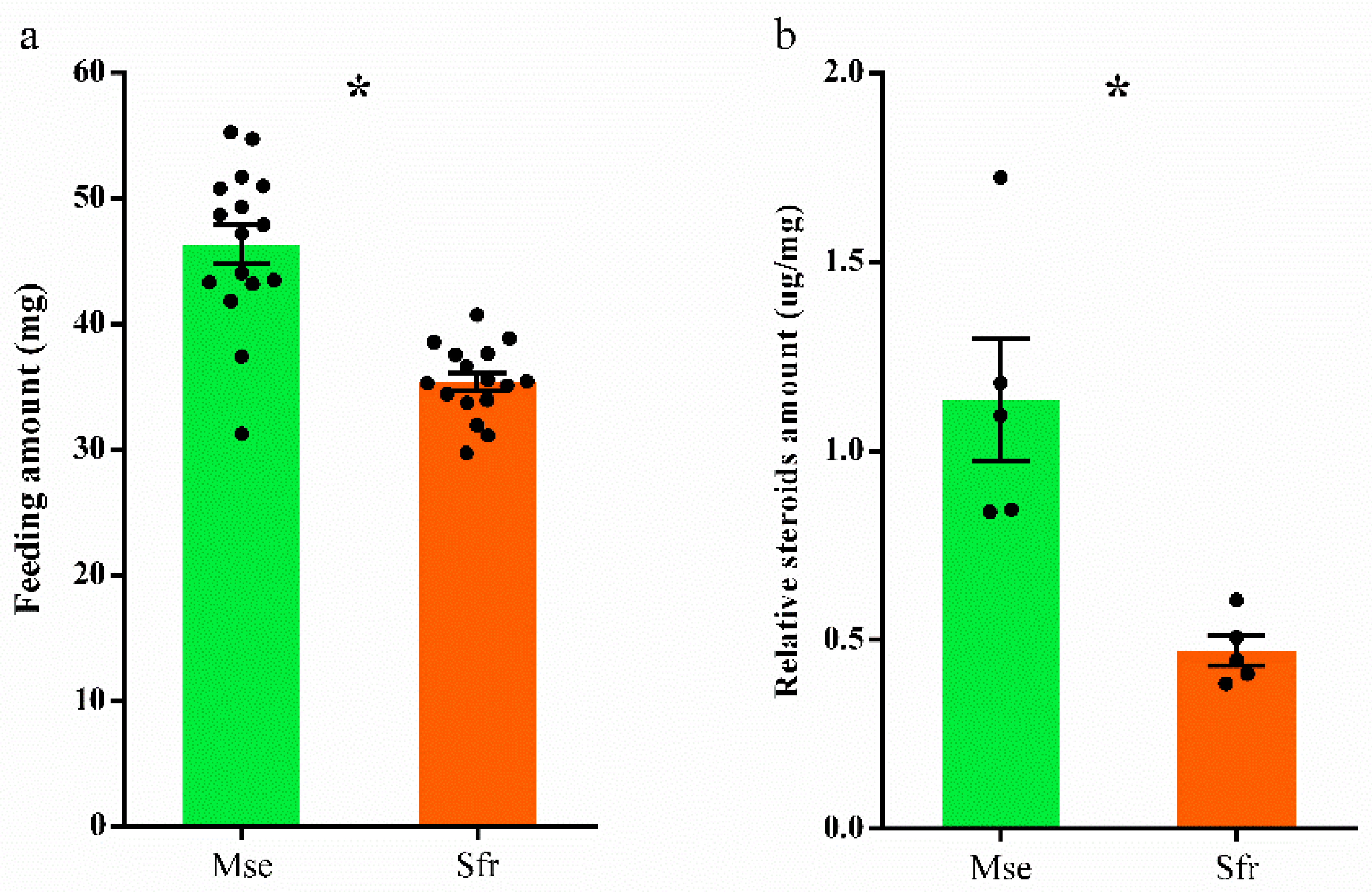
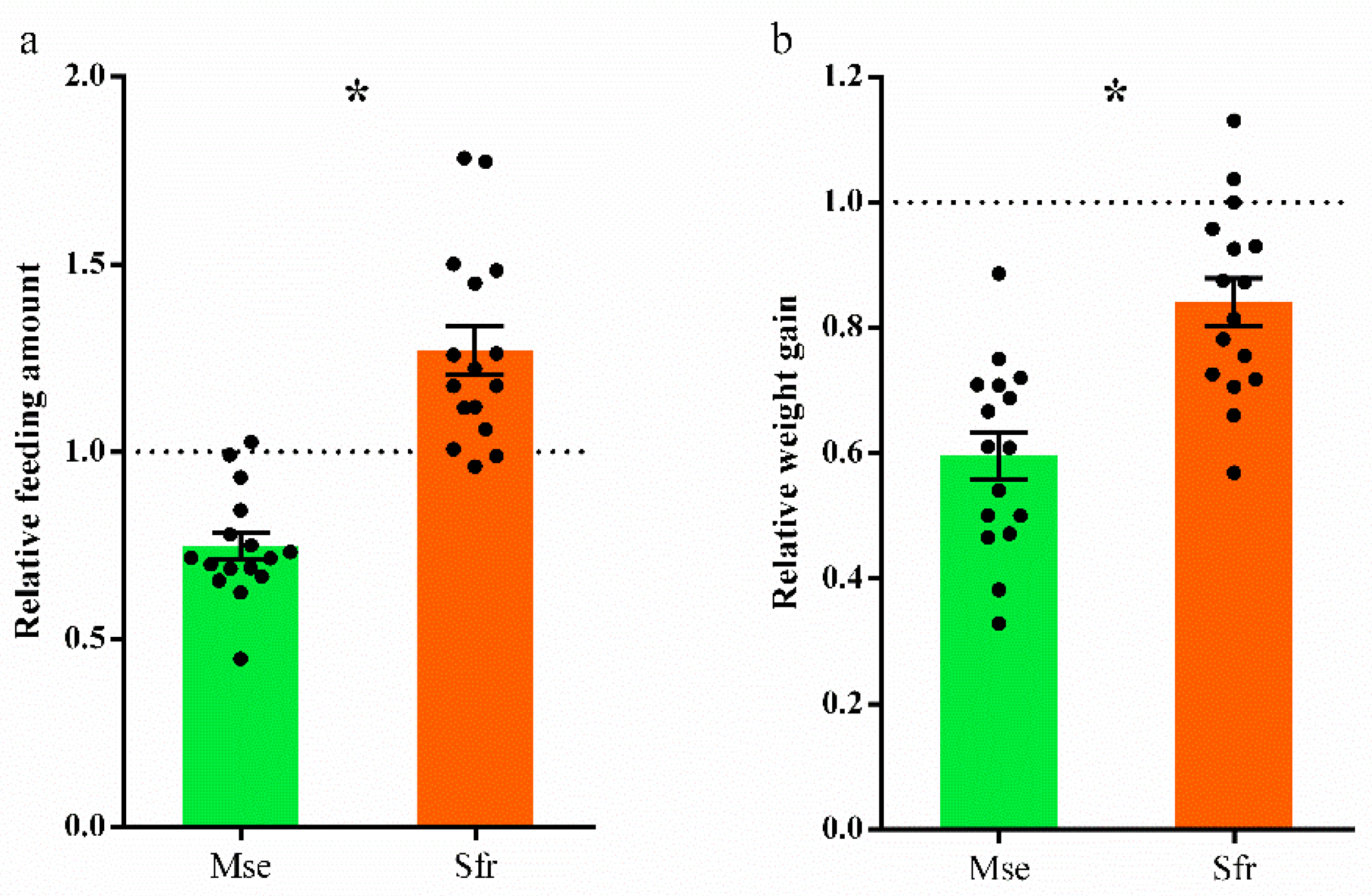
3.2. Effects of Atypical Steroids on Larval Performance
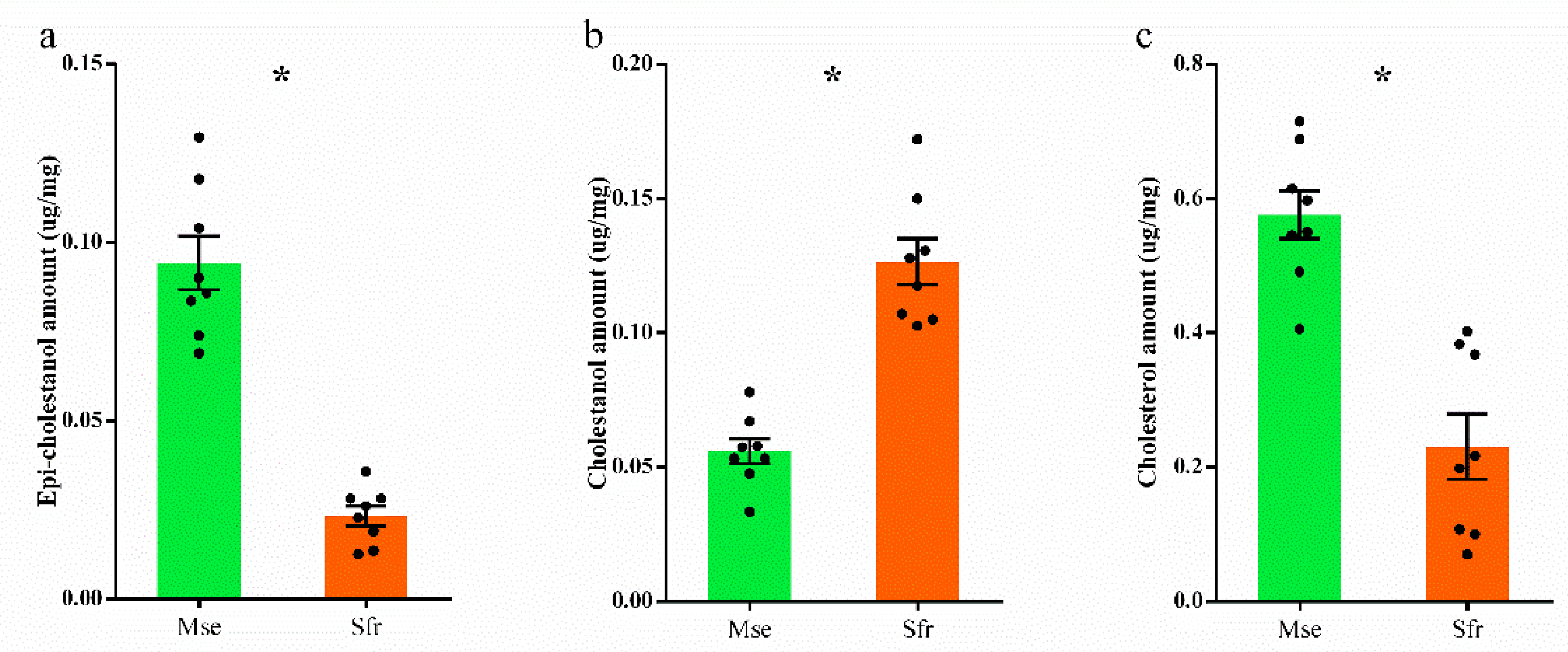
3.3. Larval Response to Cholestanone
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nboyine, J.A.; Kusi, F.; Abudulai, M.; Badii, B.K.; Zakaria, M.; Adu, G.B.; Haruna, A.; Seidu, A.; Osei, V.; Alhassan, S.; et al. A new pest, Spodoptera frugiperda (J.E. Smith), in tropical Africa: Its seasonal dynamics and damage in maize fields in northern Ghana. Crop Prot. 2020, 127, 104960. [Google Scholar] [CrossRef]
- Jiang, X.F.; Zhang, L.; Cheng, Y.X.; Luo, L.Z. Novel features, occurrence trends and economic impact of the oriental armyworm, Mythimna separata (Walker) in China. Chin. J. Appl. Entomol. 2014, 51, 1444–1449. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Asad, S.; Rehan, S.; David, A.; Liu, S.S.; Furlong, M.J. Estimating the economic cost of one of the world’s major insect pests, Plutella xylostella (Lepidoptera: Plutellidae): Just how long is a piece of string? J. Econ. Entomol. 2012, 105, 1115–1129. [Google Scholar] [CrossRef] [PubMed]
- Sheng, C.F.; Wang, H.T.; Sheng, S.Y.; Gao, L.D.; Xuan, W.J. Pest status and loss assessment of crop damage caused by the rice borers, Chilo suppressalis and Tryporyza incertulas in China. Entomol. Knowl. 2003, 40, 289–294. [Google Scholar]
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef]
- Sharma, H.C.; Sullivan, D.J.; Bhatnagar, V.S. Population dynamics and natural mortality factors of the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae), in South-Central India. Crop Prot. 2002, 21, 721–732. [Google Scholar] [CrossRef]
- Carvalho, R.A.; Omoto, C.; Field, L.M.; Williamson, M.S.; Bass, C. Investigating the molecular mechanisms of organophosphate and pyrethroid resistance in the fall armyworm Spodoptera frugiperda. PLoS ONE 2013, 8, e62268. [Google Scholar] [CrossRef]
- Song, Y.Q.; Wang, H.T.; Chen, Y.G.; Wang, S.Y.; Sun, H.Z. Cross-resistance and biochemical resistance mechanisms of emamectin benzoate resistant population of Mythimna separata. Chin. J. Pestic. Sci. 2017, 19, 18–24. [Google Scholar] [CrossRef]
- Yang, J.; Quan, Y.; Sivaprasath, P.; Shabbir, M.Z.; Wang, Z.; Ferre, J.; He, K. Insecticidal activity and synergistic combinations of ten different Bt toxins against Mythimna separata (Walker). Toxins 2018, 10, 454. [Google Scholar] [CrossRef]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Espenshade, P.J.; Hughes, A.L. Regulation of sterol synthesis in eukaryotes. Annu. Rev. Genet. 2007, 41, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Haines, T.H. Do sterols reduce proton and sodium leaks through lipid bilayers? Prog. Lipid Res. 2001, 40, 299–324. [Google Scholar] [CrossRef]
- Carvalho, M.; Schwudke, D.; Sampaio, J.L.; Palm, W.; Riezman, I.; Dey, G.; Gupta, G.D.; Mayor, S.; Riezman, H.; Shevchenko, A.; et al. Survival strategies of a sterol auxotroph. Development 2010, 137, 3675–3685. [Google Scholar] [CrossRef] [PubMed]
- Lingwood, D.; Simons, K. Lipid rafts as a membrane-organizing principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Head, B.P.; Patel, H.H.; Insel, P.A. Interaction of membrane/lipid rafts with the cytoskeleton: Impact on signaling and function: Membrane/lipid rafts, mediators of cytoskeletal arrangement and cell signaling. Biochim. Biophys. Acta 2014, 1838, 532–545. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.I.; Warren, J.T. A molecular genetic approach to the biosynthesis of the insect steroid molting hormone. In Insect Hormones; Vitamins & Hormones; Elsevier: Amsterdam, The Netherlands, 2005; pp. 31–57. [Google Scholar]
- Liu, S.; Li, K.; Gao, Y.; Liu, X.; Chen, W.; Ge, W.; Feng, Q.; Palli, S.R.; Li, S. Antagonistic actions of juvenile hormone and 20-hydroxyecdysone within the ring gland determine developmental transitions in Drosophila. Proc. Natl. Acad. Sci. USA 2018, 115, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Caragata, E.P.; Rances, E.; Hedges, L.M.; Gofton, A.W.; Johnson, K.N.; O’Neill, S.L.; McGraw, E.A. Dietary cholesterol modulates pathogen blocking by Wolbachia. PLoS Pathog. 2013, 9, e1003459. [Google Scholar] [CrossRef] [PubMed]
- Paredes, J.C.; Herren, J.K.; Schupfer, F.; Lemaitre, B. The role of lipid competition for endosymbiont-mediated protection against parasitoid wasps in Drosophila. mBio 2016, 7, e01006-16. [Google Scholar] [CrossRef]
- Cooper, M.K.; Wassif, C.A.; Krakowiak, P.A.; Taipale, J.; Gong, R.; Kelley, R.I.; Porter, F.D.; Beachy, P.A. A defective response to Hedgehog signaling in disorders of cholesterol biosynthesis. Nat. Genet. 2003, 33, 508–513. [Google Scholar] [CrossRef]
- Behmer, S.T.; Nes, W.D. Insect Sterol Nutrition and Physiology: A Global Overview; Elsevier: Amsterdam, The Netherlands, 2003; Volume 31, pp. 1–72. [Google Scholar]
- Ikekawa, N.; Morisaki, M.; Fujimoto, Y. Sterol metabolism in insects: Dealkylation of phytosterol to cholesterol. Phytochemistry 1993, 26, 485–486. [Google Scholar] [CrossRef]
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- Piironen, V.; Toivo, J.; Lampi, A.M. Plant sterols in cereals and cereal products. Cereal Chem. 2002, 79, 148–154. [Google Scholar] [CrossRef]
- Jing, X.F.; Grebenok, R.J.; Behmer, S.T. Diet micronutrient balance matters: How the ratio of dietary sterols/steroids affects development, growth and reproduction in two lepidopteran insects. J. Insect Physiol. 2014, 67, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T.; Elias, D.O. The nutritional significance of sterol metabolic constraints in the generalist grasshopper Schistocerca americana. J. Insect Physiol. 1999, 45, 339–348. [Google Scholar] [CrossRef]
- Ostlund, R.E., Jr. Phytosterols in human nutrition. Ann. Rev. Nutr. 2002, 22, 533–549. [Google Scholar] [CrossRef]
- Corio-Costet, M.F.; Charlet, M.; Benveniste, P.; Hoffman, J. Metabolism of dietary Δ8-sterols and 9β,19-cyclopropyl sterols by Locusta migratoria. Arch. Insect Biochem. 1989, 11, 47–62. [Google Scholar] [CrossRef]
- Chen, I.W.; Grebenok, R.J.; Schaller, H.; Zhu-Salzman, K.; Behmer, S.T. Aphid growth and reproduction on plants with altered sterol profiles: Novel insights using Arabidopsis mutant and overexpression lines. J. Insect Physiol. 2020, 123, 104054. [Google Scholar] [CrossRef]
- Heyer, J.; Parker, B.; Becker, D.; Ruffino, J.; Fordyce, A.; Witt, M.D.; Bedard, M.; Grebenok, R. Steroid profiles of transgenic tobacco expressing an Actinomyces 3-hydroxysteroid oxidase gene. Phytochemistry 2004, 65, 2967–2976. [Google Scholar] [CrossRef]
- Jing, X.F.; Grebenok, R.J.; Behmer, S.T. Sterol/steroid metabolism and absorption in a generalist and specialist caterpillar: Effects of dietary sterol/steroid structure, mixture and ratio. Insect Biochem. Mol. Biol. 2013, 43, 580–587. [Google Scholar] [CrossRef]
- Lang, M.; Murat, S.; Clark, A.G.; Gouppil, G.; Blais, C.; Matzkin, L.M.; Guittard, E.; Yoshiyama-Yanagawa, T.; Kataoka, H.; Niwa, R.; et al. Mutations in the neverland gene turned Drosophila pachea into an obligate specialist species. Science 2012, 337, 1658–1661. [Google Scholar] [CrossRef]
- Wang, W.W.; He, P.Y.; Zhang, Y.Y.; Liu, T.X.; Jing, X.F.; Zhang, S.Z. The population growth of Spodoptera frugiperda on six cash crop species and implications for its occurrence and damage potential in China. Insects 2020, 11, 639. [Google Scholar] [CrossRef] [PubMed]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Jia, J.; Sun, S.L.; Kuang, W.; Tang, R.; Zhang, Z.F.; Song, C.; Liu, T.X.; Jing, X. A semi-synthetic diet and the potential important chemicals for Mythimna separata (Lepidoptera: Noctuidae). J. Insect Sci. 2019, 19, 4. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.F.; Vogel, H.; Grebenok, R.J.; Zhu-Salzman, K.; Behmer, S.T. Dietary sterols/steroids and the generalist caterpillar Helicoverpa zea: Physiology, biochemistry and midgut gene expression. Insect Biochem. Mol. Bio. 2012, 42, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Corbin, D.R.; Greenplate, J.T.; Wong, E.Y.; Purcell, J.P. Cloning of an insecticidal cholesterol oxidase gene and its expression in bacteria and in plant protoplasts. Appl. Environ. Microb. 1994, 60, 4239–4244. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.F.; Grebenok, R.J.; Behmer, S.T. Plant sterols and host plant suitability for generalist and specialist caterpillars. J. Insect Physiol. 2012, 58, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Matyash, V.; Geier, C.; Henske, A.; Mukherjee, S.; Hirsh, D.; Maxfield, F.R.; Thiele, C.; Grant, B.; Kurzchalia, T.V. Distribution and transport of cholesterol in Caenorhabditis elegans. Mol. Biol. Cell 2001, 12, 1725–1736. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Park, M.S.; Hwang, S.; Hong, Y.K.; Choi, G.; Suh, Y.S.; Han, S.Y.; Kim, D.; Jeun, J.; Oh, C.T.; et al. Dietary hempseed meal intake increases body growth and shortens the larval stage via the upregulation of cell growth and sterol levels in Drosophila melanogaster. Mol. Cells 2010, 30, 29–36. [Google Scholar] [CrossRef]
- Clayton, R.B. The utilization of sterols by insects. J. Lipid Res. 1964, 5, 3–19. [Google Scholar] [CrossRef]
- Rodríguez-Acebes, S.; de la Cueva, P.; Fernandez-Hernando, C.; Ferruelo, A.J.; Lasuncion, M.A.; Rawson, R.B.; Martinez-Botas, J.; Gomez-Coronado, D. Desmosterol can replace cholesterol in sustaining cell proliferation and regulating the SREBP pathway in a sterol-Δ24-reductase-deficient cell line. Biochem. J. 2009, 420, 305–315. [Google Scholar] [CrossRef]
- Dutky, R.C.; Robbins, W.E.; Shortino, T.J.; Kaplanis, J.N.; Vroman, H.E. The conversion of cholestanone to cholestanol by the housefly, Musca domestica L. J. Insect Physiol. 1967, 13, 1501–1510. [Google Scholar] [CrossRef]
- Bouvaine, S.; Faure, M.L.; Grebenok, R.J.; Behmer, S.T.; Douglas, A.E. A dietary test of putative deleterious sterols for the aphid Myzus persicae. PLoS ONE 2014, 9, e86256. [Google Scholar] [CrossRef]
- Gilbert, L.I. Halloween genes encode P450 enzymes that mediate steroid hormone biosynthesis in Drosophila melanogaster. Mol. Cell. Endocrinol. 2004, 215, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sieber, M.H.; Thummel, C.S. Coordination of triacylglycerol and cholesterol homeostasis by DHR96 and the Drosophila LipA homolog magro. Cell Metab. 2012, 15, 122–127. [Google Scholar] [CrossRef]
- Jing, X.F.; Behmer, S.T. Insect sterol nutrition: Physiological mechanisms, ecology, and applications. Annu. Rev. Entomol. 2020, 65, 251–271. [Google Scholar] [CrossRef] [PubMed]
- Crowder, C.M.; Westover, E.J.; Kumar, A.S.; Ostlund, R.E., Jr.; Covey, D.F. Enantiospecificity of cholesterol function in vivo. J. Biol. Chem. 2001, 276, 44369–44372. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T.; Elias, D.O. Sterol metabolic constraints as a factor contributing to the maintenance of diet mixing in grasshoppers (Orthoptera: Acrididae). Physiol. Biochem. Zool. 2000, 73, 219–230. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mythimna separata | Spodoptera frugiperda | ||||||
|---|---|---|---|---|---|---|---|
| Dietary sterols | Cholesterol | Cholestanol | Cholestanone | Cholesterol | Cholestanol | Cholestanone | |
| Development time (day) | 23.21 ± 1.5 | 28 ± 1.32 * | — | 21.17 ± 1.7 | 30.71 ± 1.65 * | 31.67 ± 2.03 * | |
| Pupation rate (%) | 100 | 100 | 0 * | 100.0 | 100.0 | 79.17 ± 13.82 * | |
| Pupal mass (mg) | 432.67 ± 17.63 | 395.88 ± 30.57 * | — | 228.38 ± 11.43 | 202.83 ± 21.54 * | 209.33 ± 18.45 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, R.; Liang, J.; Jing, X.; Liu, T. Discrepancy in Sterol Usage between Two Polyphagous Caterpillars, Mythimna separata and Spodoptera frugiperda. Insects 2022, 13, 876. https://doi.org/10.3390/insects13100876
Tang R, Liang J, Jing X, Liu T. Discrepancy in Sterol Usage between Two Polyphagous Caterpillars, Mythimna separata and Spodoptera frugiperda. Insects. 2022; 13(10):876. https://doi.org/10.3390/insects13100876
Chicago/Turabian StyleTang, Rui, Junhao Liang, Xiangfeng Jing, and Tongxian Liu. 2022. "Discrepancy in Sterol Usage between Two Polyphagous Caterpillars, Mythimna separata and Spodoptera frugiperda" Insects 13, no. 10: 876. https://doi.org/10.3390/insects13100876