


Cuticle Modifications and Over-Expression of the Chitin-Synthase Gene in Diflubenzuron-Resistant Phenotype

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Establishment and Maintenance of Resistant and Susceptible Mosquito Colonies
2.2. DFB Bioassays
2.3. Expression Analysis of chs1 Gene
2.4. Cuticle Analysis
2.4.1. Analysis of Cuticle Thickness by Scanning Electron Microscopy (SEM)
2.4.2. Quantification of Chitin Content
3. Results
3.1. DFB Bioassay
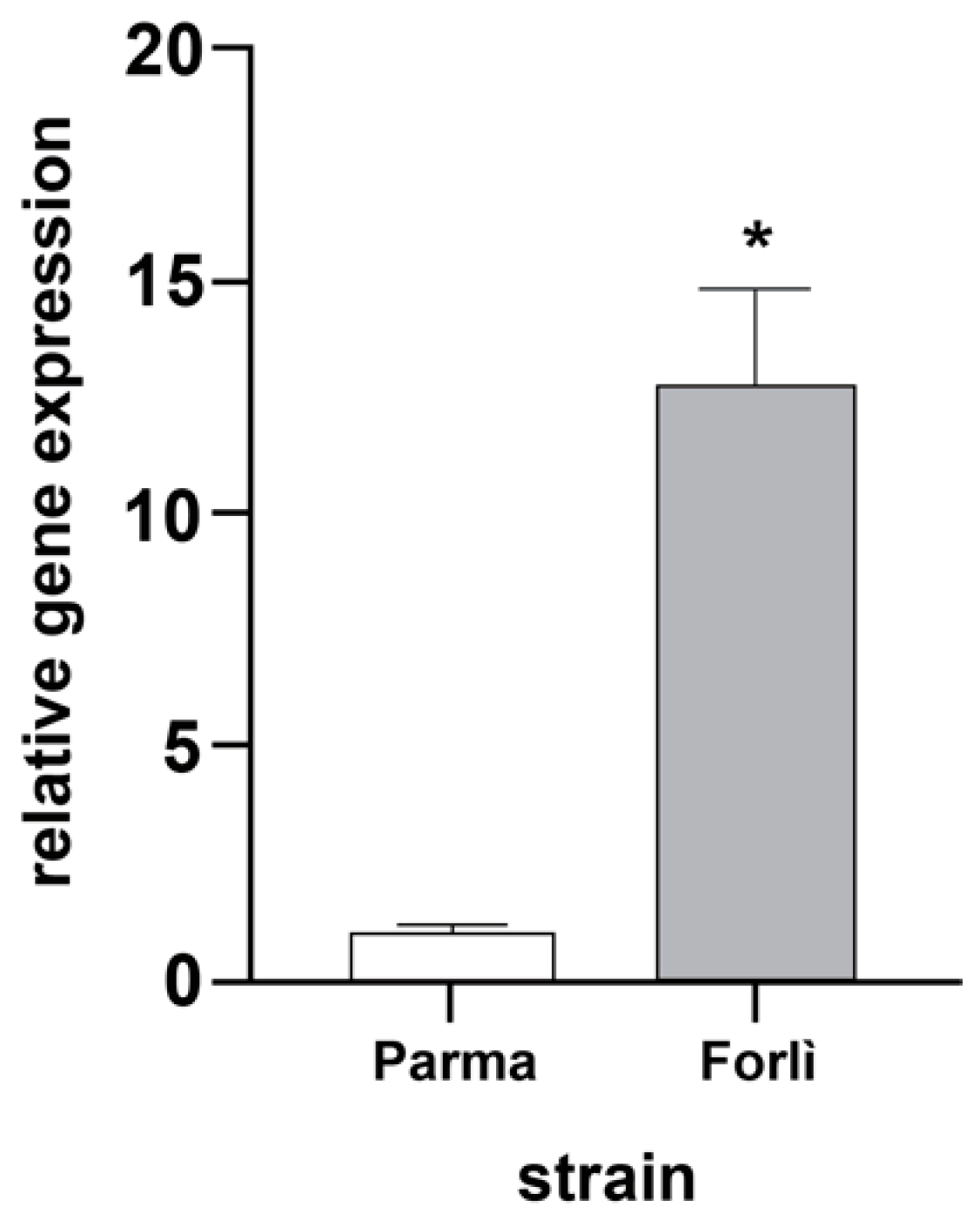
3.2. Expression of chs1 Gene in Resistant and Susceptible Cx. pipiens Mosquitoes
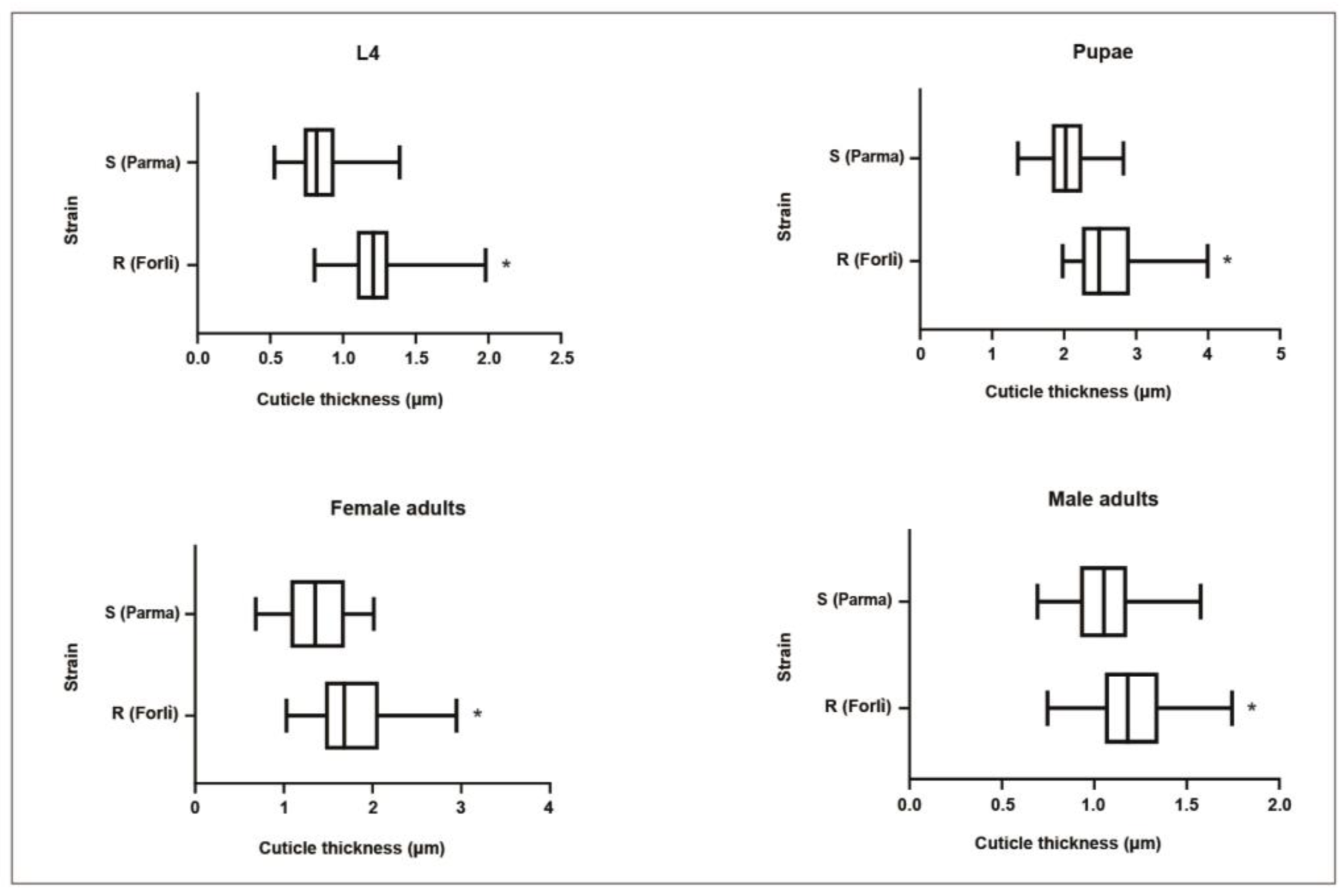
3.3. Cuticle Thickness
3.4. Quantification of Chitin Content
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-Lara, S.; Serna Saldivar, S.O. Insect Pests. In Encyclopedia of Food and Health; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- WHO. Global Vector Control Response 2017–2030; World Health Organization: Geneva, Switzerland, 2017.
- Whalon, M.E.; Mota-Sanchez, D.; Hollingworth, R.M. (Eds.) Global Pesticide Resistance in Arthropods; Cabi International: London, UK, 2008. [Google Scholar]
- Sparks, T.C.; Nauen, R. IRAC: Mode of Action Classification and Insecticide Resistance Management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Merzendorfer, H. Insect chitin synthases: A review. J. Comp. Physiol. B 2006, 176, 1–15. [Google Scholar] [CrossRef]
- Merzendorfer, H. Chitin synthesis inhibitors: Old molecules and new developments. Insect Sci. 2013, 20, 121–138. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: A review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Santos, V.C.; De Siqueira, H.A.A.; Da Silva, J.E.; De Farias, M.J.D.C. Insecticide resistance in populations of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae), from the state of Pernambuco, Brazil. Neotrop. Entomol. 2011, 40, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Ju, D.; Mota-Sanchez, D.; Fuentes-Contreras, E.; Zhang, Y.L.; Wang, X.Q.; Yang, X.Q. Insecticide resistance in the Cydia pomonella (L.): Global status, mechanisms, and research directions. Pestic. Biochem. Phys. 2021, 178, 104925. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Demaeght, P.; Osborne, E.J.; Dermauw, W.; Gohlke, S.; Nauen, R.; Grbić, M.; Tirry, L.; Merzendorfer, H.; Clark, R.M. Population bulk segregant mapping uncovers resistance mutations and the mode of action of a chitin synthesis inhibitor in arthropods. Proc. Natl. Acad. Sci. USA 2012, 109, 4407–4412. [Google Scholar] [CrossRef] [Green Version]
- Douris, V.; Steinbach, D.; Panteleri, R.; Livadaras, I.; Pickett, J.A.; Van Leeuwen, T.; Nauen, R.; Vontas, J. Resistance mutation conserved between insects and mites unravels the benzoylurea insecticide mode of action on chitin biosynthesis. Proc. Natl. Acad. Sci. USA 2016, 113, 14692–14697. [Google Scholar] [CrossRef] [Green Version]
- Grigoraki, L.; Puggioli, A.; Mavridis, K.; Douris, V.; Montanari, M.; Bellini, R.; Vontas, J. Striking diflubenzuron resistance in Culex pipiens, the prime vector of West Nile virus. Sci. Rep. 2017, 7, 11699. [Google Scholar] [CrossRef] [Green Version]
- Gomes, B.; Kioulos, E.; Papa, A.; Almeida, A.P.; Vontas, J.; Pinto, J. Distribution and hybridization of Culex pipiens forms in Greece during the West Nile virus outbreak of 2010. Infect. Genet. Evol. 2013, 16, 218–225. [Google Scholar] [CrossRef]
- Chaskopoulou, A.; L’Ambert, G.; Petric, D.; Bellini, R.; Zgomba, M.; Groen, T.A. Ecology of West Nile virus across four European countries: Review of weather profiles, vector population dynamics and vector control response. Parasit Vectors 2016, 9, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fotakis, E.A.; Mastrantonio, V.; Grigoraki, L.; Porretta, D.; Puggioli, A.; Chaskopoulou, A.; Osório, H.; Weill, M.; Bellini, R.; Urbanelli, S.; et al. Identification and detection of a novel point mutation in the Chitin Synthase gene of Culex pipiens associated with diflubenzuron resistance. PLoS Negl. Trop. Dis. 2020, 14, e0008284. [Google Scholar] [CrossRef] [PubMed]
- Guz, N.; Çağatay, N.S.; Fotakis, E.A.; Durmuşoğlu, E.; Vontas, J. Detection of diflubenzuron and pyrethroid resistance mutations in Culex pipiens from Muğla, Turkey. Acta Trop. 2020, 203, 105294. [Google Scholar] [CrossRef] [PubMed]
- Mastrantonio, V.; Porretta, D.; Lucchesi, V.; Güz, N.; Çağatay, N.S.; Bellini, R.; Vontas, J.; Urbanelli, S. Evolution of adaptive variation in the mosquito Culex pipiens: Multiple independent origins of insecticide resistance mutations. Insects 2021, 12, 676. [Google Scholar] [CrossRef]
- Porretta, D.; Fotakis, E.; Mastrantonio, V.; Chaskopoulou, A.; Michaelakis, A.; Kioulos, I.; Weill, M.; Urbanelli, S.; Vontas, J.; Bellini, R. Focal distribution of diflubenzuron resistance mutations in Culex pipiens mosquitoes from northern Italy. Acta Trop. 2019, 193, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Schaffner, F.; Angel, G.; Geoggroy, B.; Hervy, J.P.; Rhaiem, A.; Brunhes, J. The Mosquitoes of Europe/Les Moustiques d’Europe. Logiciel d’identification Et d’enseignement (CD-Rom); IRD Editions & EID Méditerranée: Montpellier, France, 2001. [Google Scholar]
- Robertson, J.A.; Preisler, H.K.; Russell, R.M. Polo Plus: Probit and Logit Analysis; LeOra Software: Petaluma, CA, USA, 2003. [Google Scholar]
- Vézilier, J.; Nicot, A.; De Lorgeril, J.; Gandon, S.; Rivero, A. The impact of insecticide resistance on Culex pipiens immunity. Evol. Appl. 2013, 6, 497–509. [Google Scholar] [CrossRef]
- Liu, N.; Li, T.; Reid, W.R.; Yang, T.; Zhang, L. Multiple Cytochrome P450 genes: Their constitutive overexpression and permethrin induction in insecticide resistant mosquitoes, Culex quinquefasciatus. PLoS ONE 2011, 6, e23403. [Google Scholar] [CrossRef]
- Timmermann, S.E.; Briegel, H. Larval growth and biosynthesis of reserves in mosquitoes. J. Insect Physiol. 1999, 45, 461–470. [Google Scholar] [CrossRef]
- Gunathilaka, P.A.; Uduwawala, U.M.; Udayanga, N.W.; Ranathunge, R.M.; Amarasinghe, L.D.; Abeyewickreme, W. Determination of the efficiency of diets for larval development in mass rearing Aedes aegypti (Diptera: Culicidae). Bull. Entomol. Res. 2018, 108, 583–592. [Google Scholar] [CrossRef]
- Yan, J.; Kibech, R.; Stone, C.M. Differential effects of larval and adult nutrition on female survival, fecundity, and size of the yellow fever mosquito, Aedes aegypti. Front. Zool. 2021, 18, 10. [Google Scholar] [CrossRef]
- Bhattacharya, R.; Saha, S.; Kostina, O.; Muravnik, M.; Mitra, A. Replacing critical point drying with a low-cost chemical drying provides comparable surface image quality of glandular trichomes from leaves of Millingtonia hortensis L. f. in scanning electron micrograph. Appl. Microsc. 2020, 50, 15. [Google Scholar] [CrossRef] [PubMed]
- Henriques, B.S.; Garcia, E.S.; Azambuja, P.; Genta, F.A. Determination of Chitin Content in Insects: An Alternate Method Based on Calcofluor Staining. Front. Physiol. 2020, 11, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, S.C.; Lockwood, J.L. Powdered chitin agar as a selective medium for enumeration of actinomycetes in water and soil. Appl. Microbiol. 1975, 29, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Porretta, D.; Mastrantonio, V.; Lucchesi, V.; Bellini, R.; Vontas, J.; Urbanelli, S. Historical samples reveal a combined role of agriculture and public-health applications in vector resistance to insecticides. Pest Manag. Sci. 2022, 78, 1567–1572. [Google Scholar] [CrossRef]
- Wood, O.R.; Hanrahan, S.; Coetzee, M.; Koekemoer, L.L.; Brooke, B.D. Cuticle thickening associated with pyrethroid resistance in the major malaria vector Anopheles funestus. Parasit Vectors 2010, 3, 67. [Google Scholar] [CrossRef] [Green Version]
- Lilly, D.G.; Latham, S.L.; Webb, C.E.; Doggett, S.L. Cuticle Thickening in a Pyrethroid-Resistant Strain of the Common Bed Bug, Cimex lectularius L. (Hemiptera: Cimicidae). PLoS ONE 2016, 11, e0153302. [Google Scholar] [CrossRef] [Green Version]
- Balabanidou, V.; Kampouraki, A.; MacLean, M.; Blomquist, G.J.; Tittiger, C.; Juárez, M.P.; Mijailovsky, S.J.; Chalepakis, G.; Anthousi, A.; Lynd, A.; et al. Cytochrome P450 associated with insecticide resistance catalyzes cuticular hydrocarbon production in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2016, 113, 9268–9273. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Guo, Q.; Sun, X.; Zhang, C.; Xu, N.; Xu, Y.; Zhou, D.; Sun, Y.; Ma, L.; Zhu, C.; et al. Culex pipiens pallens cuticular protein CPLCG5 participates in pyrethroid resistance by forming a rigid matrix. Parasit Vectors 2018, 11, 6. [Google Scholar] [CrossRef]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef]
- Muthukrishnan, S.; Mun, S.; Noh, M.Y.; Geisbrecht, E.R.; Arakane, Y. Insect Cuticular Chitin Contributes to Form and Function. Curr. Pharm. Des. 2020, 26, 3530–3545. [Google Scholar] [CrossRef]
- Andersen, S.O. Biochemistry of the insect cuticle. Annu. Rev. Entomol. 1979, 24, 29–61. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Fragment Length (bp) | Reference |
|---|---|---|---|
| G6PDH (HK) |
F: CGCGCACGAGGAAAAGTACG
R: GGTTTGCGGTCTTCCCAACC | 131 | [21] |
| 18S (HK) |
F: CGCGGTAATTCCAGCTCCACTA
R: GCATCAAGCGCCACATATAGG | 159 | [22] |
| chs1 |
F: GCCTGTCTCCATCCGCAAG
R: CCCAGGAGACGACGTTCAG | 124 | [12] |
| Strain | LD50 (95% CI) | LD90 (95% CI) | RRLD50 † |
|---|---|---|---|
| Cp_S | 0.005 mgL−1 (0.002–0.008) | 0.018 mgL−1 (0.012–0.031) | 1 |
| Cp_R-I1043M | 45.03 mgL−1 (13.06–60.66) | 85.81 mgL−1 (69.56–125.85) | 9006 |
| Strain | N | Developmental Stages | |||
|---|---|---|---|---|---|
| L4 Larvae | Pupae | Male Adults | Female Adults | ||
| Cp_S | 150 | 0.86 (±0.19) | 2.05 (±0.29) | 1.06 (±0.18) | 1.37 (±0.33) |
| Cp_R-I1043M | 150 | 1.22 (±0.21) * | 2.62(±0.19) * | 1.21 (±0.18) * | 1.78 (±0.41) * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucchesi, V.; Grimaldi, L.; Mastrantonio, V.; Porretta, D.; Di Bella, L.; Ruspandini, T.; Di Salvo, M.L.; Vontas, J.; Bellini, R.; Negri, A.; et al. Cuticle Modifications and Over-Expression of the Chitin-Synthase Gene in Diflubenzuron-Resistant Phenotype. Insects 2022, 13, 1109. https://doi.org/10.3390/insects13121109
Lucchesi V, Grimaldi L, Mastrantonio V, Porretta D, Di Bella L, Ruspandini T, Di Salvo ML, Vontas J, Bellini R, Negri A, et al. Cuticle Modifications and Over-Expression of the Chitin-Synthase Gene in Diflubenzuron-Resistant Phenotype. Insects. 2022; 13(12):1109. https://doi.org/10.3390/insects13121109
Chicago/Turabian StyleLucchesi, Valentina, Lorenzo Grimaldi, Valentina Mastrantonio, Daniele Porretta, Letizia Di Bella, Tania Ruspandini, Martino Luigi Di Salvo, John Vontas, Romeo Bellini, Agata Negri, and et al. 2022. "Cuticle Modifications and Over-Expression of the Chitin-Synthase Gene in Diflubenzuron-Resistant Phenotype" Insects 13, no. 12: 1109. https://doi.org/10.3390/insects13121109
APA StyleLucchesi, V., Grimaldi, L., Mastrantonio, V., Porretta, D., Di Bella, L., Ruspandini, T., Di Salvo, M. L., Vontas, J., Bellini, R., Negri, A., Epis, S., Caccia, S., Bandi, C., & Urbanelli, S. (2022). Cuticle Modifications and Over-Expression of the Chitin-Synthase Gene in Diflubenzuron-Resistant Phenotype. Insects, 13(12), 1109. https://doi.org/10.3390/insects13121109