

The Association between the Distribution of Resin Beads and the Emergence of Sirex noctilio on Red Pine in North America

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
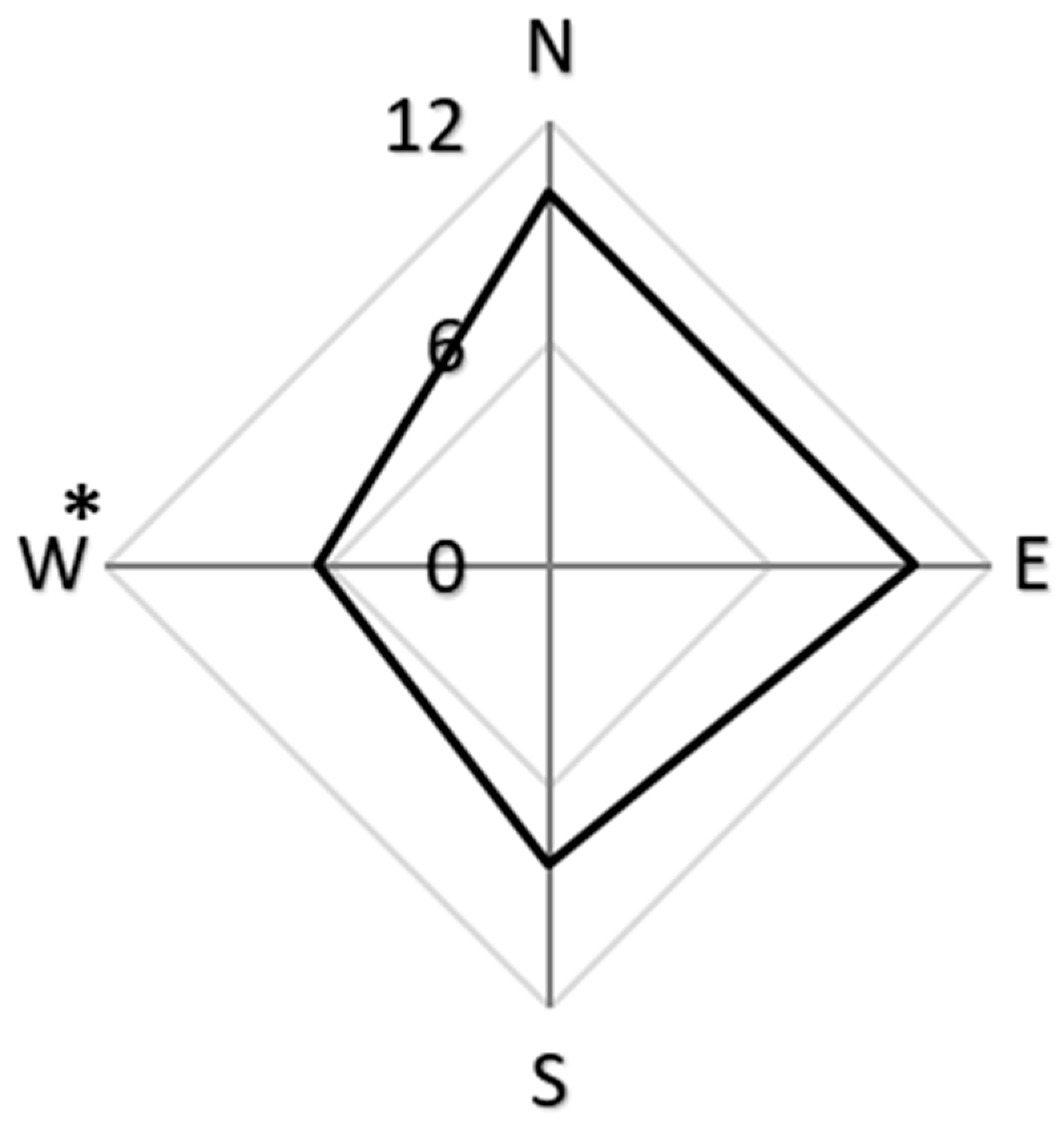
2.1. Distribution of Resin Beads
2.2. Distribution of Insects
2.3. Data Analysis
2.3.1. Distribution of Resin Beads
2.3.2. Distribution of Insects
3. Results
3.1. Distribution of Resin Beads
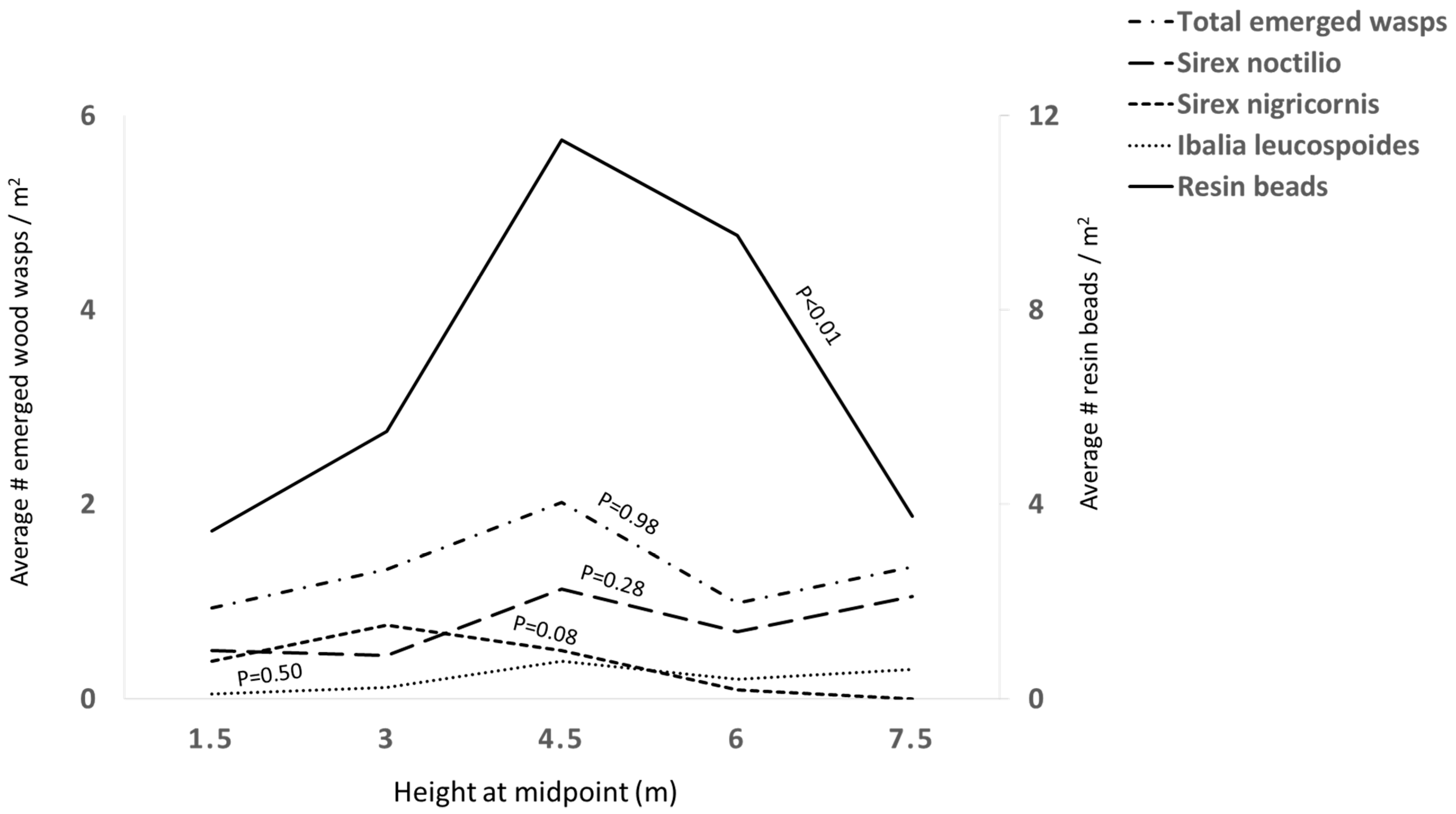
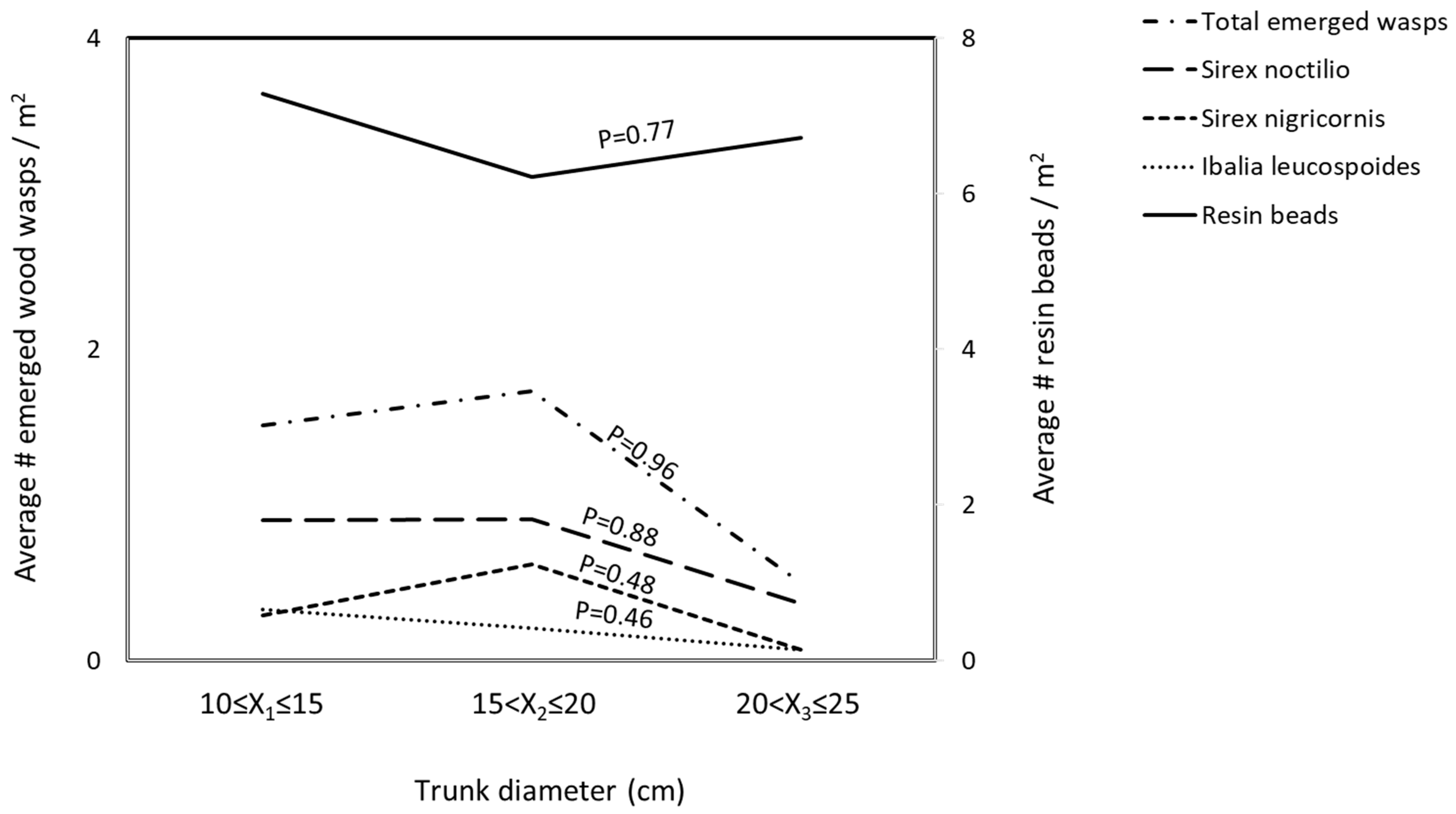
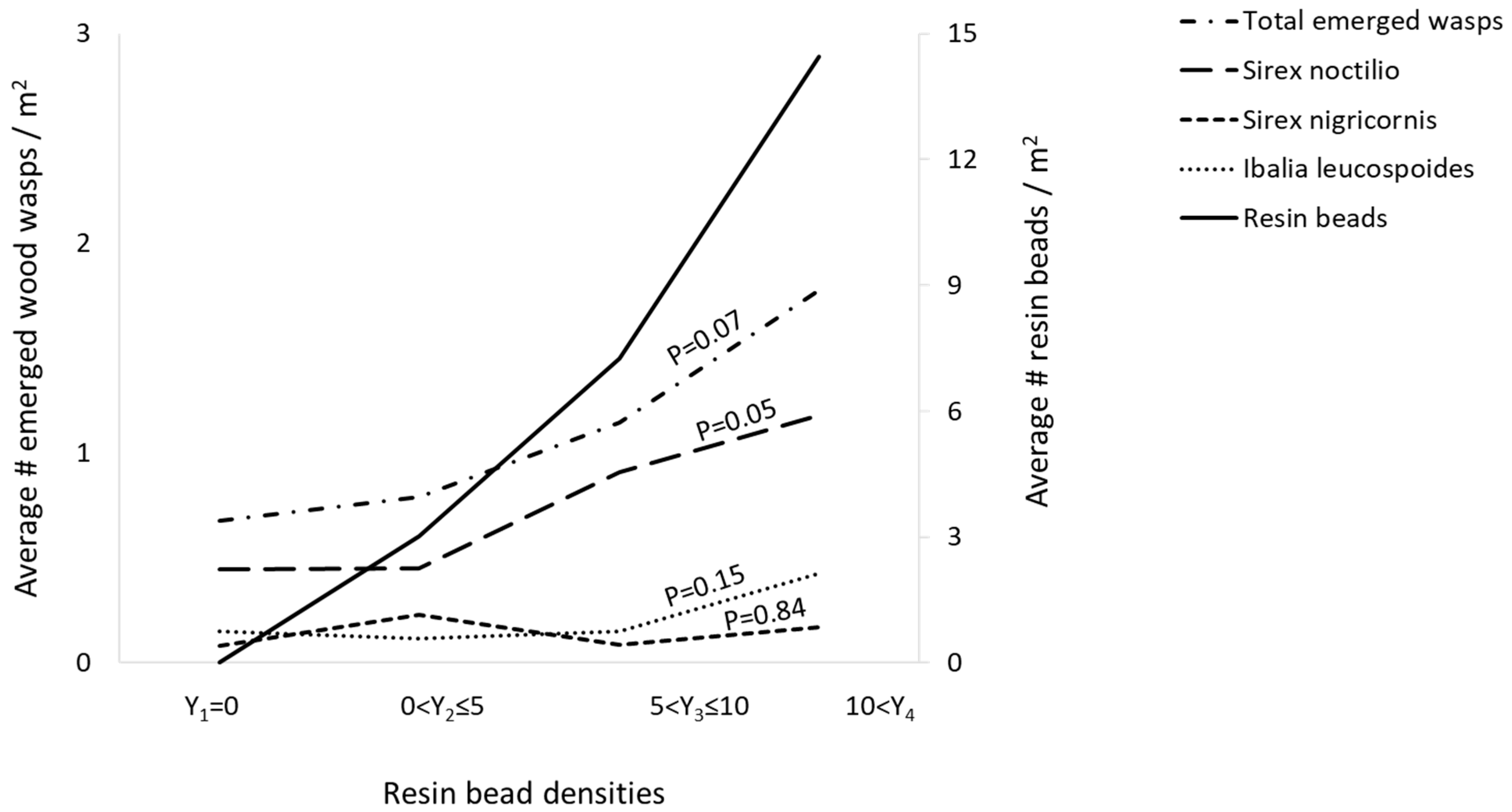
3.2. Distribution of Insects
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slippers, B.; Hurley, B.P.; Wingfield, M.J. Sirex woodwasp: A model for evolving management paradigms of invasive forest pests. Annu. Rev. Entomol. 2015, 60, 601–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, J.L. Physiological reactions of Pinus radiata to attack by woodwasp, Sirex noctilio F.(Hymenoptera: Siricidae). Bull. Entomol. Res. 1977, 67, 405–426. [Google Scholar] [CrossRef]
- Böröczky, K.; Zylstra, K.E.; McCartney, N.B.; Mastro, V.C.; Tumlinson, J.H. Volatile profile differences and the associated Sirex noctilio activity in two host tree species in the northeastern United States. J. Chem. Ecol. 2012, 38, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Ferrenberg, S.; Langenhan, J.M.; Loskot, S.A.; Rozal, L.M.; Mitton, J.B. Resin monoterpene defenses decline within three widespread species of pine (Pinus) along a 1530-m elevational gradient. Ecosphere 2017, 8, e01975. [Google Scholar] [CrossRef]
- Hudgins, J.W.; Christiansen, E.; Franceschi, V.R. Induction of anatomically based defense responses in stems of diverse conifers by methyl jasmonate: A phylogenetic perspective. Tree Physiol. 2004, 24, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Coutts, M.P.; Dolezal, J.E. Polyphenols and Resin in the Resistance Mechanisms of Pinus Radiata Attacked by the Wood Wasp, Sirex Noctilio, and Its Associated Fungus; Department of National Development, Forestry and Timber Bureau, Australia, 1966. Department of National Development, Forestry and Timber Bureau: Hawthorn, VIC, Australia.
- Madden, J.L.; Coutts, M.P. The role of fungi in the biology and ecology of woodwasps (Hymenoptera: Siricidae). In Insect–Fungus Symbiosis; Allanheld, Osmun & Co.: Montclair, NJ, USA, 1979; pp. 165–174. [Google Scholar]
- Madden, J.L. Oviposition behaviour of the woodwasp, Sirex noctilio F. Aust. J. Zool. 1974, 22, 341–351. [Google Scholar] [CrossRef]
- Madden, J.L.; Irvine, C.J. The use of lure trees for the detection of Sirex noctilio in the field. Aust. For. 1971, 35, 164–166. [Google Scholar] [CrossRef]
- Slippers, B.; De Groot, P.; Wingfield, M.J. The Sirex Woodwasp and its Fungal Symbiont: Research and Management of a Worldwide Invasive Pest. Springer: Dordrecht, The Netherlands, 2011. [Google Scholar]
- Wermelinger, B.; Rigling, A.; Schneider Mathis, D.; Dobbertin, M. Assessing the role of bark- and wood-boring insects in the decline of Scots pine (Pinus sylvestris) in the Swiss Rhone valley. Ecol. Entomol. 2008, 33, 239–249. [Google Scholar] [CrossRef]
- Hurley, B.P.; Slippers, B.; Wingfield, M.J.; Dyer, C.; Slippers, J. Perception and Knowledge of the Sirex Woodwasp and Other Forest Pests in South Africa. Agric. For. Entomol. 2012, 14, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.; de Groot, P.; Davis, C.; Smith, S.M. Effect of two bark beetle-vectored fungi on the on-host search and oviposition behavior of the introduced woodwasp Sirex noctilio (Hymenoptera: Siricidae) on Pinus sylvestris trees and logs. J. Insect Behav. 2012, 25, 453–466. [Google Scholar] [CrossRef]
- Ryan, K.; de Groot, P.; Smith, S.M. Evidence of interaction between Sirex noctilio and other species inhabiting the bole of Pinus. Agric. For. Entomol. 2012, 14, 187–195. [Google Scholar] [CrossRef]
- Yousuf, F.; Gurr, G.M.; Carnegie, A.J.; Bedding, R.A.; Bashford, R.; Gitau, C.W.; Nicol, H.I. The bark beetle, Ips grandicollis, disrupts biological control of the woodwasp, Sirex noctilio, via fungal symbiont interactions. FEMS Microbiol. Ecol. 2014, 88, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousuf, F.; Carnegie, A.J.; Bashford, R.; Bedding, R.A.; Nicol, H.I.; Gurr, G.M. Bark beetle (Ips grandicollis) disruption of woodwasp (Sirex noctilio) biocontrol: Direct and indirect mechanisms. For. Ecol. Manag. 2014, 323, 98–104. [Google Scholar] [CrossRef]
- Haavik, L.J.; Dodds, K.J.; Allison, J.D. Do native insects and associated fungi limit non-native woodwasp, Sirex noctilio, survival in a newly invaded environment? PLoS ONE 2015, 10, e0138516. [Google Scholar] [CrossRef] [PubMed]
- Bittner, T.D.; Hajek, A.E.; Liebherr, J.K. Associations among Serropalpus substriatus (Coleoptera: Melandryidae) and Sirex (Hymenoptera: Siricidae) Communities. Great Lakes Entomol. 2016, 49, 18–26. [Google Scholar]
- Clarke, C.W.; Carnegie, A.J.; Yousuf, F.; Bashford, R.; Nicol, H.I.; Bedding, R.A.; Gurr, G.M. Minimizing the disruptive effect of Ips grandicollis (Coleoptera: Scolytinae) on biocontrol of Sirex noctilio (Hymenoptera: Siricidae). For. Ecol. Manag. 2016, 381, 134–143. [Google Scholar] [CrossRef]
- Hajek, A.E.; Tobin, P.C.; Kroll, S.A.; Long, S.J. Symbionts mediate oviposition behaviour in invasive and native woodwasps. Agric. For. Entomol. 2018, 20, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Madden, J.L. Sirex in Australasia. In Dynamics of Forest Insect Populations; Plenum Pub. Corp.: New York, NY, USA, 1988; pp. 407–429. [Google Scholar]
- Carnegie, A.J.; Bashford, R. Sirex woodwasp in Australia: Current management strategies, research and emerging issues. In The Sirex woodwasp and Its Fungal Symbiont; Springer: Dordrecht, The Netherlands, 2012; pp. 175–201. [Google Scholar]
- Williams, D.W.; Zylstra, K.E.; Mastro, V.C. Ecological considerations in using Deladenus (=Beddingia) siricidicola for the biological control of Sirex noctilio in North America. In The Sirex woodwasp and Its Fungal Symbiont; Springer: Dordrecht, The Netherlands, 2012; pp. 135–148. [Google Scholar]
- Thompson, B.M.; Bodart, J.; Gruner, D.S. Community resistance to an invasive forest insect–fungus mutualism. Ecosphere 2019, 10, e02609. [Google Scholar] [CrossRef]
- Carnegie, A.J.; Eldridge, R.H.; Waterson, D.G. History and management of Sirex woodwasp in pine plantations in New South Wales, Australia. N. Z. J. For. Sci. 2005, 35, 3–24. [Google Scholar]
- Fischbein, D.; Corley, J.C. Classical biological control of an invasive forest pest: A world perspective of the management of Sirex noctilio using the parasitoid Ibalia leucospoides (Hymenoptera: Ibaliidae). Bull. Entomol. Res. 2015, 105, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Faal, H.; Cha, D.H.; Hajek, A.E.; Teale, S.A. A double-edged sword: Amylostereum areolatum odors attract both Sirex noctilio (Hymenoptera: Siricidae) and its parasitoid, Ibalia leucospoides. Fungal Ecol. 2021, 54, 101108. [Google Scholar] [CrossRef]
- Faal, H.; Silk, P.J.; Mayo, P.D.; Teale, S.A. Courtship behavior and identification of a sex pheromone in Ibalia leucospoides (Hymenoptera: Ibaliidae), a larval parasitoid of Sirex noctilio (Hymenoptera: Siricidae). PeerJ 2021, 9, e12266. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Arhex, V.; Corley, J.C. The functional response of Ibalia leucospoides (Hymenoptera: Ibaliidae), a parasitoid of Sirex noctilio (Hymenoptera: Siricidae). Biocontrol Sci. Technol. 2005, 15, 207–212. [Google Scholar] [CrossRef]
- Hurley, B.P.; Fitza, K.N.E.; Wingfield, M.J.; Slippers, B. Sequence data reflect the introduction pathways of the Sirex woodwasp parasitoid, Ibalia leucospoides (Ibaliidae, Hymenoptera). Agric. For. Entomol. 2020, 22, 129–135. [Google Scholar] [CrossRef]
- Bragg, D.C. An improved tree height measurement technique tested on mature southern pines. South. J. Appl. For. 2008, 32, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Coutts, M.P. Sirex noctilio and the Physiology of Pinus radiata: Some Studies of Interactions between the Insect, the Fungus and the Tree in Tasmania. Department of National Development, Forestry and Timber Bureau: Canberra, ACT, Austrilia, 1965. [Google Scholar]
- Madden, J.L. Physiological aspects of host tree favourability for the woodwasp, Sirex noctilio F. In Proceedings of the Ecological Society of Australia; 1968; pp. 147–149. [Google Scholar]
- Chapman, J.A. Field studies on attack flight and log selection by the ambrosia beetle Trypodendron lineatum (Oliv.)(Coleoptera: Scolytidae). Can. Entomol. 1962, 94, 74–92. [Google Scholar] [CrossRef]
- Byers, J.A. Upwind flight orientation to pheromone in western pine beetle tested with rotating wind vane traps. J. Chem. Ecol. 1988, 14, 189–198. [Google Scholar] [CrossRef]
- Safranyik, L.; Silversides, R.; McMullen, L.H.; Linton, D.A. An empirical approach to modeling the local dispersal of the mountain pine beetle (Dendroctonus ponderosae Hopk.) (Col., Scolytidae) in relation to sources of attraction, wind direction and speed. J. Appl. Entomol. 1989, 108, 498–511. [Google Scholar] [CrossRef]
- Safranyik, L.; Linton, D.A.; Silversides, R.; McMullen, L.H. Dispersal of released mountain pine beetles under the canopy of a mature lodgepole pine stand. J. Appl. Entomol. 1992, 113, 441–450. [Google Scholar] [CrossRef]
- Mahroof, R.M.; Edde, P.A.; Robertson, B.; Puckette, J.A.; Phillips, T.W. Dispersal of Rhyzopertha dominica (Coleoptera: Bostrichidae) in different habitats. Environ. Entomol. 2010, 39, 930–938. [Google Scholar] [CrossRef] [Green Version]
- Madden, J.L. An analysis of an outbreak of the woodwasp, Sirex noctilio F. (Hymenoptera, Siricidae), in Pinus radiata. Bull. Entomol. Res. 1975, 65, 491–500. [Google Scholar] [CrossRef]
- Ryan, K.; de Groot, P.; Smith, S.M.; Turgeon, J.J. Seasonal occurrence and spatial distribution of resinosis, a symptom of Sirex noctilio (Hymenoptera: Siricidae) injury, on boles of Pinus sylvestris (Pinaceae). Can. Entomol. 2013, 145, 117–122. [Google Scholar] [CrossRef]
- Dodds, K.J.; de Groot, P.; Orwig, D.A. The impact of Sirex noctilio in Pinus resinosa and Pinus sylvestris stands in New York and Ontario. Can. J. For. Res. 2010, 40, 212–223. [Google Scholar] [CrossRef]
- Dodds, K.J.; Cooke, R.R.; Gilmore, D.W. Silvicultural options to reduce pine susceptibility to attack by a newly detected invasive species, Sirex noctilio. North. J. Appl. For. 2007, 24, 165–167. [Google Scholar] [CrossRef] [Green Version]
- Rudolf, P.O. Red pine (Pinus resinosa Ait.). Silv. N. Am. 1990, 1, 442–455. [Google Scholar]
- McLaughlin, J.A.; Hsiang, T.; Hayden, G.H.; Greifenhagen, S. Abiotic and biotic factors used to assess decline risk in red pine (Pinus resinosa Ait.) plantations. For. Chron. 2011, 87, 99–115. [Google Scholar] [CrossRef] [Green Version]
- Mosseler, A.; Egger, K.N.; Hughes, G.A. Low levels of genetic diversity in red pine confirmed by random amplified polymorphic DNA markers. Can. J. For. Res. 1992, 22, 1332–1337. [Google Scholar] [CrossRef]
- Standley, C.R. Emergence Phenology and Ecological Interactions between the Exotic Sirex noctilio, Native Siricids, and a Shared Guild of Native Parasitoids. Master’s Thesis, State University of New York College of Environmental Science and Forestry, Syracuse, NY, USA, 2012. [Google Scholar]
- Albert, J.; Platek, M.; Cizek, L. Vertical stratification and microhabitat selection by the Great Capricorn Beetle (Cerambyx cerdo) (Coleoptera: Cerambycidae) in open-grown, veteran oaks. Eur. J. Entomol. 2012, 109, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Kašák, J.; Foit, J. Shortage of declining and damaged sun-exposed trees in European mountain forests limits saproxylic beetles: A case study on the endangered longhorn beetle Ropalopus ungaricus (Coleoptera: Cerambycidae). J. Insect Conserv. 2018, 22, 171–181. [Google Scholar] [CrossRef]
- Bereczki, K.; Molnár, D.; Csóka, G.; Báldi, A. Factors affecting the bird predation of low density gypsy moth egg masses in three types of hardwood forests in southwest Hungary. Bull. Insectol. 2017, 70, 201–207. [Google Scholar]
- Zargaran, M.R.; Safaralizadeh, M.H.; Pourmirza, A.A.; Valizadegan, O. Effect of cardinal directions on gall morphology and parasitization of the gall wasp, Cynips quercusfolii. J. Insect Sci. 2011, 11, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoeller, E.N. Characterization of the Subcortical Interactions between Larvae of the Southern Pine Sawyer Monochamus titillator (F.) and the Larvae of the Southern Pine Beetle Guild Using Molecular Gut Analyses. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2011. [Google Scholar]
- Thompson, B.M.; Bodart, J.; McEwen, C.; Gruner, D.S. Adaptations for symbiont-mediated external digestion in Sirex noctilio (Hymenoptera: Siricidae). Ann. Entomol. Soc. Am. 2014, 107, 453–460. [Google Scholar] [CrossRef]
- Foelker, C.J.; Standley, C.R.; Parry, D.; Fierke, M.K. Complex ecological relationships among an assemblage of indigenous hymenopteran parasitoids, the exotic European woodwasp (Sirex noctilio; Hymenoptera: Siricidae), and a native congener. Can. Entomol. 2016, 148, 532–542. [Google Scholar] [CrossRef]
- Bookwalter, J.D.; Riggins, J.J.; Dean, J.F.D.; Mastro, V.C.; Schimleck, L.R.; Sullivan, B.T.; Gandhi, K.J.K. Colonization and development of Sirex noctilio (Hymenoptera: Siricidae) in bolts of a native pine host and six species of pine grown in the southeastern United States. J. Entomol. Sci. 2019, 54, 1–18. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Emerged | Total Emerged Wasps | Total Unemerged Wasps | ||
|---|---|---|---|---|---|
| Sirex noctilio | Sirex nigricornis | Ibalia leucospoides | |||
| Resin beads | |||||
| β 1 | 0.48 | −0.07 | 0.48 | 0.35 | 0.19 |
| F | 4.05 | 0.04 | 2.21 | 3.41 | 0.20 |
| P | 0.05 * | 0.84 | 0.15 | 0.07 | 0.66 |
| Df 2 | 1, 37 | 1, 37 | 1, 37 | 1, 37 | 1, 37 |
| Height | |||||
| β | 0.16 | −0.40 | 0.14 | 0.02 | −0.07 |
| F | 1.19 | 3.34 | 0.46 | 0.00 | 0.06 |
| P | 0.28 | 0.08 | 0.50 | 0.98 | 0.81 |
| Df | 1, 37 | 1, 37 | 1, 37 | 1, 37 | 1, 37 |
| Diameter | |||||
| β | 0.08 | −0.53 | −0.38 | 0.03 | −0.37 |
| F | 0.02 | 0.49 | 0.54 | 0.00 | 0.30 |
| P | 0.88 | 0.48 | 0.46 | 0.96 | 0.59 |
| Df | 1, 37 | 1, 37 | 1, 37 | 1, 37 | 1, 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faal, H.; Teale, S.A. The Association between the Distribution of Resin Beads and the Emergence of Sirex noctilio on Red Pine in North America. Insects 2022, 13, 1111. https://doi.org/10.3390/insects13121111
Faal H, Teale SA. The Association between the Distribution of Resin Beads and the Emergence of Sirex noctilio on Red Pine in North America. Insects. 2022; 13(12):1111. https://doi.org/10.3390/insects13121111
Chicago/Turabian StyleFaal, Hajar, and Stephen A. Teale. 2022. "The Association between the Distribution of Resin Beads and the Emergence of Sirex noctilio on Red Pine in North America" Insects 13, no. 12: 1111. https://doi.org/10.3390/insects13121111
APA StyleFaal, H., & Teale, S. A. (2022). The Association between the Distribution of Resin Beads and the Emergence of Sirex noctilio on Red Pine in North America. Insects, 13(12), 1111. https://doi.org/10.3390/insects13121111