
Enterococci as Dominant Xylose Utilizing Lactic Acid Bacteria in Eri Silkworm Midgut and the Potential Use of Enterococcus hirae as Probiotic for Eri Culture

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
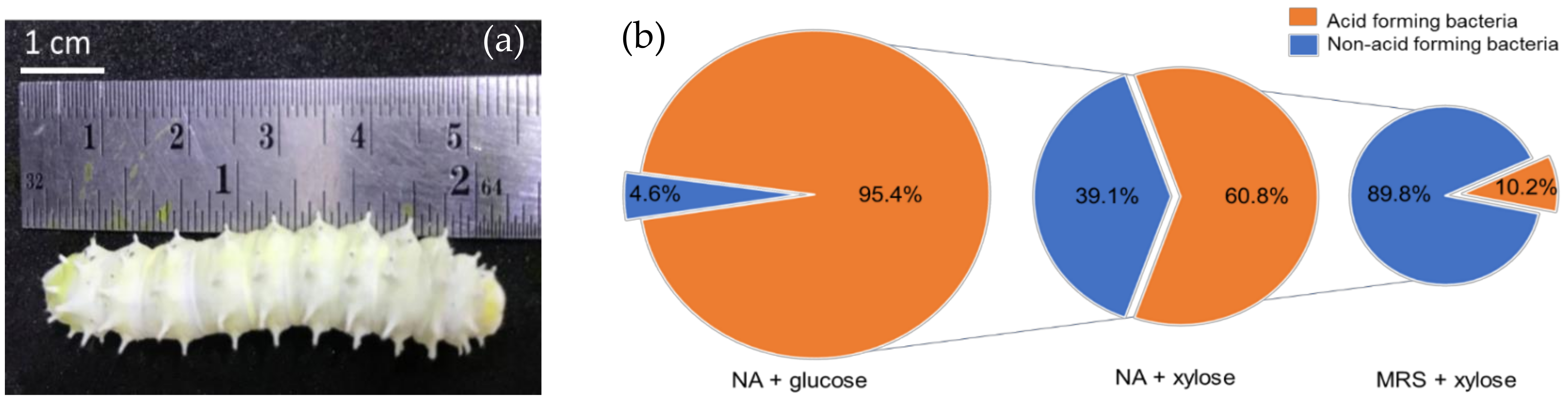
2.1. Isolation and Screening of Xylose Utilizing LAB
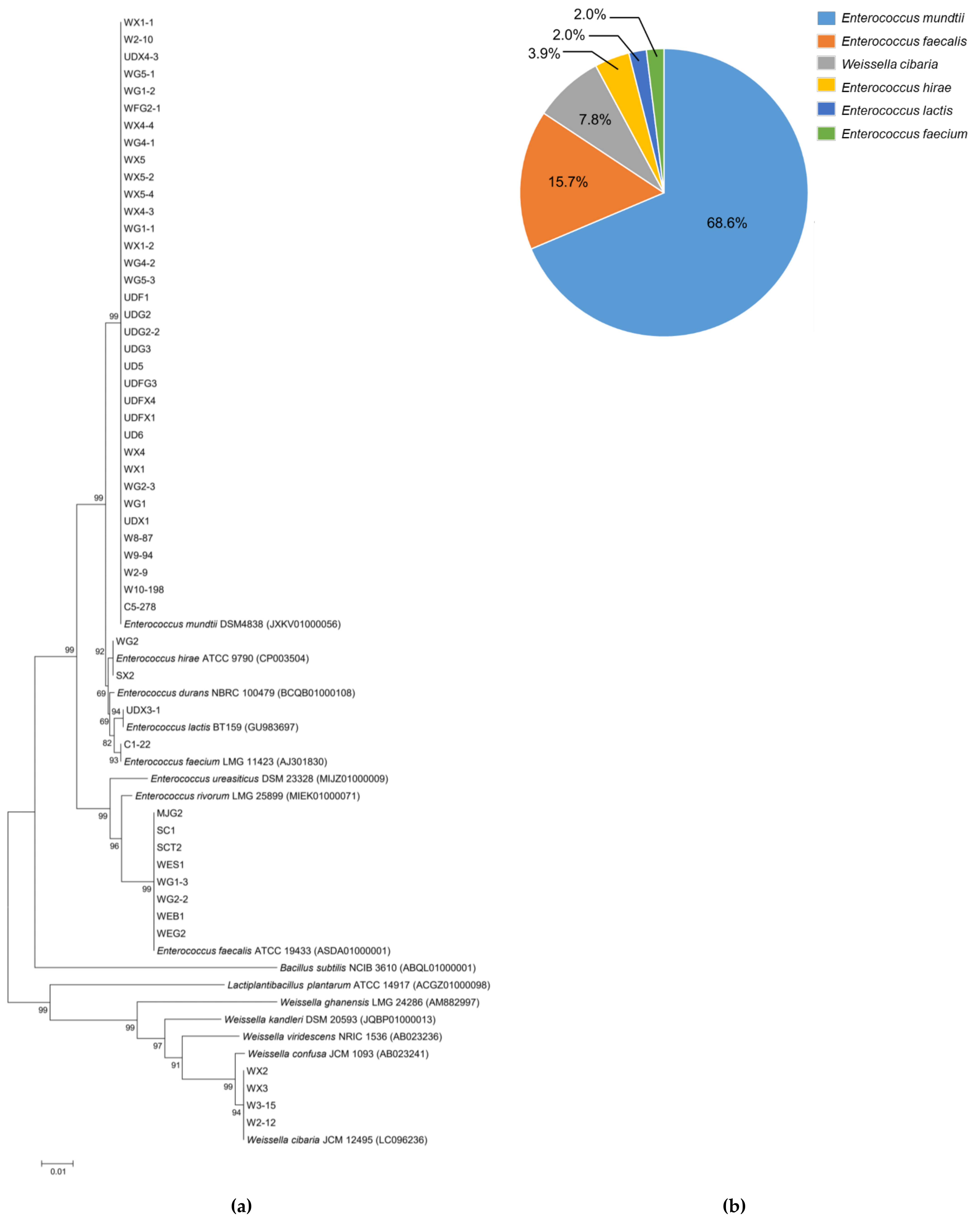
2.2. Identification of LAB by 16S rDNA Sequence Analysis
2.3. Growth and Acid Formation on Polysaccharides
2.4. Tannin-Tolerance Test
2.5. Antimicrobial Activity against Insect Pathogens
2.6. Effect of E. hirae SX2 Supplementation on Eri Silkworm Growth
2.7. Statistical Analysis
3. Results and Discussion
3.1. Xylose Utilizing LAB Isolation
3.2. Xylose Utilizing LAB Identification and Phylogenetic Analysis
3.3. Growth and Acid Formation on Polysaccharide Substrates
3.4. Tannin-Tolerance Test
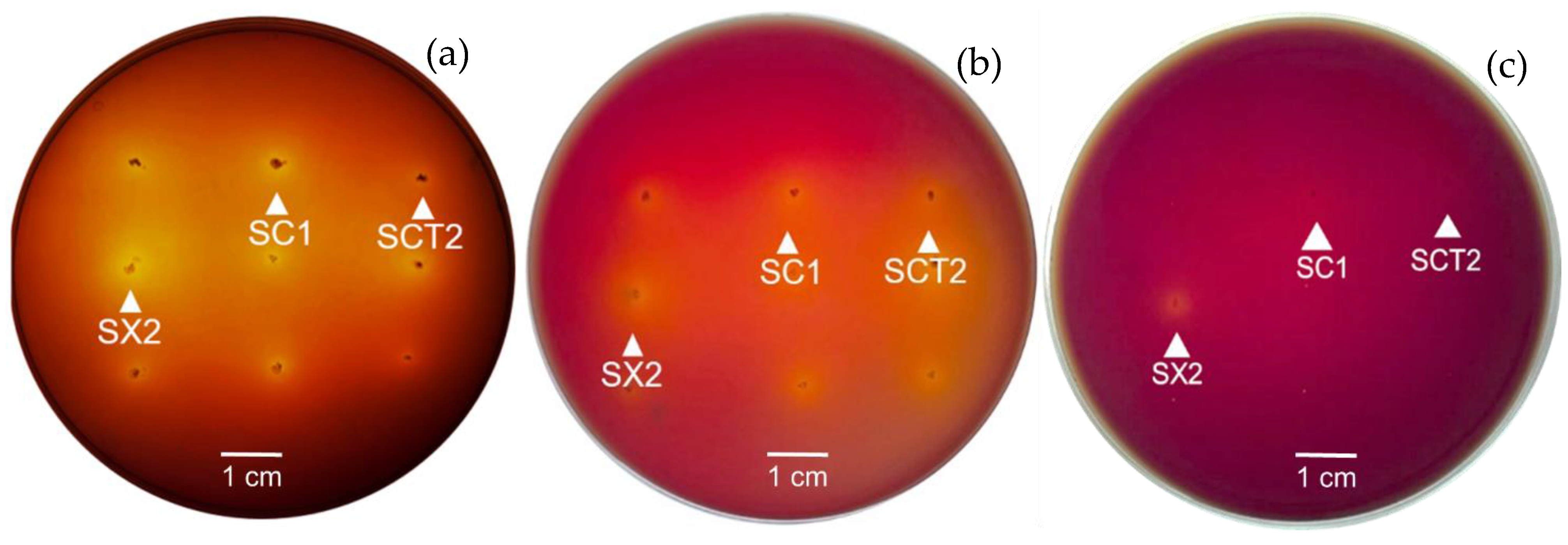
3.5. Antimicrobial Activity
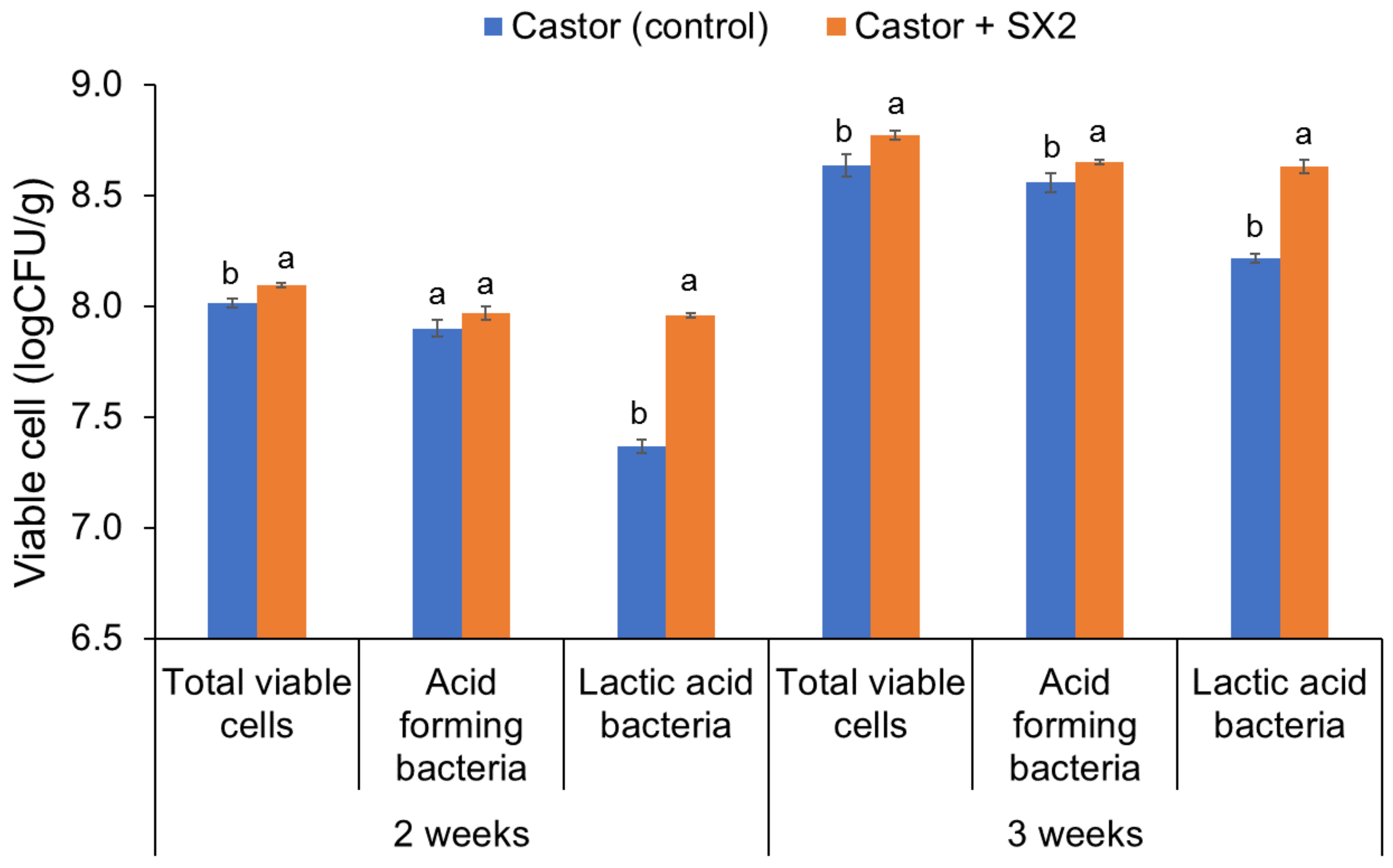
3.6. Effect of E. hirae SX2 Supplementation on Eri Silkworm Growth
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pallavi, S.B. Commercial characters of selected eco-races of eri silkworm (Samia cynthia ricini Boisduval) reared on castor hybrid/variety. Inter. J. Res. Anal. Rev. 2018, 5, 513–518. [Google Scholar]
- Charungkiattikul, S.; Joneurairatana, E. A revival for thailand’s textile traditions: New value for local materials (eri silk) through art practice. Textile 2021, 1–14. [Google Scholar] [CrossRef]
- Leamkaew, V.; Jitjankarn, P.; Chairat, M. Polydopamine-dyed eri silk yarn for the improvement of wash and light fastness properties. J. Text. Inst. 2021, 112, 553–560. [Google Scholar] [CrossRef]
- Prabhakar, C.S.; Sood, P.; Kanwar, S.S.; Sharma, P.N.; Kumar, A.; Mehta, P.K. Isolation and characterization of gut bacteria of fruit fly, Bactrocera tau (Walker). Phytoparasitica 2013, 41, 193–201. [Google Scholar] [CrossRef]
- Paniagua Voirol, L.R.; Frago, E.; Kaltenpoth, M.; Hilker, M.; Fatouros, N.E. Bacterial symbionts in Lepidoptera: Their diversity, transmission, and impact on the host. Front. Microbiol. 2018, 9, 556. [Google Scholar] [CrossRef] [PubMed]
- Nandy, G.; Chakraborti, M.; Shee, A.; Aditya, G.; Acharya, K. Gut microbiota from lower groups of animals: An upcoming source for cellulolytic enzymes with industrial potentials. Biointerface Res. Appl. Chem. 2021, 11, 13614–13637. [Google Scholar]
- Ceja-Navarro, J.A.; Vega, F.E.; Karaoz, U.; Hao, Z.; Jenkins, S.; Lim, H.C.; Kosina, P.; Infante, F.; Northen, T.R.; Brodie, E.L. Gut microbiota mediate caffeine detoxification in the primary insect pest of coffee. Nat. Commun. 2015, 6, 7618. [Google Scholar] [CrossRef] [Green Version]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Lactobacillus plantarum promotes Drosophila systemic growth by modulating hormonal signals through TOR-dependent nutrient sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Naser, A.Z.; Deiab, I.; Darras, B.M. Polylactic acid (PLA) and polyhydroxyalkanoates (PHAs), green alternatives to petroleum-based plastics: A review. RSC Adv. 2021, 11, 17151–17196. [Google Scholar] [CrossRef]
- Kim, J.-H.; Block, D.E.; Mills, D.A. Simultaneous consumption of pentose and hexose sugars: An optimal microbial phenotype for efficient fermentation of lignocellulosic biomass. Appl. Microbiol. Biotechnol. 2010, 88, 1077–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, P.; Bai, M. Studies on the effect of aprobiotic and a neutraceutical agent on growth, development and commercial characteristics of silkworm, Bombyx mori L. Indian J. Seric. 2012, 51, 37–42. [Google Scholar]
- Dantur, K.I.; Enrique, R.; Welin, B.; Castagnaro, A.P. Isolation of cellulolytic bacteria from the intestine of Diatraea saccharalis larvae and evaluation of their capacity to degrade sugarcane biomass. AMB Express 2015, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mala, M.; Vijila, K. Beneficial effects of Bacillus licheniformis and Bacillus niabensis on growth and economic characteristics of silkworm, Bombyx mori L. Int. J. Chem. Stud. 2018, 6, 1750–1754. [Google Scholar]
- Prem Anand, A.A.; Vennison, S.J.; Sankar, S.G.; Gilwax Prabhu, D.I.; Vasan, P.T.; Raghuraman, T.; Jerome Geoffrey, C.; Vendan, S.E. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J. Insect Sci. 2010, 10, 107. [Google Scholar]
- Quinto, E.J.; Jiménez, P.; Caro, I.; Tejero, J.; Mateo, J.; Girbés, T. Probiotic lactic acid bacteria: A review. Food Nutr. Sci. 2014, 5, 1765. [Google Scholar] [CrossRef] [Green Version]
- Sonnenburg, J.L.; Chen, C.T.L.; Gordon, J.I. Genomic and metabolic studies of the impact of probiotics on a model gut symbiont and host. PLoS Biol. 2006, 4, e413. [Google Scholar] [CrossRef] [Green Version]
- Unban, K.; Kodchasee, P.; Shetty, K.; Khanongnuch, C. Tannin-tolerant and extracellular tannase producing Bacillus isolated from traditional fermented tea leaves and their probiotic functional properties. Foods 2020, 9, 490. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Fehlberg, L.C.C.; Nicoletti, A.G.; Ramos, A.C.; Rodrigues-Costa, F.; de Matos, A.P.; Girardello, R.; Marques, E.A.; Gales, A.C. In vitro susceptibility of Burkholderia cepacia complex isolates: Comparison of disk diffusion, Etest®, agar dilution, and broth microdilution methods. Diagn. Microbiol. Infect. Dis. 2016, 86, 422–427. [Google Scholar] [CrossRef] [PubMed]
- MsangoSoko, K.; Gandotra, S.; Chandel, R.K.; Sharma, K.; Ramakrishinan, B.; Subramanian, S. Composition and diversity of gut bacteria associated with the eri silk moth, Samia ricini, (Lepidoptera: Saturniidae) as revealed by culture-dependent and metagenomics analysis. J. Microbiol. Biotechnol. 2020, 30, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Yeruva, T.; Vankadara, S.; Ramasamy, S.; Lingaiah, K. Identification of potential probiotics in the midgut of mulberry silkworm, Bombyx mori through metagenomic approach. Probiotics Antimicro. Prot. 2020, 12, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Lu, Y.; Zhang, H.; Kumar, D.; Liu, B.; Gong, Y.; Zhu, M.; Zhu, L.; Liang, Z.; Kuang, S. Effects of BmCPV infection on silkworm Bombyx mori intestinal bacteria. PLoS ONE 2016, 11, e0146313. [Google Scholar] [CrossRef]
- Chen, B.; Teh, B.-S.; Sun, C.; Hu, S.; Lu, X.; Boland, W.; Shao, Y. Biodiversity and activity of the gut microbiota across the life history of the insect herbivore Spodoptera littoralis. Sci. Rep. 2016, 6, 29505. [Google Scholar] [CrossRef]
- Masthan, K.; Rajkumar, T.; Narasimha Murthy, C.V. Studies on fortification of mulberry leaves with probiotics for improvement of silk quality. Int. J. Biotechlol. Biochem. 2017, 13, 73–80. [Google Scholar]
- Moustafa, M.N.; Soliman, S. Nutritional efficiency and economic traits of silkworm Bombyx mori, L. reared on mulberry leaves fortified with synbiotics. J. Plant Prot. Pathol. 2019, 10, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Yu, T.; Xie, S.; Du, K.; Liang, X.; Lan, Y.; Sun, C.; Lu, X.; Shao, Y. Comparative shotgun metagenomic data of the silkworm Bombyx mori gut microbiome. Sci. Data 2018, 5, 1–10. [Google Scholar] [CrossRef]
- Chen, B.; Du, K.; Sun, C.; Vimalanathan, A.; Liang, X.; Li, Y.; Wang, B.; Lu, X.; Li, L.; Shao, Y. Gut bacterial and fungal communities of the domesticated silkworm (Bombyx mori) and wild mulberry-feeding relatives. ISME J. 2018, 12, 2252–2262. [Google Scholar] [CrossRef] [Green Version]
- Klongklaew, A.; Unban, K.; Kanpiengjai, A.; Wongputtisin, P.; Pamueangmun, P.; Shetty, K.; Khanongnuch, C. Improvement of enantiomeric L-lactic acid production from mixed hexose-pentose sugars by coculture of Enterococcus mundtii WX1 and Lactobacillus rhamnosus SCJ9. Fermentation 2021, 7, 95. [Google Scholar] [CrossRef]
- Pandiarajan, J.; Revathy, K. Cellulolytic potential of gut bacterial biomass in silkworm Bombyx mori. L. Ecol. Genet. Genom. 2020, 14, 100045. [Google Scholar] [CrossRef]
- Shil, R.K.; Mojumder, S.; Sadida, F.F.; Uddin, M.; Sikdar, D. Isolation and identification of cellulolytic bacteria from the gut of three phytophagus insect species. Braz. Arch. Biol. Technol. 2014, 57, 927–932. [Google Scholar] [CrossRef]
- Despres, L.; David, J.-P.; Gallet, C. The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 2007, 22, 298–307. [Google Scholar] [CrossRef]
- Buzzini, P.; Arapitsas, P.; Goretti, M.; Branda, E.; Turchetti, B.; Pinelli, P.; Ieri, F.; Romani, A. Antimicrobial and antiviral activity of hydrolysable tannins. Mini-Rev. Med. Chem. 2008, 8, 1179. [Google Scholar] [CrossRef] [PubMed]
- Gandotra, S.; Kumar, A.; Naga, K.; Bhuyan, P.; Gogoi, D.; Sharma, K.; Subramanian, S. Bacterial community structure and diversity in the gut of the muga silkworm, Antheraea assamensis (Lepidoptera: Saturniidae), from India. Insect Mol. Biol. 2018, 27, 603–619. [Google Scholar] [CrossRef]
- Bhalchandra, P.M.; Pathade, G. Antibacterial and cholesterol reducing lactic acid bacteria from silk worm (Bombyx mori) gut environment—A review. Nat. Environ. Pollut. Technol. 2011, 10, 319–326. [Google Scholar]
- Anitha, J.; Sathish, J.; Sujatha, K. Nutritional efficiency and economic traits in Samia ricini donovan reared on castor leaves fortified with probiotic agent. Int. J. Inn. Res. Sci. Eng. Tech 2015, 4, 2319–8753. [Google Scholar]
- Shruti, A.; Hadimani, D.; Sreenivas, A.; Beladhadi, R. Effect of probiotic feed supplements to mulberry silkworm, Bombyx mori L. for larval growth and development parameters. Int. J. Chem. Stud. 2019, 7, 3914–3919. [Google Scholar]
- Singh, K.; Chauhan, R.; Pande, A.; Gokhale, S.; Hegde, N. Effect of use of Lactobacillus plantarum as a probiotics to improve cocoon production of mulberry silkworm Bombyx mori (L). J. Basic Appl. Sci. 2005, 1, 1–8. [Google Scholar]
- Suraporn, S.; Sangsuk, W.; Chanhan, P.; Promma, S. Effects of probiotic bacteria on the growth parameters of the Thai silkworm, Bombyx mori. Thai J. Agric. Sci. 2015, 48, 29–33. [Google Scholar]
- Shao, Y.; Chen, B.; Sun, C.; Ishida, K.; Hertweck, C.; Boland, W. Symbiont-derived antimicrobials contribute to the control of the Lepidopteran gut microbiota. Cell Chem. Biol. 2017, 24, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Zhan, M.; Evivie, S.E.; Jin, D.; Zhao, L.; Chowdhury, S.; Sarker, S.K.; Huo, G.; Liu, F. Evaluating the safety of potential probiotic Enterococcus durans KLDS6. 0930 using whole genome sequencing and oral toxicity study. Front. Microbiol. 2018, 9, 1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I. Probiotic potential and safety evaluation of Enterococcus faecalis OB14 and OB15, isolated from traditional tunisian testouri cheese and rigouta, using physiological and genomic analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Fu, Y.; Liu, H. Isolation and characterization of enzyme-producing bacteria of the silkworm larval gut in bioregenerative life support system. Acta Astronaut. 2015, 116, 247–253. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Inhibition Zones against Pathogenic Bacteria (mm) | |||||
|---|---|---|---|---|---|---|
| Proteus vulgaris | Bacillus cereus | Streptococcus aureus | ||||
| Unneutralized CFCS | Neutralized CFCS | Unneutralized CFCS | Neutralized CFCS | Unneutralized CFCS | Neutralized CFCS | |
| C1-22 | ++ | ++ | + | + | ND | ND |
| C5-278 | ++ | ++ | + | + | ND | ND |
| MJG2 | ++ | ++ | + | + | ND | ND |
| SC1 | ++ | ++ | + | + | ND | ND |
| SCT2 | +++ | +++ | ++ | + | ++ | + |
| SX2 | +++ | +++ | ++ | ++ | ++ | + |
| UD5 | +++ | +++ | + | + | ND | ND |
| UD6 | ++ | ++ | + | + | ND | ND |
| UDF1 | +++ | +++ | ++ | + | + | ND |
| UDFG3 | + | + | + | + | ND | ND |
| UDFX1 | +++ | +++ | ++ | + | + | ND |
| UDFX4 | ++ | ++ | + | + | ND | ND |
| UDG2 | ++ | ++ | + | + | ND | ND |
| UDG2-2 | ++ | ++ | + | + | ND | ND |
| UDG3 | +++ | +++ | ++ | + | + | ND |
| UDX1 | ++ | ++ | + | + | ND | ND |
| UDX3-1 | ++ | ++ | + | + | ND | ND |
| UDX4-3 | ++ | ++ | + | + | ND | ND |
| W10-198 | ++ | ++ | + | + | ND | ND |
| W2-10 | ++ | ++ | + | + | ND | ND |
| W2-12 | ++ | ++ | + | + | ND | ND |
| W2-9 | ++ | ++ | + | + | ND | ND |
| W3-15 | ++ | ++ | + | + | ND | ND |
| W8-87 | ++ | ++ | + | + | ND | ND |
| W9-94 | ++ | ++ | + | + | ND | ND |
| WEB1 | ++ | ++ | + | + | ND | ND |
| WEG2 | ++ | ++ | + | + | ND | ND |
| WES1 | +++ | +++ | + | + | ND | ND |
| WFG2-1 | + | + | + | + | ND | ND |
| WG1 | ++ | ++ | + | + | ND | ND |
| WG1-1 | ++ | ++ | + | + | ND | ND |
| WG1-2 | + | + | + | + | ND | ND |
| WG1-3 | + | + | + | + | ND | ND |
| WG2 | ++ | ++ | + | + | ND | ND |
| WG2-2 | ++ | ++ | + | + | ND | ND |
| WG2-3 | + | + | + | + | ND | ND |
| WG4-1 | + | + | + | + | ND | ND |
| WG4-2 | ++ | ++ | + | + | ND | ND |
| WG5-1 | + | + | + | + | ND | ND |
| WG5-3 | + | + | + | + | ND | ND |
| WX1 | + | + | + | + | ND | ND |
| WX1-1 | + | + | + | + | ND | ND |
| WX1-2 | ++ | ++ | + | + | ND | ND |
| WX2 | +++ | +++ | ++ | + | ND | ND |
| WX3 | +++ | +++ | ++ | + | ND | ND |
| WX4 | +++ | +++ | + | + | ND | ND |
| WX4-3 | +++ | +++ | + | + | ND | ND |
| WX4-4 | ++ | ++ | + | + | ND | ND |
| WX5 | ++ | ++ | + | + | ND | ND |
| WX5-2 | ++ | ++ | + | + | ND | ND |
| WX5-4 | ++ | ++ | + | + | ND | ND |
| Treatments | 0 Week (1st Instar Larva) | 1 Week | 2 Weeks | 3 Weeks |
|---|---|---|---|---|
| Castor | 0.03 ± 0.04 | 0.48 ± 0.03 a | 1.53 ± 0.04 b | 4.20 ± 0.04 b |
| Castor + SX2 | 0.02 ± 0.02 | 0.47 ± 0.01 a | 1.69 ± 0.09 a | 4.53 ± 0.07 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unban, K.; Klongklaew, A.; Kodchasee, P.; Pamueangmun, P.; Shetty, K.; Khanongnuch, C. Enterococci as Dominant Xylose Utilizing Lactic Acid Bacteria in Eri Silkworm Midgut and the Potential Use of Enterococcus hirae as Probiotic for Eri Culture. Insects 2022, 13, 136. https://doi.org/10.3390/insects13020136
Unban K, Klongklaew A, Kodchasee P, Pamueangmun P, Shetty K, Khanongnuch C. Enterococci as Dominant Xylose Utilizing Lactic Acid Bacteria in Eri Silkworm Midgut and the Potential Use of Enterococcus hirae as Probiotic for Eri Culture. Insects. 2022; 13(2):136. https://doi.org/10.3390/insects13020136
Chicago/Turabian StyleUnban, Kridsada, Augchararat Klongklaew, Pratthana Kodchasee, Punnita Pamueangmun, Kalidas Shetty, and Chartchai Khanongnuch. 2022. "Enterococci as Dominant Xylose Utilizing Lactic Acid Bacteria in Eri Silkworm Midgut and the Potential Use of Enterococcus hirae as Probiotic for Eri Culture" Insects 13, no. 2: 136. https://doi.org/10.3390/insects13020136