
An Interactive Teaching Tool Describing Resistance Evolution and Basic Economics of Insecticide-Based Pest Management


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
- Carrying capacity (K): A theoretical pest population equilibrium density for a given “cropping universe” (i.e., crop field, greenhouse, orchard, etc.). A carrying capacity value is needed for the inclusion of density-dependent population dynamics. As default, K = 1000. However, it can be changed when applied to specific case stories of economics of pest management strategies, in which density of crop stand (and therefore insect pest carrying capacity) is assessed [15,16]. It may also be changed if pest population densities are expected to vary markedly among seasons as a function of crop management practices (i.e., crop rotation) and/or due to fluctuating seasonal weather conditions. Finally, the carrying capacity may be set to a specific value if it is known that a given crop field equals a certain number of plants per hectare and it is known that each crop plant can host a certain number of insect pest individuals.
- Resistance allele frequency: The initial frequency of allele(s) conferring insecticide resistance. With two alleles, initial frequencies of SS, SR, and RR genotypes are calculated based on assumption of the Hardy–Weinberg equilibrium. For simplicity, only two alleles and three genotypes are included. If three or more alleles or multiple genes are assumed to be involved and to maintain assumption of Hardy–Weinberg equilibrium, the six or more possible genotypes may be grouped into low, medium, high levels of (polygenic and/or metabolic) resistance levels [17]. In other words, this simple model structure can be used to examine and visualize expected outcomes from scenarios that are more complex than those involving only one gene and two alleles.
- Survival (W): Average survival of each genotype in response to insecticide treatment (0 = no survival and 1 = 100% survival). That is, insect pests have different survival rates for each developmental stage, but for simplicity, a single average survival coefficient for all life stages is used.
- Net reproductive rate (R): The natural logarithm of the offspring/adults ratio from one generation to the next, and a value of Ln(5) is proposed as default. Generally speaking, a high R coefficient shifts temporal trends left-wards (resistance evolution after few generations), while a low R coefficient shift temporal trends right-wards (resistance evolution rate).
- Fitness cost (F): A large body of literature describes how the evolution of insecticide resistance may be associated with a wide range of fitness costs [10,14,18,19,20,21,22,23,24]. For simplicity, only a single model parameter is included to encompass the total relative reduction in net reproductive rate (loss of fitness) due to insecticide resistance (0 = no fitness cost and 1 = complete loss of reproductive rate).
- Emigration (E): Number of pest individuals leaving the given cropping universe at the end of each generation. Emigration is calculated based on a fixed percentage of population density in the previous generation, N, of each genotype. Thus, if E = 10, then 10% of pest individuals of that genotype will emigrate. The fixed E-value for each generation does not take into account that pest individuals may leave at different time periods during the growing season and, therefore, cause different levels of crop damage. Moreover, the fixed E-value should be considered an average emigration rate for each generation.
- Immigration (I): It is assumed that insect pest individuals in the given cropping universe originate from a “global pest population” with a fixed genotypic composition. The genotypic composition of this global pest population is described by resistance allele frequency (cell C3 in the interactive teaching tool) and by assumption of the Hardy–Weinberg equilibrium. Thus, the genotypic composition of insect pest individuals during initial immigration as well as during immigration in each of the 30 generations is based on these fixed assumptions. The actual number of pest individuals immigrating into the given cropping universe in each generation is calculated based on a fixed percentage, I, of the carrying capacity, K. Thus, if I = 10 and K = 1000, then 100 insect pest individuals immigrate into the cropping universe each generation. Furthermore, these 100 immigrating insect pest individuals in each generation have a genotypic composition based on cell C3 in the interactive teaching tool. The effect of immigration is added to the model before a reduction in survival (W) imposed by insecticide treatment. The fixed I-value for each generation does not take into account that pest individuals may arrive at different time periods during the growing season and, therefore, cause different levels of crop damage. Moreover, the fixed I-value should be considered an average immigration rate for each generation. The fixed I-value may also be viewed as representing the effect of a non-insecticidal refuge in efforts to avoid/delay resistance evolution [25].
- Economic injury level (EIL): As originally stated by Stern, Smith, Bosch and Hagen [29] and discussed in detail by Pedigo, Hutchins and Higley [26], EIL is defined as “the lowest population density that will cause economic damage”. In cell C10 in the interactive tool, an EIL value is: 0 ≤ EIL ≤ K. If EIL = 0, then mortality imposed by insecticide treatment (and selection pressure), W, is applied in all 30 discrete and univoltine generations. If EIL = K, then no mortality imposed by insecticide treatment will occur in ant of the 30 discrete and univoltine generations. An EIL of about 2-10% of K may be realistic starting point when developing and comparing model scenarios. For simplicity, it is assumed that pest population density and crop damage are directly (and linearly) correlated. As reviewed and discussed elsewhere in great detail [26], there are many important reasons for this correlation not to be considered linear and to vary among locations and growing seasons. However, as an introduction to basic economics of insecticide-based pest management, a linear and constant (fixed average for all life stages) response may in many cases be considered an acceptable assumption [26]. Accordingly, loss of survival, W, is only invoked in a given generation if total pest population density exceeds the user-defined EIL. In the interactive teaching tool, it is assumed that only a single insect pest is present, and it is exclusively responsible for all potential crop damage and therefore yield loss. If scenarios involving Bt-transgenic (Bacillus thuringiensis) crops are examined, then it would be assumed that mortality imposed by Bt toxins in crop plants is constitutive, so an EIL = 0 should be used.
- Damage rate: The relationship between insect pest density and crop damage (loss) is, for simplicity, assumed to be linear, and the slope must be >0. It is one of the most important quantitative aspects of insect pest management, and it was the main topic of a seminal article [26]. In cases where insect pests feed directly on what is harvested (i.e., fruits and seeds), direct crop damage is typically calculated as a direct function of the number of pest individuals. However, accurate calculation of the damage rate becomes considerably more complicated when insect pests cause indirect crop damage, which is also referred to as crop injury. Moreover, leaf herbivores or root-feeders cause stress of crop plants, so that plant injury indirectly leads to loss of yield. Additionally, the yield effect of such indirect crop damage varies as a function of crop stage and of which part of crop plants is injured, and there may be certain levels or intensities of injury leading to crop damage (damage boundary) [26]. Thus, many pest–crop scenarios encompass rather complex and frequently non-linear associations of insect pest density with injury, crop damage, and crop loss. Here, these associations have been markedly simplified, and a single and fixed parameter, damage rate (encompassing both crop injury and loss), describes the linear increase in crop damage as a function of insect pest density.
- Treatment cost (in US Dollars): total insecticide treatment costs of insecticide, labor, and use of equipment. It may also represent the relative cost of planting Bt-transgenic crops versus non-Bt crops. This parameter is used in well-described calculations of pest density action thresholds [27]. If the cropping universe is measured in hectares or acres, a large body of applied research articles and websites can provide specific information about the costs of insecticide treatment costs. Treatment cost does not take into account the secondary effects of insecticide treatments. Thus, possible adverse effects on natural enemies, outbreaks, or suppression of secondary pests are not factored into the simple economic calculations.
- Profit potential (USD): the potential value of crop harvest under the assumption of zero crop damage due to the specific pest and after excluding all non-insecticidal operational production costs. For simplicity, loss of profit as a function of insect pest-induced crop damage is assumed to be linear. Such a simplification does not apply to crop harvests, in which quality grading exists, and/or when a given harvest has to meet certain minimum specifications to be sold on niche—or restrictive—export markets. Finally, such a linear association between profit potential and crop damage does not take into account the possibility of damage by one insect pest potentially increasing the risks of secondary pest issues, such as higher risks of mycotoxins in moth-infested grain or nuts [30]. Thus, the basic economics portion of the interactive teaching tool does not include many of the important reasons why more advanced agricultural economic models are needed for reliable predictions of specific pest–crop systems. However, as general and basic introduction, it was found acceptable to use a highly simplified model structure and to assume linear association between crop damage and profit potential. In column M in the Excel spreadsheet, a logical statement is used to ensure that estimated profit potential cannot be negative.
3. Results
3.1. How to Use the Interactive Teaching Tool
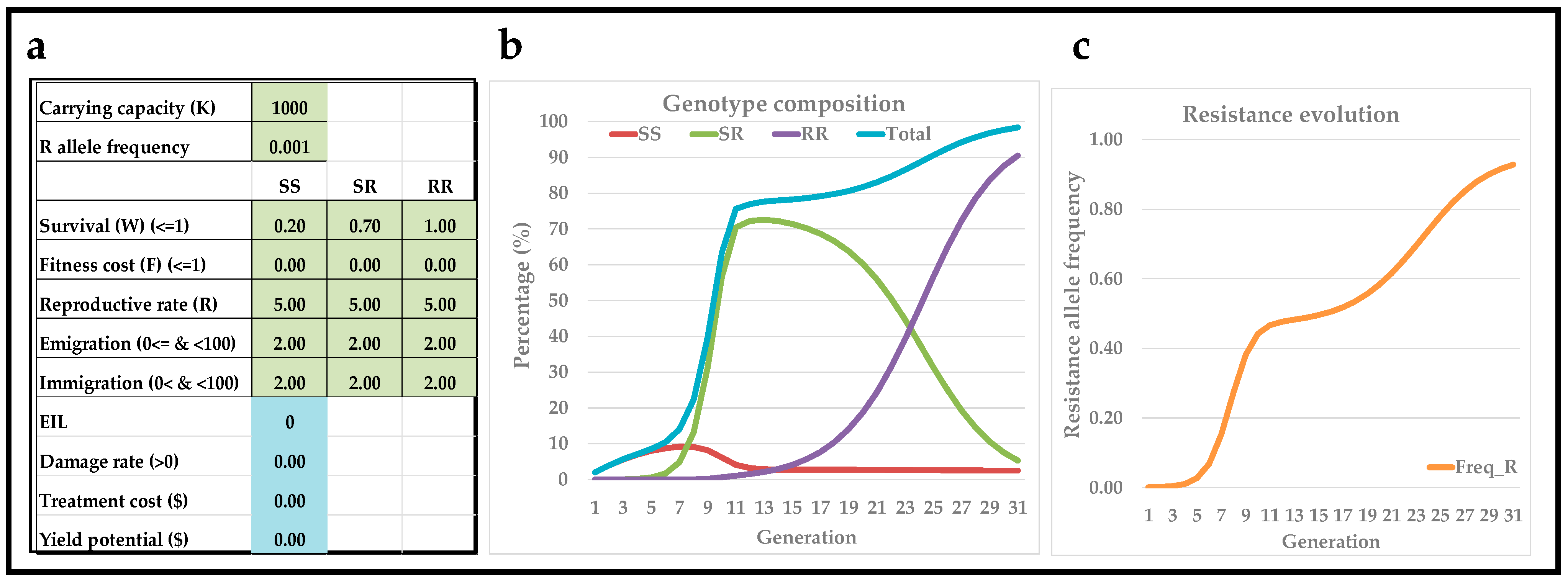
3.2. Interactive Modeling of Resistance Evolution
3.3. Interactive Modeling of Basic Pest Management Economics
4. Discussion
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Essig, E.O. Needed changes in teaching entomology. J. Econ. Entomol. 1944, 37, 563–565. [Google Scholar] [CrossRef]
- Manter, J.A. Charts and forms as aids in teaching economic entomology. J. Econ. Entomol. 1927, 20, 76–79. [Google Scholar] [CrossRef]
- Demmick, R.E. Notes on laboratory methods in teaching entomology. J. Econ. Entomol. 1930, 23, 794. [Google Scholar] [CrossRef]
- Stewart, M.A. The teaching of entomology. J. Econ. Entomol. 1929, 22, 777–781. [Google Scholar] [CrossRef]
- Comstock, J.H. The present methods of teaching entomology. J. Econ. Entomol. 1911, 4, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Fernald, H.T. Present methods of teaching entomology. J. Econ. Entomol. 1911, 4, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Georghiou, G.P.; Taylor, C.E. Pesticide resistance as an evolutionary phenomenon. In Proceedings of the 15th International Conference of Entomology, Washington, DC, USA, 19–27 August 1977; pp. 759–785. [Google Scholar]
- Georghiou, G.P.; Taylor, C.E. Genetic and biological influences in the evolution of insecticide resistance. J. Econ. Entomol. 1977, 70, 319–323. [Google Scholar] [CrossRef] [Green Version]
- Georghiou, G.P.; Taylor, C.E. Operational influences in the evolution of insecticide resistance. J. Econ. Entomol. 1977, 70, 653–658. [Google Scholar] [CrossRef]
- Georghiou, O.P. The evolution of resistance to pesticides. Annu. Rev. Ecol. Evol. Syst. 1972, 3, 133–168. [Google Scholar] [CrossRef]
- Ringland, J.; George, P. Analysis of sustainable pest control using a pesticide and a screened refuge. Evol. Appl. 2011, 4, 459–470. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Gould, F.; Carrière, Y. Delaying evolution of insect resistance to transgenic crops by decreasing dominance and heritability. J. Evol. Biol. 2004, 17, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Gassmann, A.J.; Crowder, D.W.; Carriere, Y. Insect resistance to Bt crops: Evidence versus theory. Nat. Biotechnol. 2008, 26, 199–202. Available online: http://www.nature.com/nbt/journal/v26/n2/suppinfo/nbt1382_S1.html (accessed on 1 February 2022). [CrossRef]
- Gould, F.; Cohen, M.B.; Bentur, J.S.; Kennedy, G.G.; VanDuyn, J. Impact of small fitness costs on pest adaptation to crop varieties with multiple toxins: A heuristic model. J. Econ. Entomol. 2006, 99, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Coulter, J.A.; Nafziger, E.D.; Janssen, M.R.; Pedersen, P. Response of Bt and near-isoline corn hybrids to plant density. Agron. J. 2010, 102, 103–111. [Google Scholar] [CrossRef]
- Singh, A.; Weisser, W.W.; Hanna, R.; Houmgny, R.; Zytynska, S.E. Reduce pests, enhance production: Benefits of intercropping at high densities for okra farmers in Cameroon. Pest Manag. Sci. 2017, 73, 2017–2027. [Google Scholar] [CrossRef]
- Anonymous. IRAC—Insecticide Resistance Action Committee—Resistance Mechanisms. Available online: https://irac-online.org/ (accessed on 23 September 2021).
- Hackett, S.C.; Bonsall, M.B. Type of fitness cost influences the rate of evolution of resistance to transgenic Bt crops. J. Appl. Ecol. 2016, 53, 1391–1401. [Google Scholar] [CrossRef] [Green Version]
- Carriere, Y.; Deland, J.P.; Roff, D.A.; Vincent, C. Life-history costs associated with the evolution of insecticide resistance. Proc. R. Soc. London. Ser. B Biol. Sci. 1994, 258, 35–40. [Google Scholar] [CrossRef]
- Carriére, Y.; Ellers-Kirk, C.; Liu, Y.-B.; Sims, M.A.; Patin, A.L.; Dennehy, T.J.; Tabashnik, B.E. Fitness costs and maternal effects associated with resistance to transgenic cotton in the pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2001, 94, 1571–1576. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Carrière, Y.; Tabashnik, B.E. Fitness costs of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2009, 54, 147–163. [Google Scholar] [CrossRef]
- Demkovich, M.; Siegel, J.P.; Higbee, B.S.; Berenbaum, M.R. Mechanism of resistance acquisition and potential associated fitness costs in Amyelois transitella (Lepidoptera: Pyralidae) exposed to pyrethroid insecticides. Environ. Entomol. 2015, 44, 855–863. [Google Scholar] [CrossRef]
- Zhou, L.; Alphey, N.; Walker, A.S.; Travers, L.M.; Hasan, F.; Morrison, N.I.; Bonsall, M.B.; Raymond, B. Combining the high-dose/refuge strategy and self-limiting transgenic insects in resistance management—A test in experimental mesocosms. Evol. Appl. 2018, 11, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Tchouakui, M.; Miranda, J.R.; Mugenzi, L.M.J.; Djonabaye, D.; Wondji, M.J.; Tchoupo, M.; Tchapga, W.; Njiokou, F.; Wondji, C.S. Cytochrome P450 metabolic resistance (CYP6P9a) to pyrethroids imposes a fitness cost in the major African malaria vector Anopheles funestus. Heredity 2020, 124, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Fabrick, J.A.; Tabashnik, B.E. Can pyramids and seed mixtures delay resistance to Bt crops? Trends Biotechnol. 2016, 34, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Pedigo, L.P.; Hutchins, S.H.; Higley, L.G. Economic injury levels in theory and practice. Annu. Rev. Entomol. 1986, 31, 341–368. [Google Scholar] [CrossRef]
- Higley, L.G.; Pedigo, L.P. The EIL concept. In Economic Thresholds for Integrated Pest Management; University of Nebraska Press: Lincoln, NE, USA, 1996; pp. 9–21. [Google Scholar]
- Pedigo, L.; Rice, M. Entomology and Pest Management, 5th ed.; Waveland Press: Long Grove, IL, USA, 2006. [Google Scholar]
- Stern, V.M.; Smith, R.F.; Bosch, R.v.d.; Hagen, K.S. Integrated control concept. Hilgardia 1959, 29, 81–101. [Google Scholar] [CrossRef] [Green Version]
- Niermans, K.; Meyer, A.M.; Hil, E.F.H.v.d.; Loon, J.J.A.v.; Fels-Klerx, H.J.v.d. A systematic literature review on the effects of mycotoxin exposure on insects and on mycotoxin accumulation and biotransformation. Mycotoxin Res. 2021, 37, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Nansen, C.; Villar, G.d.; Recalde, A.; Alvarado, E.; Chennapragada, K. Phone app to perform quality control of pesticide spray applications in field crops. Agriculture 2021, 11, 916. [Google Scholar] [CrossRef]
- Nansen, C.; Ferguson, J.C.; Moore, J.; Groves, L.; Emery, R.; Garel, N.; Hewitt, A. Optimizing pesticide spray coverage using a novel web and smartphone tool, SnapCard. Agron. Sustain. Dev. 2015, 35, 1075–1085. [Google Scholar] [CrossRef]
- Khodaverdi, H.; Fowles, T.; Bick, E.; Nansen, C. Does drought increase the risk of insects developing behavioral resistance to systemic insecticides? J. Econ. Entomol. 2016, 109, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Nansen, C.; Ridsdill-Smith, T.J. The performance of insecticides—A critical review. In Insecticides; Trdan, S., Ed.; IntechOpen Limited: London, UK, 2013; pp. 195–232. [Google Scholar]
- Gryspeirt, A.; Grégoire, J.-C. Effectiveness of the high dose/refuge strategy for managing pest resistance to Bacillus thuringiensis (Bt) plants expressing one or two toxins. Toxins 2012, 4, 810–835. [Google Scholar] [CrossRef] [Green Version]
- Tabashnik, B.E.; Van Rensburg, J.B.J.; Carrière, Y. Field-evolved insect resistance to bt crops: Definition, theory, and data. J. Econ. Entomol. 2009, 102, 2011–2025. [Google Scholar] [CrossRef]
- Wu, K. Monitoring and management strategy for Helicoverpa armigera resistance to Bt cotton in China. J. Invertebr. Pathol. 2007, 95, 220–223. [Google Scholar] [CrossRef]
- Edwards, K.T.; Caprio, M.A.; Allen, K.C.; Musser, F.R. Risk assessment for Helicoverpa zea (Lepidoptera: Noctuidae) resistance on dual-gene versus single-gene corn. J. Econ. Entomol. 2013, 106, 382–392. [Google Scholar] [CrossRef]
- Kukanur, V.S.; Singh, T.V.K.; Kranthi, K.R.; Andow, D.A. Cry1Ac resistance allele frequency in field populations of Helicoverpa armigera (Hübner) collected in Telangana and Andhra Pradesh, India. Crop Prot. 2018, 107, 34–40. [Google Scholar] [CrossRef]
- Kranthi, K.R.; Kranthi, N.R. Modelling adaptability of cotton bollworm, Helicoverpa armigera (Hübner) to Bt-cotton in India. Curr. Sci. 2004, 87, 1096–1107. [Google Scholar]
- Miyao, G.; Brenna, B.A.; Sumner, D.; Stewart, D. Sample Costs to Produce Processing Tomatoes in the Sacramento Valley and Northern Delta 2017; University of California Cooperative Extension: Napa, CA, USA, 2017. [Google Scholar]
- Torres-Vila, L.M.; Rodríguez-Molina, M.C.; Lacasa-Plasencia, A. Impact of Helicoverpa armigera larval density and crop phenology on yield and quality losses in processing tomato: Developing fruit count-based damage thresholds for IPM decision-making. Crop Prot. 2003, 22, 521–532. [Google Scholar] [CrossRef]
- Bidabadi, N.S.; Isfahani, A.N.; Rouhollahi, A.; Khalili, R. Effective teaching methods in higher education: Requirements and barriers. J. Adv. Med. Educ. Prof. 2016, 4, 170–178. [Google Scholar]
- Caprio, M.A.; Nowatzki, T.; Siegfried, B.; Meinke, L.J.; Wright, R.J.; Chandler, L.D. Assessing risk of resistance to aerial applications of methyl-parathion in western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2006, 99, 483–493. [Google Scholar] [CrossRef]
- Onstad, D.W. Modelling for prediction and management. In Insect Resistance Management; Onstad, D.W., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 453–484. [Google Scholar]
- Caprio, M.A.; Storer, N.P.; Sisterson, M.S.; Peck, S.L.; Maia, A.H.N. Assessing the risk of the evolution of resistance to pesticides using spatially complex simulation models. In Global Pesticide Resistance in Arthropods; Whalon, M.E., Mota-Sanchez, D., Hollingworth, R.M., Eds.; CABI: Wallingford, UK, 2008; pp. 90–117. [Google Scholar]
- Caprio, M.A. Bacillus thuringiensis gene deployment and resistance management in single- and multi-tactic environments. Biocontrol. Sci. Technol. 1994, 4, 487–497. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Mota-Sanchez, D.; Whalon, M.E.; Hollingworth, R.M.; Carrière, Y. Defining terms for proactive management of resistance to Bt crops and pesticides. J. Econ. Entomol. 2014, 107, 496–507. [Google Scholar] [CrossRef]
- Cerda, H.; Wright, D.J. Modeling the spatial and temporal location of refugia to manage resistance in Bt transgenic crops. Agric. Ecosyst. Environ. 2004, 102, 163–174. [Google Scholar] [CrossRef]
- Kang, J.K.; Krupke, C.H.; Murphy, A.F.; Spencer, J.L.; Gray, M.E.; Onstad, D.W. Modeling a western corn rootworm, Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae), maturation delay and resistance evolution in Bt corn. Pest Manag. Sci. 2014, 70, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.C.; Caprio, M.A. IPM use with the deployment of a non-high dose Bt pyramid and mitigation of resistance for Western corn rootworm (Diabrotica virgifera virgifera). Environ. Entomol. 2016, 45, 747–761. [Google Scholar] [CrossRef] [Green Version]
- Lenormand, T.; Raymond, M. Resistance management: The stable zone strategy. Proc. R. Soc. B Biol. Sci. 1998, 265, 1985–1990. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.E.; Georghiou, G.P. Influence of pesticide persistence in evolution of resistance. Environ. Entomol. 1982, 11, 746–750. [Google Scholar] [CrossRef]
- Argentine, J.A.; Clark, J.M.; Ferro, D.N. Computer-simulation of insecticide resistance management strategies for control of colorado potato beetle (Coleoptera, Chrysomelidae). J. Agric. Entomol. 1994, 11, 137–155. [Google Scholar]
- Boivin, T.; Chadœuf, J.; Bouvier, J.-C.; Beslay, D.; Sauphanor, B. Modelling the interactions between phenology and insecticide resistance genes in the codling moth Cydia pomonella. Pest Manag. Sci. 2005, 61, 53–67. [Google Scholar] [CrossRef]
- Crowder, D.W.; Carrière, Y.; Tabashnik, B.E.; Ellsworth, P.C.; Dennehy, T.J. Modeling evolution of resistance to pyriproxyfen by the sweetpotato whitefly (Homoptera: Aleyrodidae). J. Econ. Entomol. 2006, 99, 1396–1406. [Google Scholar] [CrossRef]
- Helps, J.C.; Paveley, N.D.; van den Bosch, F. Identifying circumstances under which high insecticide dose increases or decreases resistance selection. J. Theor. Biol. 2017, 428, 153–167. [Google Scholar] [CrossRef]
- Thrall, P.H.; Oakeshott, J.G.; Fitt, G.; Southerton, S.; Burdon, J.J.; Sheppard, A.; Russell, R.J.; Zalucki, M.; Heino, M.; Ford Denison, R. Evolution in agriculture: The application of evolutionary approaches to the management of biotic interactions in agro-ecosystems. Evol. Appl. 2011, 4, 200–215. [Google Scholar] [CrossRef]
- Liu, Y.; Pan, X.; Li, J. A 1961–2010 record of fertilizer use, pesticide application and cereal yields: A review. Agron. Sustain. Dev. 2015, 35, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nansen, C. An Interactive Teaching Tool Describing Resistance Evolution and Basic Economics of Insecticide-Based Pest Management. Insects 2022, 13, 169. https://doi.org/10.3390/insects13020169
Nansen C. An Interactive Teaching Tool Describing Resistance Evolution and Basic Economics of Insecticide-Based Pest Management. Insects. 2022; 13(2):169. https://doi.org/10.3390/insects13020169
Chicago/Turabian StyleNansen, Christian. 2022. "An Interactive Teaching Tool Describing Resistance Evolution and Basic Economics of Insecticide-Based Pest Management" Insects 13, no. 2: 169. https://doi.org/10.3390/insects13020169